
Abstract
Abstract
This study investigated the evolution of archaeal and bacterial populations of two submerged anaerobic membrane bioreactors (SAMBRs) operating at a mean solids residence time of 30 (SAMBR30) and 300 days (SAMBR300) at mesophilic and psychrophilic temperatures. SAMBRs were fed with leachate produced in a hydrolytic reactor (HR) treating the organic fraction of municipal solid waste. The archaeal fingerprint using denaturing gradient gel electrophoresis showed different populations in the first and second stage of the two-stage anaerobic process. A build up of volatile fatty acids (VFAs) was observed at 20°C in SAMBR30; whereas in SAMBR300, the VFAs only built up at 10°C. The dominant bacterial species in the HR belonged to Prevotella and Thauera, whereas the dominant ones in SAMBR300 belonged to Sphingobacteriales, Anaerovorax, Spirochaetaceae, Hydrogenophaga, Ralstonia, Prevotella, and Smithella. Low bacterial diversity in SAMBR30 compared with SAMBR300 resulted in a persistently high soluble chemical oxygen demand (>2 g/L) in the bulk reactor due to an insufficient residence time for bacteria to carry out the degradation of recalcitrant chemical oxygen demand found in the leachate.
Introduction
There is a lack of information regarding the microbial populations in anaerobic digestion, especially at lower temperatures. Methanosaeta and Methanosarcina were reported to be dominant at low and elevated concentrations of acetate, respectively; whereas Methanobacteriaceae were the most abundant hydrogenotrophic methanogens in mesophilic digesters fed with simulated municipal solid waste (Griffin et al., 1998). Connaughton et al. (2006) noted the emergence of hydrogenotrophic Methanocorpusculum-like and Methanococcales and Methanobacteriales organisms when treating synthetic volatile fatty acid (VFA)-based wastewater at 18°C using a mesophilic inoculum. Moreover, they noticed an apparent reduction in the abundance of acetoclastic methanogens from the Methanosaeta and Methanosarcina groups. In addition to methanogenic Archaea, Crenarchaeota were also detected. Regarding bacterial species, McHugh et al. (2004) have observed variations in population structure, but no clear pattern could be discerned when gradually decreasing the temperature from 37° to 16°C in a reactor treating a synthetic wastewater consisting of acetate, ethanol, butyrate, and propionate. At 16°C, they detected gram-positive bacteria such as members of the groups Bacillus, Lactobacillus, and Streptococcus; whereas the remaining clones showed strong similarities with Bacteroides and Spirochaeta. Their studies focused on synthetic wastewaters; however, very little is known about bacterial communities in anaerobic membrane bioreactors treating complex wastewater such as landfill leachate. Moreover, the identification and selection of such microorganisms capable of treating leachate at low temperatures would be important when monitoring the stability of a psychrophilic digester and optimizing its performance.
Studying the response of anaerobic reactors to low temperatures is vital, especially when the temperature change affects the rate-limiting step by altering the growth rate of individual species, and, hence, changes the predominant group of microorganisms in the long run. In addition, heating of anaerobic reactors requires energy and capital expenditure, and, hence, efficient digestion of wastewater at low temperatures (5°C–10°C) would be highly desirable to lower costs and increase energy output. Hence, the identification and selection of such microorganisms capable of treating leachate at low temperatures would be important when monitoring the health of a psychrophilic digester and optimizing its performance. The development of high-rate anaerobic reactor designs such as the submerged anaerobic membrane bioreactor (SAMBR) containing high biomass concentrations should enable the biomass to operate at lower temperatures where the specific rate per unit of biomass is lower. However, the microbial population in suspension in the SAMBR and membrane biofilm operating at low temperatures is not well documented. Thus, the purpose of this study was to investigate the evolution of archeal and bacterial species in suspension and on the membrane biofilm of the SAMBR treating municipal solid waste leachate at ambient and psychrophilic temperatures.
