
Abstract
Abstract
Mechanisms by which soluble (i.e., dissolved and colloidal) humic acids (HA) and fulvic acids (FA) alter the transformation kinetics of aromatic substrates by oxidoreductive enzymes such as lignin peroxidase and laccase are not well characterized nor have been quantitatively compared. Hypothesized mechanisms are that soluble HA and FA (1) deactivate the enzyme; (2) act as a competitive substrate; or (3) sequester the aromatic substrate. In this study the transformation of pentachlorophenol (PCP) by purified laccase isolated from Trametes versicolor at 28°C and pH 5.0, in combination with Aldrich HA, Leonardite HA, Suwannee River FA, or Waskish peat FA, were used as a model system to quantitatively test and compare these hypotheses. The four HA and FA did not deactivate laccase after adding catechol as the enzyme's substrate. Slower PCP transformation rates by laccase in the presence of HA and FA were found with higher inhibitor binding constants of HA and FA (Ki, ranging from 3.63±0.78 to 6.47±1.07 μg mL−1) and sorption coefficients of HA and FA to PCP (Kdom, log Kdom ranging from 4.09 to 4.81). Comparison of experimental PCP transformation rates in the presence of soluble HA and FA with predicted transformation rates of dissolved PCP (i.e., PCP not associated with soluble HA and FA) indicated that soluble HA and FA modified PCP transformation by lacasse through sequestration and by acting as a competitive substrate.
Introduction
Although the enzyme-catalyzed coupling products and the corresponding reaction mechanisms have been extensively characterized in studies using model HA monomers (Filazzola et al., 1999; Wang et al., 2002; Nyanhongo et al., 2006; Bialk et al., 2007; Canas et al., 2007; Bialk and Pedersen, 2008) and extracted humic substances (Pflug, 1980; Park et al., 1999, 2000; Kang et al., 2002; Thiele et al., 2002; Witte et al., 2002), very little has been quantitatively characterized regarding the mechanistic influence of soluble (i.e., dissolved and colloidal) HA and FA upon the overall oxidoreductive enzyme kinetics of aromatic compounds. The presence of HA and FA often decreases the transformation rate of the organic compound by the oxidoreductive enzyme, although in a few cases it resulted in an increased transformation rate depending upon the HA and FA source (Claus and Filip, 1998). Zavarzina et al. (2004) demonstrated a difference in the inhibition of HA towards Panus tigrinus laccase for the substrate 2,2′-azino-bis-(3-ethylthiazoline-6-sulfonate): the Ki ranged from 0.003 to 0.025 μg mL−1 for HA isolated from peat soils and chernozem soils respectively. Hypotheses explaining this inhibition are as follows: (1) HA and FA deactivate the oxidoreductive enzyme either by irreversibly changing the quaternary or tertiary structure of the enzyme protein (Műller-Wegerner, 1998) or by HA complexation of copper ions resulting in their dissociation from the enzymatic active site (Keum and Li, 2004); (2) HA and FA act as competitive substrates; and (3) HA and FA sequester the organic compound from the enzyme. Regarding the second hypothesis, numerous laccase-producing fungi have demonstrated an ability to depolymerize, decolor, or degrade HA isolated from coal and aquatic systems (Claus and Fillip, 1998; Fakoussa and Frost, 1999; Scheel et al., 1999). The third hypothesis was posited by many studies, some even specifically proposing that pentachlorophenol (PCP) is sequestered by soluble HA and FA and protected from laccase transformation (Schellenberg et al., 1984; Paolis and Kukkenen, 1997; Peuravuori et al., 2001). The third hypothesis is experimentally supported by fluorescence experiments demonstrating that soluble HA and FA sorb hydrophobic organic compounds (Gadad et al., 2007).