Materials and Methods
Wet mesophilic two-stage lab-scale anaerobic process
The hydrolytic reactor (HR, 10 L working volume) producing the leachate was an acrylic cylinder with a stainless steel mesh that followed a concentric arrangement inside the cylinder and had a grid of 1 mm holes (Fig. 1). A stirrer moved inside the mesh, thus allowing two pieces of rubber to rub against the perforated mesh: the speed of the stirrer was 40 rpm (Heidolph RZR 2021) and was operated intermittently (15 min on, 15 min off). The HR was inoculated with 10 L of anaerobic sludge, was maintained at 37°C throughout the study, and was fed daily with a simulated feedstock of the organic fraction of municipal solid waste at an organic loading rate of 4–5 g volatile solids/L/day until day 46, and 10 g volatile solids/L/day from day 47 until the end of the experiment. Details of the HR, composition, and properties of the municipal solid waste feedstock can be found elsewhere (Trzcinski and Stuckey, 2009). In addition to screening in the HR the leachate pumped from the HR was also centrifuged (1500 rpm for 20 min; Heraeus Biofuge Stratos) daily to avoid a build-up of recalcitrant suspended solids in the laboratory scale SAMBR, and the supernatant was poured into an intermediate tank from which the SAMBRs were fed. The leachate was treated in two SAMBRs, SAMBR30 and SAMBR300, operating at mean solids residence times (SRTs) of 30 and 300 days, respectively. The SRT was controlled by removing a known volume from the SAMBR daily. The SAMBRs were 3 L reactors fitted with a Kubota polyethylene flat sheet membrane (0.1 m2 surface area—pore size of 0.4 μm) (Trzcinski and Stuckey, 2009). The SAMBR contained a standing baffle designed to direct the fluid to the upcomer and downcomer regimes. The biomass was continuously mixed using headspace biogas that was pumped through a stainless steel tube diffuser to generate coarse bubbles. The bubbles pushed the sludge flow upward between the membrane module and the reactor wall in the upper section. The biogas sparging rate was set at 3 L/min to minimize cake formation on the membrane. SAMBR300 was inoculated with an acclimatized anaerobic sludge treating leachate, and its initial mixed liquor total suspended solids (MLTSS) and mixed liquor volatile suspended solids (MLVSS) were 5.2 and 3.9 g/L, respectively; whereas in SAMBR30, it was 4.4 and 3.2 g/L, respectively. The SAMBRs were operated at 35°C until day 95, at 20°C from day 95 to 100, and at 10°C after day 100 using a chiller (Model RC400, Grant). Microbial communities from the bioreactors were sampled by taking a small volume with a syringe, whereas the sample from the biofilm formed on the membrane was taken at the end of the experiment after opening the bioreactor. The permeates of SAMBR30 and SAMBR300 were recycled to the HR or used to moisten the fresh feedstock to minimise fresh water consumption (Fig. 1).
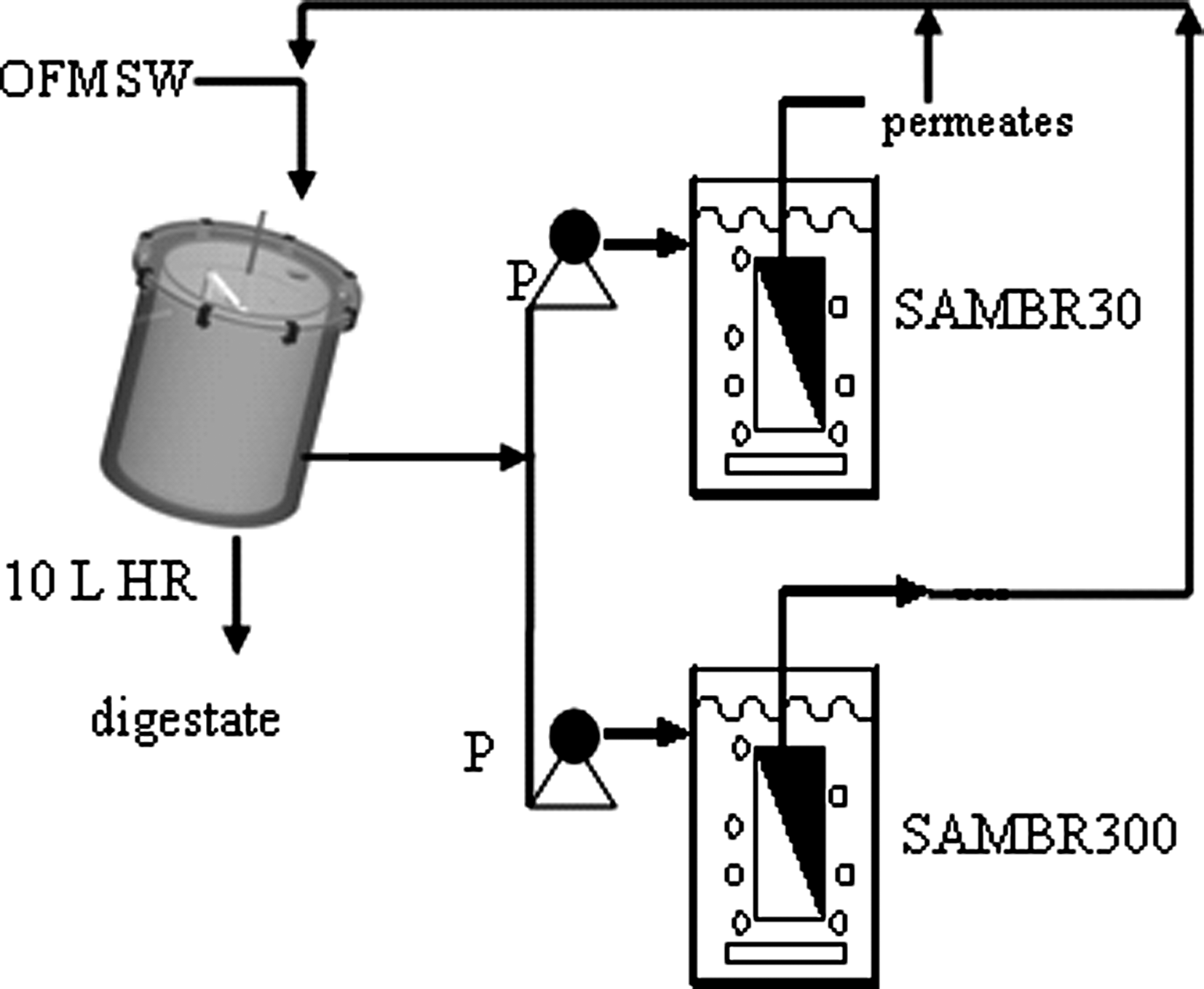
Schematic diagram of the anaerobic process. OFMSW, organic fraction of municipal solid waste; HR, hydrolytic reactor; P, peristaltic pump; SAMBR30 and SAMBR300, submerged anaerobic membrane bioreactor at 30 and 300 days average solid retention times.
Analytical methods
The measurement of pH (Jenway) was accurate to within±0.02 units. MLTSS, MLVSS, sulfate concentration, and soluble chemical oxygen demand (SCOD is COD after filtration through a 0.2 μm filter) were measured as described in Standard Methods (APHA, 1999). Their coefficient of variation (COV) for 10 identical samples was 4%, 3.1%, 5%, and 2.6%, respectively. VFAs (acetic, propionic, iso- and n-butyric, iso- and n-valeric, and n-caproic acids) were measured using a Shimadzu Gas Chromatograph with a flame-ionization detector and a SGE capillary column (12 m×0.53 mm ID-BP21 0.5 μm). The mobile phase was helium; and the injector, column, and detector temperatures were 200°C, 80°C, and 250°C, respectively. The COV was 3% for 10 identical samples. The water (2% H2SO4, 10% NaCl) displacement method was used to measure the gas production rate (Lbiogas/Lreactor/day) in the SAMBR. The composition of biogas was determined using a Shimadzu GC-TCD fitted with a Porapak N column (1500 × 6.35 mm). The COV for 10 identical samples was±2%.
For the denaturing gradient gel electrophoresis (DGGE) analysis, the microbial community structure was evaluated when the reactor reached equilibrium state and after several alterations in the operational conditions. Samples were collected from the SAMBRs sludge, and the DNA from the mixed culture was extracted using the FastDNA SPin for soil kit from MP Biomedicals. The polymerase chain reaction (PCR) mixture (50 μL) contained 2 μL of each primer, 0.2 μL of the Taq Polymerase (Promega), 2 μL of dNTPs stock solution (10 μM), 5 μL of Taq buffer solution containing MgCl2, and between 1 and 20 μL of DNA template. The final volume was adjusted to 50 μL using DNA-free water. Archaeal DNA was amplified using a nested PCR in a G-Storm thermocycler. First, the DNA template was amplified using the primers 46F (5′-YTA AGC CAT GCR AGT-3′) and 1017R (5′-GGC CAT GCA CCW CCT CTC-3′) in 25 μL. The temperature program was as follows: initial denaturation 95°C for 3 min, then 35 cycles of denaturation at 95°C for 1 min, annealing at 40°C for 1 min, elongation at 72°C for 1 min, and the final elongation at 72°C for 7 min (Gray et al., 2002; Akarsubasi et al., 2005). Then, 1 μL of the PCR product was used for the second PCR in 50 μL using the primers 344F-GC (5′-CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG GAC GGG GHG CAG CAG GCG CGA-3′) and Univ522R (5′-GWA TTA CCG CGG CKG CTG-3′). The temperature program was as follows: initial denaturation at 95°C for 3 min, then 35 cycles of 95°C for 1 min, 53°C for 1 min, 72°C for 1 min, and the final elongation at 72°C for 7 min (Gray et al., 2002; Akarsubasi et al., 2005). Bacterial DNA was amplified using the primers (with GC clamp in italics) 341F-GC (5′-CGC CCG CCG CGC GCG GCG GGC GGG GCG GGG GCA CGG GGG G CCT ACG GGA GGC AGC AG-3′) and 907R (5′-CCG TCA ATT CCT TTR AGT TT-3′) (Muyzer et al., 1993) in 50 μL PCR volume. The temperature program was as follows: initial denaturation at 94°C for 5 min, then 30 cycles of 94°C for 1 min, 52°C for 1 min, 72°C for 1 min, and the final elongation at 72°C for 10 min. Presence of PCR products was confirmed by electrophoresis on 1% agarose gels stained with ethidium bromide. DGGE of the 16Sr DNA PCR products was carried out using the D CODE system (Bio-Rad Laboratories Ltd.) according to the manufacturer's instructions and protocols. A 6% polyacrylamide gel was made up if bacterial primers were used, whereas 8% was used for archaeal primers. The PCR products were electrophorised at 60 V for 16 h at 65°C with a denaturing gradient ranging from 40% to 60% for bacterial primers and from 45% to 65% for archaeal primers. A 100% solution contains 7 M urea and 40% formamide. The gel was stained in 1× TAE buffer containing SYBR green before visualizing on a UV transilluminator and photographed. The brightest DGGE bands were cut out to determine which species were dominant in each reactor. The bands were then eluted in DNA-free water overnight at 4°C and re-amplified using the same set of primers as for the first PCR. The purified PCR product was then cloned with the pCR 2.1-TOPO cloning kit (Invitrogen) according to the manual instructions. Briefly, 1 μL of vector was carefully mixed with 1 μL of salt solution (Invitrogen) and 4 μL of purified PCR product. Meanwhile, chemically competent Escherichia coli were placed on ice; and 2 μL of the cloning reaction was mixed with the competent cells. The cells were transformed using a heat-shock protocol (30 s at 42°C), then incubated for 1 h at 37°C in SOC medium (Invitrogen), and eventually spread onto Luria-Bertaniagar (LB) agar plates containing 50 μg/mL of kanamycin to incubate overnight at 37°C. Three well-separated colonies were picked from the plate using a pipette tip and were grown between 12 and 16 h in LB medium (without agar) at 37°C. The plasmid DNA from cloned colonies was then purified using the purification kit from Qiagen (QIAprep Spin Miniprep Kit), and the resulting DNA was sent for sequencing (Cogenics). DNA sequences analyses were performed using the BLAST server of the National Center for Biotechnology Information (www.ncbi.nlm.nih.gov). Sequences from the DGGE analysis were deposited in the GenBank database under the accession numbers HQ113101 for the Archaea and HQ111174 to HQ111193 for the bacteria.