In this study we quantitatively tested these three hypotheses using PCP as a model substrate, laccase as the model enzyme, and four HA and FA ranging in aromaticity and molecular weight. The study compared the three hypothetical mechanisms by which soluble HA and FA are proposed to alter the transformation kinetics of PCP by laccase. By utilizing four HA and FA, a correlation was developed between their chemical structure and the PCP transformation kinetics, along with a proposed mechanistic explanation for the kinetic inhibition by soluble HA and FA. A ratio was developed comparing the experimental PCP transformation rate in the presence of soluble HA and FA against the predicted transformation rate of the dissolved PCP (i.e., total PCP minus PCP sorbed to soluble HA and FA) to provide a quantitative assessment of the relative roles of the competitive substrate mechanism (Hypothesis 2) and the sequestration mechanism (Hypothesis 3) as a function of HA and FA type and concentration.
Experimental Protocols
Chemicals
PCP (98% purity; Sigma-Aldrich) was dissolved in hexane to produce a 1.0 g PCP L−1 stock solution. Purified laccase from Trametes versicolor (>20 units mg−1; Fluka) was prepared fresh for each analysis in a 200 m
Ashley (1996).
Malcolm and MacCarthy (1986).
O'Loughlin et al. (2000).
IHSS (2009).
Beckett et al. (1987).
Thorn et al. (1989).
AHA, Aldrich humic acid; LHA, Leonardite humic acid; SRFA, Suwannee River fulvic acid; WFA, Waskish peat fulvic acid; IHSS, International Humic Substance Society; nd, not determined.
Reaction of laccase with PCP
Km, the Michaelis-Menten constant, and Vmax, the maximum velocity, for the reaction of laccase with PCP were calculated from Lineweaver-Burk plots. Laccase at 20 units mL−1 was incubated with 10, 20, 25, 30, or 40 mg PCP L−1 in 8 mL glass vials containing 0.4 mL of 200 mM sodium acetate buffer at pH 5.0. This pH provided the highest laccase enzyme activity (Ullah et al., 2000). The reaction vials were sealed with Teflon-lined caps and covered with aluminum foil to avoid photobleaching and incubated at 28°C for 4 h while shaken at 200 rpm. Sodium acetate buffer (200 mM) at pH 5.0 was used to adjust the volume. PCP concentrations were measured every hour over 4 h, a period during which the velocity was demonstrated to be constant. Total PCP concentrations were measured by determined by extracting with hexane and analyzing with gas chromatography (6890N; Agilent Technologies) (Supplementary Fig. S1). The effect of HA and FA on laccase activity with PCP was investigated by adding individual HA or FA to a reaction mixture containing the same PCP concentrations listed above and 20 units laccase mL−1. The HA or FA concentrations were 10, 15, and 20 mg HA or FA L−1. PCP concentrations were analyzed every hour over 4 h during which a linear decrease of PCP was observed. Triplicate samples were used for all analyses.
Reaction of laccase with soluble HA and FA
Km and Vmax of laccase reacting with HA and FA were calculated from Lineweaver-Burk plots. Triplicate 4 mL reaction mixtures consisting of 10, 15, 20, 25, 30, or 40 mg HA or FA L−1 and 15 units laccase mL−1 were incubated at 28°C while shaken continuously at 200 rpm. Reaction vials were covered with aluminum foil to protect from photodegradation. Controls were soluble HA and FA without laccase. HA and FA concentrations and laccase activity were analyzed every 24 h over a 96 h period during which there was a linear decrease in the HA and FA concentration. The sample was sacrificed at each analysis. Changes in the HA and FA concentration were monitored by changes in the absorbance at 465 nm using a UV-vis spectrophotometer (Specord 40; Analytik Jena AG) (Zavarzina et al., 2002, 2004) (Supplementary Fig. S2). Laccase did not absorb at this wavelength. Laccase activity was measured by adding 100 m
Sorption of PCP to soluble HA and FA
Dissolved PCP-HA and -FA sorption experiments were modified from O'Loughlin et al. (2000). Dialysis tubing (Spectra/Por® Biotech Cellulose Ester dialysis membranes, MWCO 500 [Spectrum Laboratories Inc.]) was filled with 1.0 mL of 20 mg HA or FA L−1 solution, which was the highest HA and FA concentration used for the reaction of laccase with PCP experiments. The control treatments did not contain soluble HA or FA. The tubing was placed in a 500 mL beaker containing 200 mL of 200 m
Results
Reaction of laccase with either PCP or HA and FA