Results and Discussion
SAMBR performance
The SCOD and VFAs concentrations were monitored in bulk and the permeates of SAMBR30 and SAMBR300 to control the stability at various HRTs and operating temperature. Generally, the VFA concentration in the leachate from the HR was in the range 1–3 g/L, but peaked on some occasions up to 7 g/L. Acetic and propionic acids were the main acids produced, so the other VFAs were omitted from Fig. 2. Figure 2 (top) shows that SAMBR30 could start up at 3.7 days HRT and 35°C, and that it could achieve >90% SCOD removal at 1.5 days HRT. When the temperature dropped to 20°C, SAMBR30 did not cope, and the SCOD removal dropped to below 50%, whereas VFAs started to build up. SAMBR300 was fed with centrifuged leachate from the HR from day 34 onward, because results obtained from SAMBR30 showed a high SCOD in the bulk (>2 g/L), presumably due to the low sludge retention times of 30 days. SAMBR300 was, therefore, run at 300 days solid retention time to investigate whether lower-bulk SCODs could be attained. The initial HRT was set at 19 days to check that the inoculum was active, and was then gradually reduced to low HRTs and temperatures. SAMBR300 achieved 95% SCOD removal at 1.1 days HRT and 20°C (Fig. 2 bottom). When the temperature dropped to 10°C, SCOD and VFAs concentrations started to build up in the bulk, presumably due to lower degradation rates at psychrophilic temperatures.
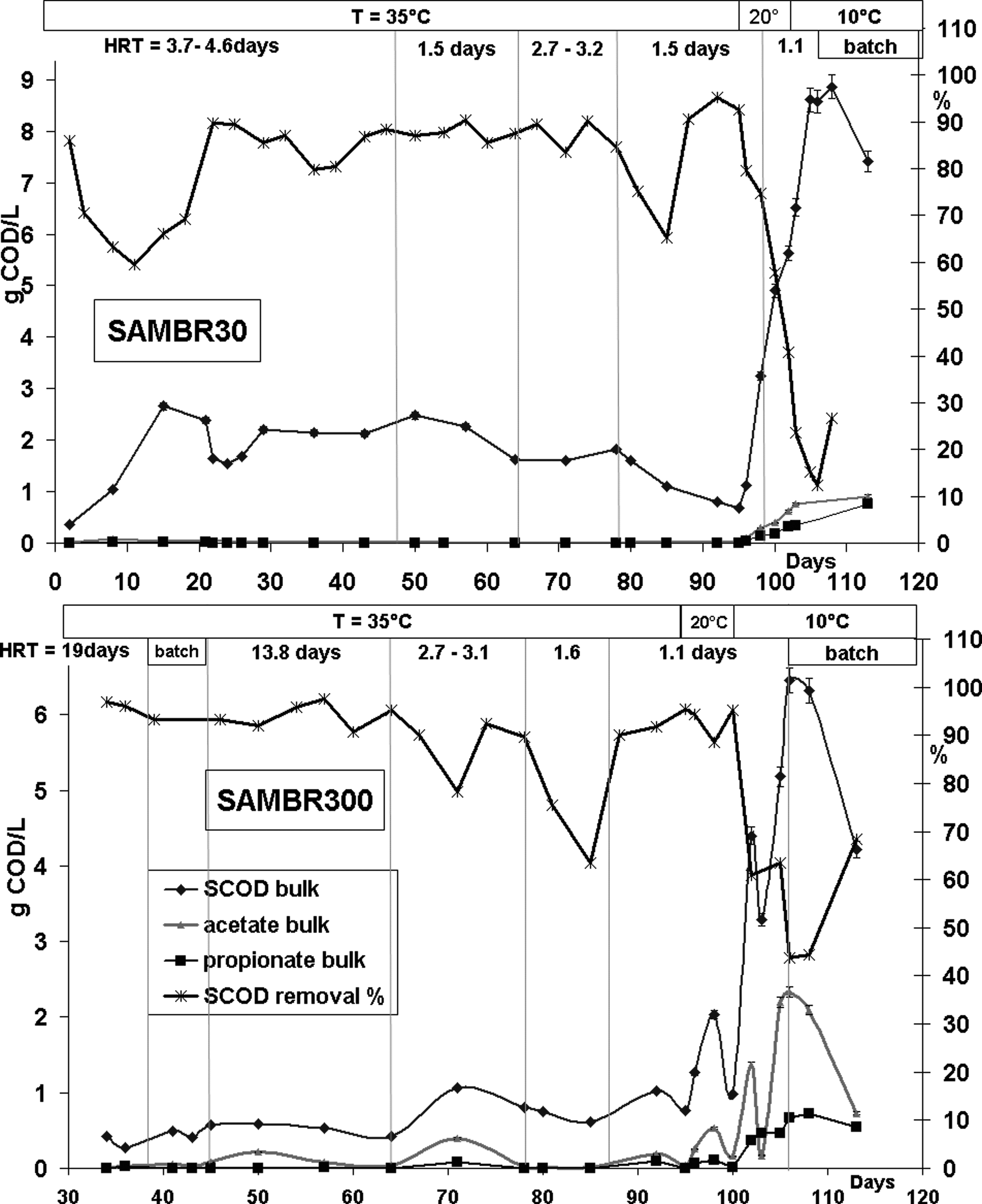
Evolution with time of the soluble chemical oxygen demand (SCOD), acetate, propionate concentrations in SAMBR bulk and the SCOD removal percentage in SAMBR30 (top) and SAMBR300 (bottom).
The methane content and the biogas production rate are shown in Table 1 where it can be seen that a methane percentage of 70% was obtained in SAMBR300, even at 20°C. However, a drop in the biogas production rate was observed in both SAMBRs when the temperature was set at 20°C. At 10°C, the methane percentage dropped to 55% in the SAMBR headspace, whereas the gas production rate was only 1 and 1.7 Lbiogas/Lreactor/day in SAMBR30 and SAMBR300, respectively.
Some variability in the MLTSS measurement can be attributed to some particles present in the leachate fed to the SAMBR, although the leachate was centrifuged beforehand. The measured MLTSS does not include the biomass attached on the reactor walls and the membrane. The attached biomass tended to increase over time.
GPR, gas production rate; MLTSS, mixed liquor total suspended solids; SAMBR, submerged anaerobic membrane bioreactor.
Archaeal population in HR, SAMBR30, and SAMBR300
The archaeal populations from the HR, SAMBR30, and SAMBR300 were analyzed using the DGGE technique as described in the Materials and Methods section, and the gel picture is shown in Fig. 3. The brightest bands were cut and sequenced, but no match with known Archaea was found. Only one sequence from a band in the gel was found to have a high similarity (100%) to the sequence of Methanosarcina mazeii. It was found in SAMBR30, but its presence in the other reactors cannot be ruled out because of bands at similar migration distances in the gel. Further, the DGGE picture shows that the SAMBR can start up within a week using an acclimatized inoculum, as methanogens were detected in SAMBR30 after only 2 days and this was confirmed by the absence of VFAs at a HRT of 3.7 days (Fig. 2, top).
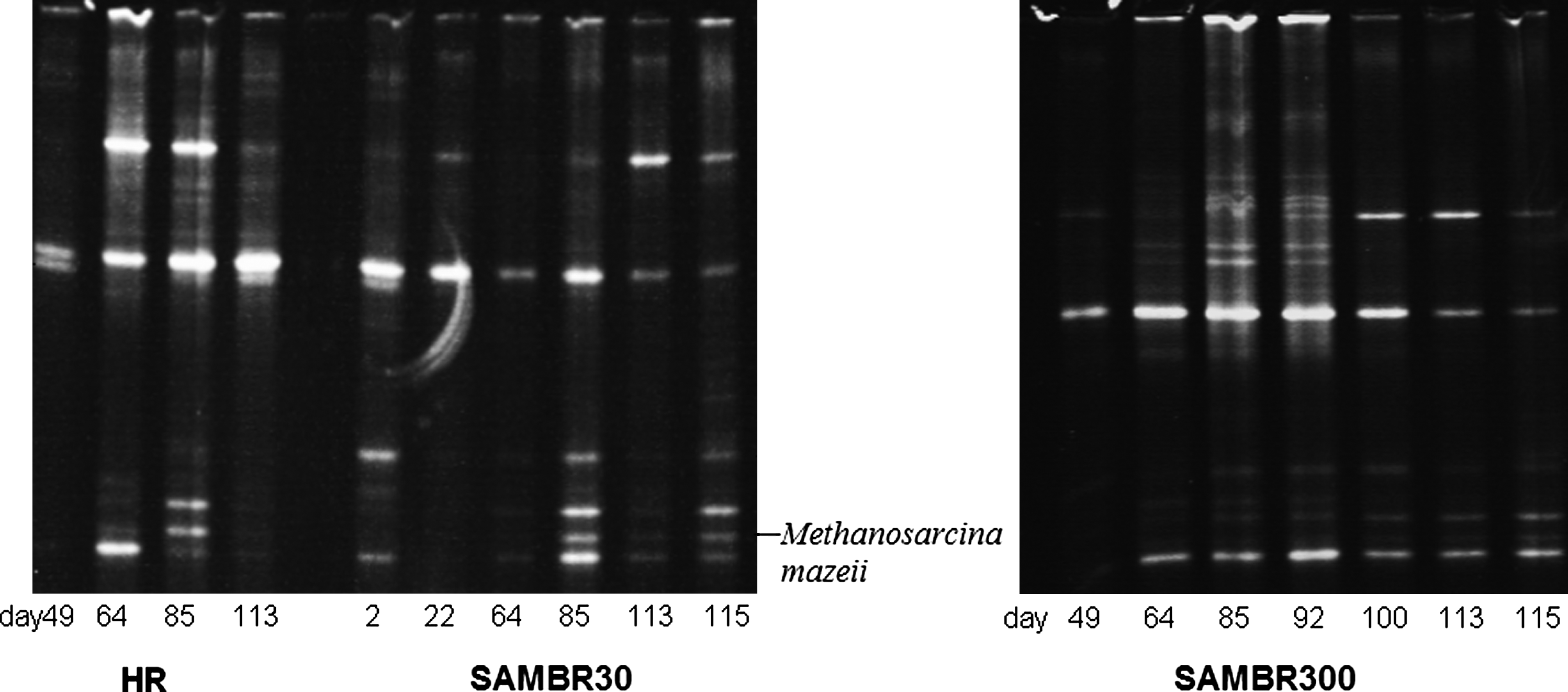
Evolution with time of the archeal population in the HR (left), SAMBR30 (middle), and SAMBR300 (right). Samples on day 115 were taken from the biofilm formed on the submerged membrane.
In SAMBR30, the detected archaeal diversity was lower than in SAMBR300 for the samples taken on days 64 and 85. There were a maximum of five or six bands detected in the former, whereas the latter had up to 12 bands. The lower SRT in SAMBR30 appears to be the main reason; nevertheless, the lower archaeal diversity did not impede VFA degradation, as their concentrations remained low until the shock to low temperatures on day 95. Eventually, on day 115, the SAMBR was opened; and a sample from the biofilm on the membrane was taken. The archaeal population on the biofilm formed on the membrane was very diverse, and this shows evidence that Archaea remain attached to the membrane of a SAMBR even at very low HRTs, which may not necessarily be the case in a more conventional anaerobic reactor in which the hydraulic flowrate is suddenly increased. However, it should be borne in mind that differences between lanes in the gel may be the result of different amounts of DNA loaded into each well. On day 113, a lower archaeal activity was observed at 10°C, as indicated with a build up of acetate and propionate to 760 and 340 mg/L, respectively. This build up started even when the temperature was set at 20°C on day 95, which indicates that the number of methanogens was not sufficient to cope with the load at 20°C, because the specific reaction rate of the methanogens decreased. It can be seen from the bands on lane 115 that the biofilm formed on the membrane in the SAMBR contained methanogenic species even at 10°C.