Laccase isolated from T. versicolor was found to transform PCP with Km and Vmax values of 56 mg L−1 and 18.54 mg L−1 h−1, respectively (Table 2; Lineweaver-Burke plot for PCP and PCP with soluble HA and FA are shown in Supplementary Fig. S3). The Vmax values of laccase for HA and FA transformation ranged from 0.82 to 2.89 mg L−1 h−1 (Table 2); thus, all HA and FA employed in this study were slowly transformed, with the smaller molecular sized SRFA transforming the most rapidly. The Km values for HA and FA transformation by laccase were about 180–1,600 times higher than for the transformation of PCP; thus, PCP is a more favorable substrate. To determine the effect of HA and FA on PCP transformation by laccase, the activity of laccase in the HA and FA reaction mixtures was periodically analyzed using a catechol assay. Laccase activity was constant at about 80% of its full activity for over 96 h for all HA and FA concentrations ranging from 0 to 40 mg L−1 (Supplementary Fig. S4).
Km and Vmax were calculated using Lineweaver-Burke plots.
PCP, pentachlorophenol.
Reaction of laccase with PCP and HA or FA mixtures
A positive relationship between Km for the transformation of PCP and the HA or FA concentration was observed as shown in Fig. 1a, illustrating that PCP transformation by laccase decreased in the presence of soluble HA or FA. The independence of Vmax from the HA and FA concentration (Fig. 1b) indicates that the inhibitory effect of HA and FA was competitive; that is, HA and FA acted as competitive substrates. Enzymatic inhibition by HA and FA was quantified by the inhibitor binding constant, Ki. The values for Ki resulting from the presence of LHA, AHA, SRFA, and WFA were 3.63±0.78 μg mL−1, 5.59±0.96 μg mL−1, 5.99±0.33 μg mL−1, and 6.47±1.07 μg mL−1, respectively; thus, LHA was the strongest inhibitor.
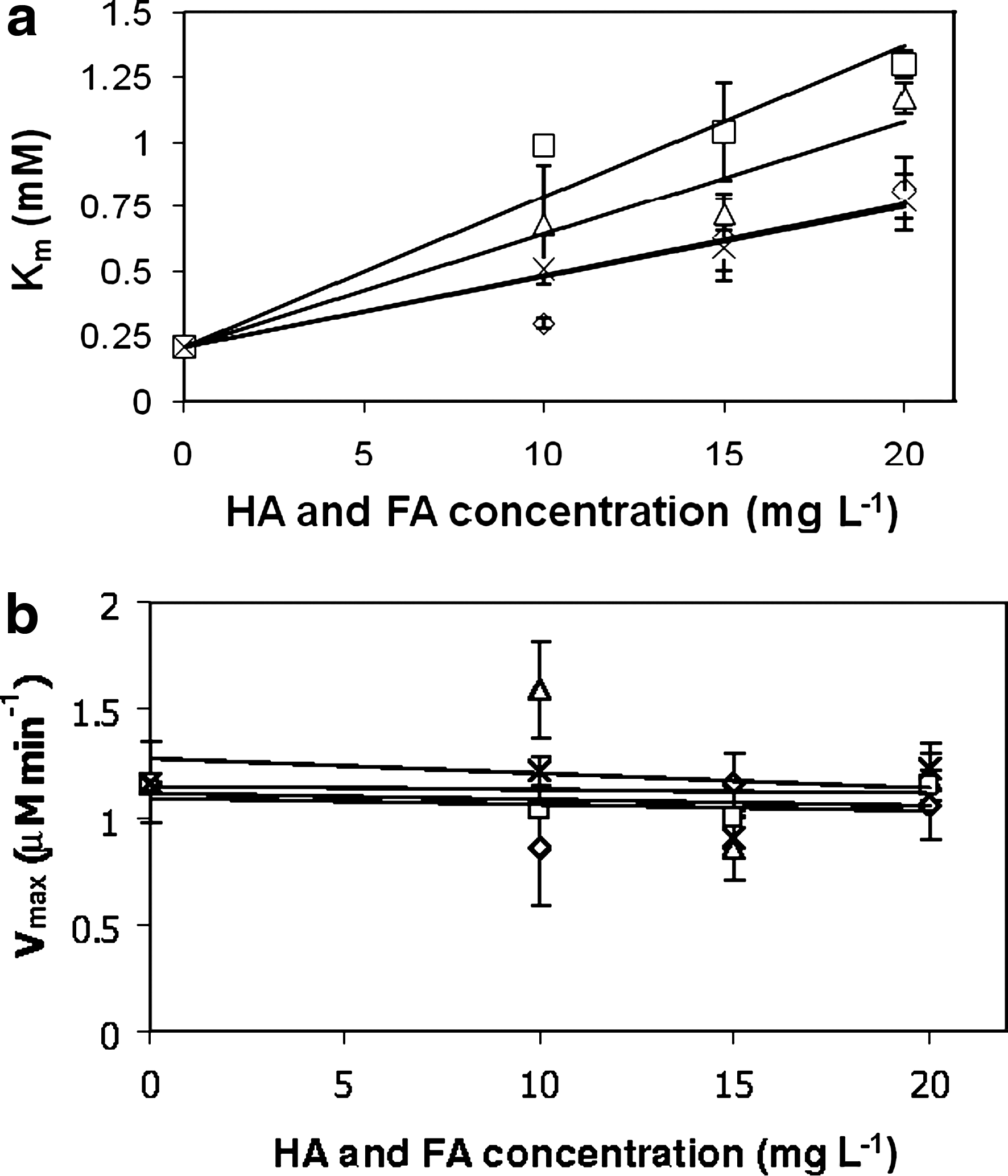
Effect of HA and FA concentrations: ◊, AHA; □, LHA; Δ, SRFA; ×, WFA on
Sorption of PCP to soluble HA and FA
Figure 2 indicates linear sorption isotherms for PCP interacting with all of the HA and FA, each at 20 mg HA or FA L−1. The amount of PCP bound to soluble HA or FA was determined as described by O'Loughlin et al. (2000). All isotherms appeared linear, with average log Kdom (dissolved organic matter normalized sorption coefficient, Kdom=Cdom/Cw, where Cdom is the moles of PCP bound per unit of soluble HA or FA, and Ct is the total concentration of PCP) values for LHA, AHA, WFA, and SRFA equal to 4.81, 4.65, 4.56, and 4.09 L kg−1HS, respectively.
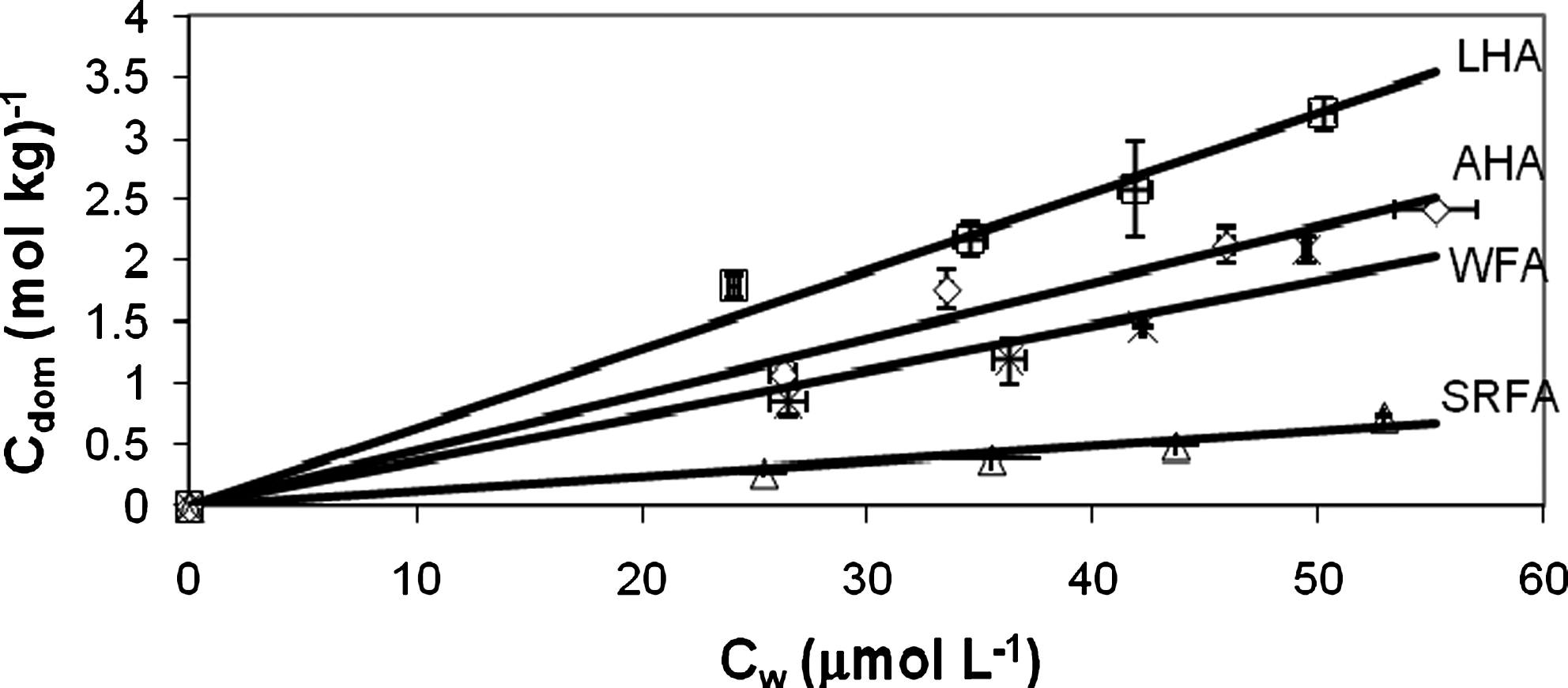
Sorption isotherms of PCP on HA and FA: ◊, AHA; □, LHA; Δ, SRFA; ×, WFA. Error bars represent one standard deviation. R2 for AHA, LHA, SRFA, and WFA were 0.98, 0.99, 0.96, and 0.87, respectively. Cdom, amount of PCP per unit mass of each HA or FA, determined by Cdom=(Ct−Cw)/CHS; Cw, concentration of free PCP.
Discussion
Mechanisms by which HA and FA alter the PCP transformation rate
To elucidate the mechanisms by which soluble HA and FA alter PCP transformation by laccase, three hypotheses were tested: (1) soluble HA and FA deactivate laccase; (2) soluble HA and FA act as competitive substrates; and (3) soluble HA and FA sequester PCP from enzymatic attack. The first hypothesis was considered invalidated since laccase consistently maintained approximately 80% of its full activity in the presence of all the tested humic substances (Supplementary Fig. S4) and over a wide range of humic substance concentrations for up to 96 h at room temperature. The slightly lower laccase activity towards catechol in the presence of soluble HA and FA likely resulted from catechol being sequestered.