Two main species were detected on day 49 in SAMBR300 at an HRT of 14 days, and this was sufficient to keep VFA concentrations low; however, with time, the number of bands increased to >12 as the HRT was reduced on day 87 to 1.1 days at 35°C. The sample taken on days 85 and 92 were the samples containing the most bands, but only one dominant species was sufficient to guarantee complete VFA degradation as it was the case on day 49. Interestingly, the population changed markedly as the temperature was set at 20°C on day 95 which resulted in a decrease in the number of bands to five. This showed that different species responded differently to the shock from 35°C to 20°C. This observation was supported by the fact that on day 113, a build up of VFAs was observed at 10°C due to lower growth rate and metabolic activity of Archaea at such low temperatures. In contrast with SAMBR30, SAMBR300 could maintain complete VFA degradation at 20°C due to the high SRT that enabled a greater number of methanogens to be present in the bulk, but not at 10°C, where the acetate and propionate concentrations increased to 2330 and 670 mg/L, respectively. The last lane (day 115) in Fig. 3 shows the archaeal population present on the biofilm formed on the Kubota membrane. Based on our DGGE picture, it was found that in both SAMBRs, the archaeal fingerprint of the biofilm was very similar to the one in the bulk, and this may be because there were psychrotolerant mesophiles present.
Bacterial population in HR, SAMBR30, and SAMBR300
Figure 4 shows the bacterial population in the HR, SAMBR30, and SAMBR300; and the closest match associated with the DNA extracted from bands 1 to 21 can be found in Table 2. The lane noted MS on the right-hand side of Fig. 4 is a sample of the anaerobic sludge from the Mogden wastewater treatment plant (United Kingdom) that was used as inoculum in the experiment. The bacteria identified were an unclassified Bacteroidetes and a Propionibacteriaceae bacterium. By comparing this lane with other lanes, it can be seen that an important change in bacterial population occurred during this study.