The second hypothesis was validated by the reaction of laccase with either only PCP or only a humic substance, as well as the reaction of laccase with mixtures of PCP and humic substances. PCP, HA, and FA were individually transformed by laccase, with PCP being a more favorable substrate as indicated by the lower Km value. When soluble HA or FA was present, the kinetics of PCP transformation by laccase was altered. A positive correlation was observed between Km for PCP transformation and soluble HA or FA concentration (Fig. 1a). However, the inhibitory effect of HA or FA was considered competitive since the Vmax values of PCP transformation were independent from the humic substance concentration (Fig. 1b). Gianfreda and Bollag (2002), as well as Zavarzina et al. (2002), reported similar positive linear relationships between dissolved organic matter concentrations and the inhibitory effect on laccase activity. Furthermore, competitive inhibition has been shown demonstrated for other enzymes incubated with soluble HA and FA such as lignin peroxidase (Wondrak et al., 1989), and pronase, trypsin, and carboxypeptidase (Ladd and Butler, 1971).
The sequestration mechanism hypothesis was investigated through the dissolved PCP-HA and -FA sorption experiments. Since the competitive substrate mechanism (Hypothesis 2) and the sequestration mechanism (Hypothesis 3) may occur simultaneously, we assessed the role of each mechanism by using a ratio of the experimental PCP transformation rate in the presence of soluble HA and FA to the predicted transformation rate of only the dissolved PCP. If the ratio of the experimental transformation rate to the predicted transformation rate equals 1, this suggests that only dissolved PCP is transformed by laccase. A ratio <1 implies HA and FA influences the overall PCP transformation rate by acting as a competitive substrate. For a ratio >1, it suggests that PCP sorbed to HA and FA is available for enzymatic transformation: either sorbed PCP is directly accessible to laccase, or desorption kinetics maintain thermodynamic equilibrium by the rapid release of HA- and FA-sorbed PCP upon enzymatic transformation of the freely dissolved PCP. The results showed that the ratios of HA and FA, at all concentrations were nearly equal to or less than the value of 1 (Table 3; ratio calculations are discussed in Supplementary Data). This indicates that PCP sorbed to soluble HA and FA is not readily available for transformation by laccase within the experimental time frame; thus, sequestration is one mechanism by which soluble HA and FA facilitate a decreased PCP transformation rate. Situations where the ratio values are less than 1 suggest that the decrease in PCP transformation results from HA and FA acting as a competitive substrate. The low and unchanging ratio value for all concentrations of LHA implies that it is a competitive substrate for laccase, which is not surprising considering its high 58% aromaticity. The likelihood that the decreasing ratio values for AHA and SRFA, observed as a function of increasing HA and FA concentrations, result from dissolved PCP sorbing to high molecular weight, laccase-induced polymers of PCP with HA and FA (Roy-Arcand and Archibald, 1991) seems unlikely given the linear response of PCP transformation rates as a function of PCP concentration. This unlikelihood is further supported by Supplementary Fig. S5, illustrating that the linear response is independent of HA or FA concentration. If sorption of dissolved PCP to polymerized PCP-HA or FA was significant for the utilized experimental conditions and reaction time periods, non-linear behavior would be expected at higher PCP concentrations where more polymerized material would be present. As such, this study does not address the influence of PCP-HA or -FA polymers, which may form at higher PCP, HA, and FA concentrations or over longer time periods of enzymatic transformation, upon the enzymatic transformation and concurrent fate of aromatic substrates.
Total PCP concentration=5.63×10−5 M for all experiments. Calculations are presented in Supplementary Data.
A=k′(5.63×10−5); k′ is a slope of the plotting of transformation rate and PCP concentration.
B=Cw=Ct/[(Kdom×CHS)+1]
C=B/4 h; 4 h was the reaction time carried out in the experiment.
Correlation between HA and FA chemical properties and PCP transformation
The inhibitory effect (Ki) of HA and FA on PCP transformation displayed a negative correlation with both molecular weight (R2=0.99) and % aromaticity (R2=0.74) (Fig. 3a, c), showing that HA and FA became a more competitive substrate with increasing % aromaticity and molecular weight. However, Km for HA and FA transformation by laccase was positively correlated with molecular weight and % aromaticity, with R2 equal to 0.99 and 0.79, respectively (Fig. 3b, d). There was a strong relationship between the PCP sorption coefficient, Kdom, and % aromaticity (R2=0.97) (Fig. 3d). Additionally, a weaker relationship between Kdom and molecular weight (R2=0.73) was found (Fig. 3b). Thus, this study indicated that % aromaticity and molecular weight of HA and FA were positively correlated with the inhibition of laccase activity for PCP transformation. Similarly, Morimoto et al. (2003) reported a positive correlation between the inhibition of enzymatic transformation of PCP and the degree of unsaturation of HA and FA. Since aromatic compounds are known to be laccase substrates, HA and FA should become more competitive substrates with increasing % aromaticity. Likewise, sequestration of PCP should be favored by increasing % aromaticity and molecular weight of HA and FA as these parameters favor noncovalent interactions between aromatic compounds and soluble HA and FA (Gadad et al., 2007). Similarly, Zavarzina et al. (2004) reported a positive correlation between HA hydrophobicity, determined chromatographically, and HA inhibitor capability towards laccase isolated from Panus tigrinus.
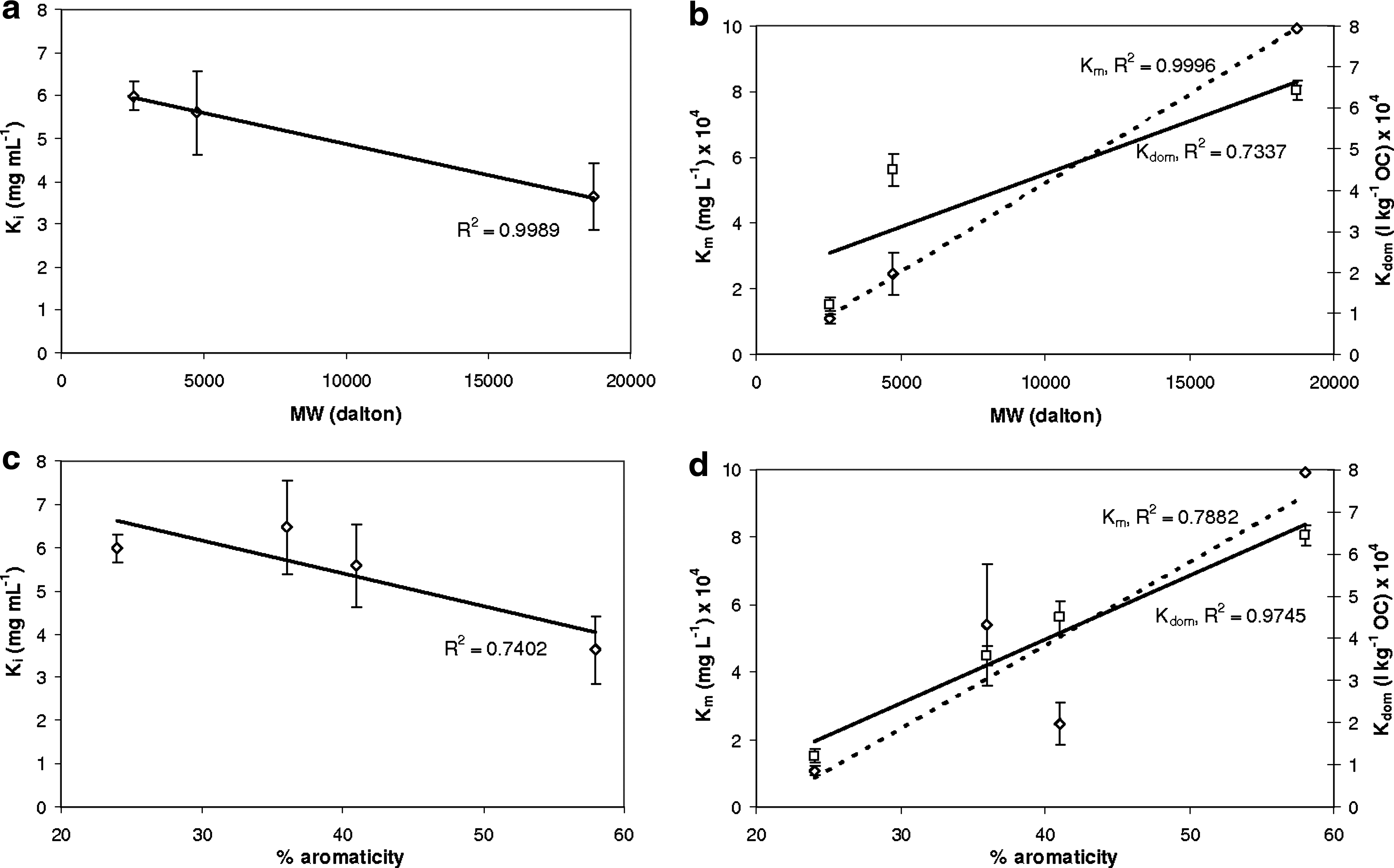
Relationship between kinetic and thermodynamic parameters and HA and FA properties:
Meanwhile, the positive correlation between Km and the molecular weight and the % aromaticity of the HA and FA is unexpected; a greater concentration of aromatic functional groups as available substrates would generally be expected to lead to lower values of Km. However, aromatic functional groups may be sequestered within the HA or FA macromolecule and therefore not readily available for enzymatic attack. Additionally, UV-Vis spectroscopy may not be the optimal analytical method for monitoring the enzymatic transformation of soluble HA and FA since increases in UV-Vis absorbance have been reported as a result of polymerization of HA and FA by oxidative enzymes (Zavarzina et al., 2004; AbuBaker et al., 2008).