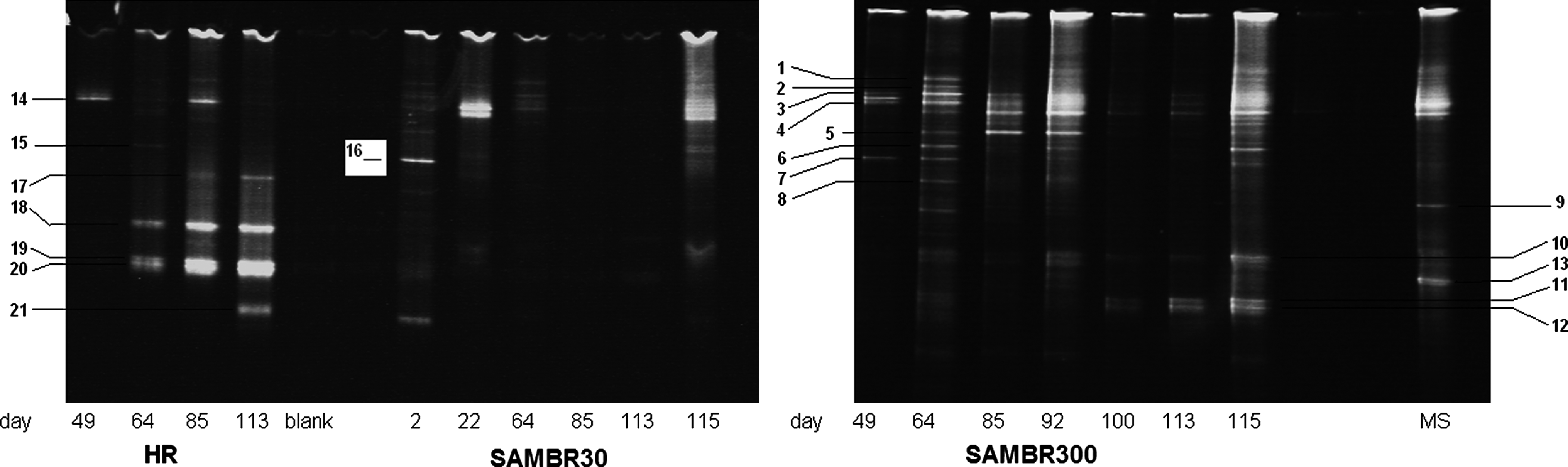
Evolution with time of the bacterial population in the HR (left), SAMBR30 (middle), and SAMBR300 (right). MS, Mogden Sludge, which is an anaerobic sludge taken from Mogden wastewater plant (United Kingdom), was used as inoculum. Samples on day 115 were taken from the biofilm formed on the submerged membrane. Numbers refer to the band identification number described in Table 2.
HR, hydrolytic reactor; SAMBR30, submerged anaerobic membrane bioreactor operating at 30 days average solid retention time; SAMBR300, submerged anaerobic membrane bioreactor operating at 300 days average solid retention time; MS, Mogden Sludge, that is, initial anaerobic inoculum from Mogden wastewater treatment plant (United Kingdom).
From Fig. 4 and Table 2, it can be seen that the dominant bacterial species in the HR belonged to Prevotella and Thauera; whereas the dominant ones in SAMBR300 belonged to Sphingobacteriales, Anaerovorax, Spirochaetaceae, Hydrogenophaga, Ralstonia, Prevotella, and Smithella, although the absence of the species in the HR cannot be totally ruled out. This suggests that the membrane in the SAMBR allows different genus to appear in the second-stage reactor compared with a single-stage process. Madigan and Martinko (1984) reported that members of the Prevotella genus are a starch decomposer found in the rumen, and their fermentation products are formate, acetate, and succinate.
As far as SAMBR30 was concerned, the bacterium Brachymonas denitrificans was found in band 16 on day 2. It is an aerobic chemoorganotrophic bacterium able to denitrify and also remove phosphorus (Shi and Lee, 2007). The last band in Table 2 was found to be Thauera phenylacetica, and it was detected in the HR on day 113. This bacterium is a nitrate-reducing bacterium capable of anaerobically degrading methoxylated aromatic compounds with a bacterium from the genus Clostridium (Mechichi et al., 2005). The Thauera strain was found to use metabolic intermediates such as protocatechuate, acetate, and butyrate as a carbon source anaerobically or aerobically in the presence of nitrate. In SAMBR30, no bands were seen on day 85 because of a failed PCR. On day 113, DNA was detected on the agarose gel but only as a faint band, and no band could be seen on the DGGE gel, probably due to insufficient amounts of DNA. Nonetheless, bands were observed on the biofilm developed on the membrane of SAMBR30 (Lane day 115), which suggests that some of the degradation occurring in SAMBR30 toward the end of the experiment was probably due to the bacteria attached to the biofilm. However, it should be borne in mind that differences between lanes in the gel may be the result of different amounts of DNA loaded in each well.