The strong relationship between the PCP sorption coefficient, Kdom, and % aromaticity supports the idea of PCP sequestration by soluble HA and FA as a protective mechanism. Similar relationships have been previously reported (Chin et al., 1997; Perminova et al., 1999; Uhle et al., 1999; Gadad et al., 2007) and are explained by the increased polarizability of the aromatic-rich functional groups (Gauthier et al., 1987; Chin et al., 1997). Increasing the polarizability of HA and FA result in an increase in van der Waals interactions and π−π interactions between the solute and substrate. In an aromatic-rich environment such as LHA, PCP would be susceptible to these interactions. The lack of a strong relationship between Kdom and molecular weight in our study highlights the importance of the soluble HA and FA aromatic functional groups interacting with PCP.
Summary
Based upon these results, it is concluded that soluble (i.e., dissolved and colloidal) HA and FA modify the transformation of aromatic substrates by oxidative enzymes, in particular laccase, through a combination of two mechanisms: HA and FA acting as competitive substrates and, secondly, sequestration of the aromatic substrates by the aromatic functional groups of HA and FA. The % aromaticity and molecular weight of the HA and FA are indicated as important chemical parameters with both being relevant for overall inhibition of enzymatic transformation, whereas the former is most appropriate for assessing the association of PCP with the soluble HA and FA. As such, in modeling the enzymatic transformation of aromatic substrates, it must be considered that the freely dissolved aromatic substrate will be degraded while soluble HA and FA act as competitive substrates. Moreover, increasing the % aromaticity of the soluble HA or FA will lead to a higher propensity for sequestration of the aromatic substrate resulting in decreasing transformation rates and requiring a longer time for substrate oxidation.
Footnotes
Acknowledgments
The authors thank Dr. Pairoh Pinphanichakarn for her invaluable suggestions. We also thank the National Center of Excellence for Environmental and Hazardous Waste Management, Chulalongkorn University, and the Chulalongkorn University Graduate Thesis Grant for financial support.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.