The most diverse population was observed on day 64 in SAMBR300 with >20 bands, which contrasts with SAMBR30 where the low SRT did not allow the bacteria to grow in sufficient numbers. Indeed, this was associated in SAMBR30 with a persistently high SCOD (>2 g/L) in the bulk compared with values in the range of 400–1000 mg/L for SAMBR300 (Fig. 2). Nonetheless, both SAMBRs achieved about 95% SCOD removal at 35°C and a HRT of 1.1–1.5 days because of the retention of SCOD by the cake/gel layer on the membrane that provided a similar permeate SCOD for both SAMBRs. The population changed dramatically when SAMBR300 was operated at 20°C on day 100. Band 4 was found to have a very similar sequence to the one of a member of the genus Anaerovorax, which is known to be important for the digestion of polysaccharides and oligosaccharides (Krause et al., 2008).
Band 11 was found to have a very similar sequence to a member of the Syntrophaceae group, which includes propionate-utilizing bacteria (Ariesyady et al., 2007). Propionate was found to build up frequently in the HR in previous experiments (Trzcinski and Stuckey, 2009). Therefore, the presence of this bacterium was associated with stability regarding propionate degradation in this experiment.
Band 5 was found to have a very similar sequence to a member of the phylum Spirochaetes also detected by McHugh et al. (2004). The sequence from bands 7 and 8 was found to be highly similar to the sequence of Ralstonia species that are reported to be hydrogen-oxidizing bacteria (Song et al., 2002). Among the other species detected in SAMBR300, it is worth highlighting the presence of a sulfate-reducing bacterium in band 2 on day 64 only, but no attempt was made in this study to measure H2S to confirm sulfate reduction. The temporary appearance of this bacterium strongly suggests that sulfate reduction was not important in the SAMBR, and there was no competition with methanogens for the carbon source. Moreover, its presence can be related to the inoculum used in the HR (from Mogden wastewater treatment plant, United Kingdom) which also contained a band at that position and is referred to as MS in Fig. 4. In the HR, band 17 was identified as Uncultured bacterium clone G-25, which was detected in a study where lactate and sulfate were degraded in a continuously stirred tank reactor in which acidogenic and sulfate-reducing bacteria were detected (Zhao et al., 2008). The presence of this bacterium in our system suggests that sulfate-reducing bacteria could have also been present in the HR as they were in SAMBR300 on day 64 (band 2 in Table 2). However, the concentration of sulfate remained in the range 30–40 mg/L in the HR and SAMBRs effluents showing that no significant sulfate reduction was taking place in the process.
A bacterium of the genus Hydrogenophaga was also found to have a very similar sequence to the one extracted from band 6 of SAMBR300. Some Hydrogenophaga species are chemolithotrophic bacteria at the expense of hydrogen or carbon dioxide (Willems, 1991). Normally, methanogens would consume hydrogen and CO2 to produce methane according to the following reaction (Mosey, 1983; de Bok et al., 2004):
Alternatively, hydrogen could be utilized by hydrogen-consuming acetogenic bacteria such as Clostridium aceticum according to the following reaction:
However, in this study, it was found that other bacteria (99% similarity with Hydrogenophaga and a sulfate-reducing bacterium) might have used hydrogen as a source of energy (electron donor). Morvan and co-workers (1994) have shown that the rumen ecosystem can harbor other hydrogenotrophic populations such as H2-utilizing sulfate reducers before the apparition of methanogens in the newborn lamb, and this underlines the competition that exists between different H2-utilizing species. Methanogens and sulfate-reducing bacteria have a similar affinity for hydrogen (33 for some methanogens and 10 ppm, respectively) as pointed out by Morvan et al. (1994). However, in our study, it is likely that methanogens outcompeted sulfate-reducing bacteria because of the residual 30–40 mg/L sulfate detected in the SAMBR. Uchida et al. (2009) also reported their presence in landfill leachate, as they are known to play significant roles in the biodegradation of aromatic compounds, in denitrification, and in the bioremediation of contaminated environments.
Conclusions
An experiment with a wet two-stage lab-scale membrane bioprocess was performed to investigate its microbial population using the DGGE technique, and it was found that archaeal populations were different in both stages of the two-stage anaerobic process with Methanosarcina as dominant species. The SRT in the SAMBR significantly affected the archaeal diversity in the bulk, but a low diversity did not impede VFA degradation. An increase in VFAs concentration was observed in SAMBR30 at 20°C; whereas in SAMBR300, VFAs build up only appeared at 10°C. The dominant bacterial species in the HR belonged to Prevotella and Thauera; whereas the dominant ones in SAMBR300 belonged to Sphingobacteriales, Anaerovorax, Spirochaetaceae, Hydrogenophaga, Ralstonia, Prevotella, and Smithella. The low bacterial diversity in SAMBR30 compared with SAMBR300 resulted in a persistently high SCOD (>2 g/L) in the bulk reactor phase due to insufficient residence time for the bacteria to degrade recalcitrant COD. The presence of a member of the genus Hydrogenophaga in SAMBR300 suggests that hydrogen was found in the SAMBR.
Footnotes
Acknowledgments
This research was supported by a grant from the Department of Environment, Food, and Rural Affairs in the United Kingdom. The authors would like to thank Dr. Michael J. Ray for his assistance during the DGGE analysis.
Author Disclosure Statement
The authors declare that no competing financial conflicts exist.