
Abstract
Abstract
Two species of perennial grasses—Lolium perenne var. Top One (LPT) and Lolium perenne var. respect (LPR)—and a microbial consortium composed of two denitrifying polyphosphate-accumulating organisms (DPAOs)—strain Bacillus sp. MOE1 and Microbacterium sp. MOE2—were used to study the effect of plant–microorganisms interaction on removing nutrients from eutrophic water. The two perennial grasses could greatly remove the nutrients and pollutants; LPT could remove up to 40% of total nitrogen (TN) and 87% of TP in 20 days. Among the two species of perennial grasses, LPT is more efficient in removal of ammonium nitrogen (NH4+-N), phosphorus, and CODMn. Without combining with LPT and LPR, the immobilized DPAOs could remove more than 73% phosphorus, 32% total nitrogen, and 81% ammonium nitrogen. Removal efficiency of phosphorus, TN, and NH4+-N enhanced after combining DPAOs with perennial grasses, especially with LPT. The combination of DPAOs and LPT may prove to be a promising approach in bioremediation of eutrophic water. In addition, the activities of urease, invertase, and alkaline phosphatases positively changed with the removal of nitrogen, chemical oxygen demand (COD), and phosphorus correspondingly.
Introduction
Bioremediation is a rapidly changing and expanding area of environmental biotechnology (Tomotada and Masao, 2001), using the metabolic potential of plants or microorganisms to clean up contaminated environments, which contain a variety of organisms. Of these, bacteria, such as those capable of degrading PAHs, PCBs, and other pollutants (Dercova et al., 2008; Pathmalal et al., 2009; Liu et al., 2010), usually have prominent roles, whereas other organisms, such as plants (Jan and Lenka, 2005; Ana et al., 2008), also affect the process (Watanabe, 2001).
Ecological engineering could repair the environment without damaging the former ecosystem cost-effectively, and it is an emerging field dedicated to the design and construction of sustainable ecosystems that provide a balance of natural and human values (Mitsch et al., 2002; El-Shafai et al., 2007; William et al., 2009). The ecological engineering for wastewater treatment has been applied nationwide in China because of its advantages (Lu et al., 2008; Zhou et al., 2009).
As an indispensable component for ecological engineering, aquatic macrophytes can be used to remove nutrients and heavy metals (Hou et al., 2007; Maine et al., 2009), by consuming them in the form of plant nutrients (Elifantz and Tel-Or, 2002). Though many researchers have investigated nutrients removal capacities by different aquatic plants (Coleman et al., 2001; Lauchlan et al., 2004; Ana et al., 2008), the utilization of aquatic mcrophytes is limited in some aspects (Li et al., 2007). Some plants such as Eichhornia crassipes and Pistia stratiotes Linn. are considered to be notorious weeds in freshwater ecosystems; even though they have significant nutrients removal capacity, some are dependent on a few plant species for colonization of the water surface, causing sudden breakdown of eco-population (Vaillant et al., 2002). The applications of these plants were limited by these shortages. Therefore, selecting plant species with good nutrients removal efficiency and easy-to-deal plant residues are of great importance.
Although the key role of microorganisms in nutrients removal was realized, little is known of microbial community versus environmental regulation of enzyme activity (Sinsabaugh et al., 2005). Besides, despite the consensus that microbial communities play the leading role in treatment processes in plant floating-bed system, how the plant induces and affects microorganisms, as well as the combined effects of plant and additive microorganisms on nutrients removal, remains unclear.
To better understand the relationship between enzyme activities and nutrients removal efficiency, three enzymes that involved in the hydrolysis or transformation of nitrogen, phosphorus, and organic material in eutrophic water were measured. Therefore, the main objectives of the present study were to (1) evaluate the ability of nutrients removal of two species of perennial grasses, (2) investigate the relation between enzyme activities and nutrients removal in perennial grasses-immobilized microorganisms (IMs) integrated system, and (3) obtain a possible understanding of the mechanism of plants and microorganisms in nutrients removal.
Experimental Methods
Preparation of microorganisms
The isolation and identification of two strains of denitrifying polyphosphate-accumulating organisms (Zeng et al., 2003) were carried out according to Shi and Lee (2006). The characterization of isolated strains (Bacillus sp. MOE1 and Microbacterium sp. MOE2) and subsequent studies are to be yet published (Li et al., 2011).
Plant culture
Two species of perennial grasses Lolium perenne. var. Top One (LPT) and Lolium perenne var. respect (LPR; Fig. 1), supplied by Beijing Clover Seed and Turf Co. China (Beijing, China), were artificially germinated. When sprouted, they were transferred onto floating beds. Beds were made of polyethylene foam with 77 cm length, on which there were 2-mm-diameter holes in every 4 mm. The roots of plants could reach water by holes. The floating beds were daily irrigated with distilled water to compensate evaporation and transpiration loss.

The picture of LPT
Experimental design
The eutrophic water was sampled from Huajiachi pool (located in Zhejiang University campus), and the chemical characteristics of the water are TN 2.99 mg/L, TP 0.56 mg/L, NH4+-N 1.46 mg/L, NO3−-N 1.10 mg/L, CODMn 28.12 mg/L, and pH 7.95.
The study included six treatments, with three replicates for each: (1) water with the same floating bed but without plant as control (CK), (2) water inoculated with IMs, (3) water introduced with LPT, (4) water introduced with IMs and LPT (LPT+IM), (5) water with LPR, and (6) water introduced with IMs and LPR (LPR+IM).
The experiment was carried out in Zhejiang University during October 2008 to April 2009. The wastewater was filled into 75-L plastic containers with diameter of 80 cm and height of 100 cm. As long as the plant seedling was 5 cm long, they were transferred to the experimental pond in Zhejiang University for seedling establishment for 10 days. Plants growing in floating beds with similar growth performance were thoroughly washed with distilled water before they were transferred into the containers mentioned earlier, covering about 95% of the water surface. The control was designed to cover the container with the same floating bed but without plant. Distilled water was supplied everyday to compensate the water loss. The nutrients removal experiment was carried out under natural light and the temperature varied between 20°C and 25°C during the experiment; the steps involved in this research are shown in Fig. 2.
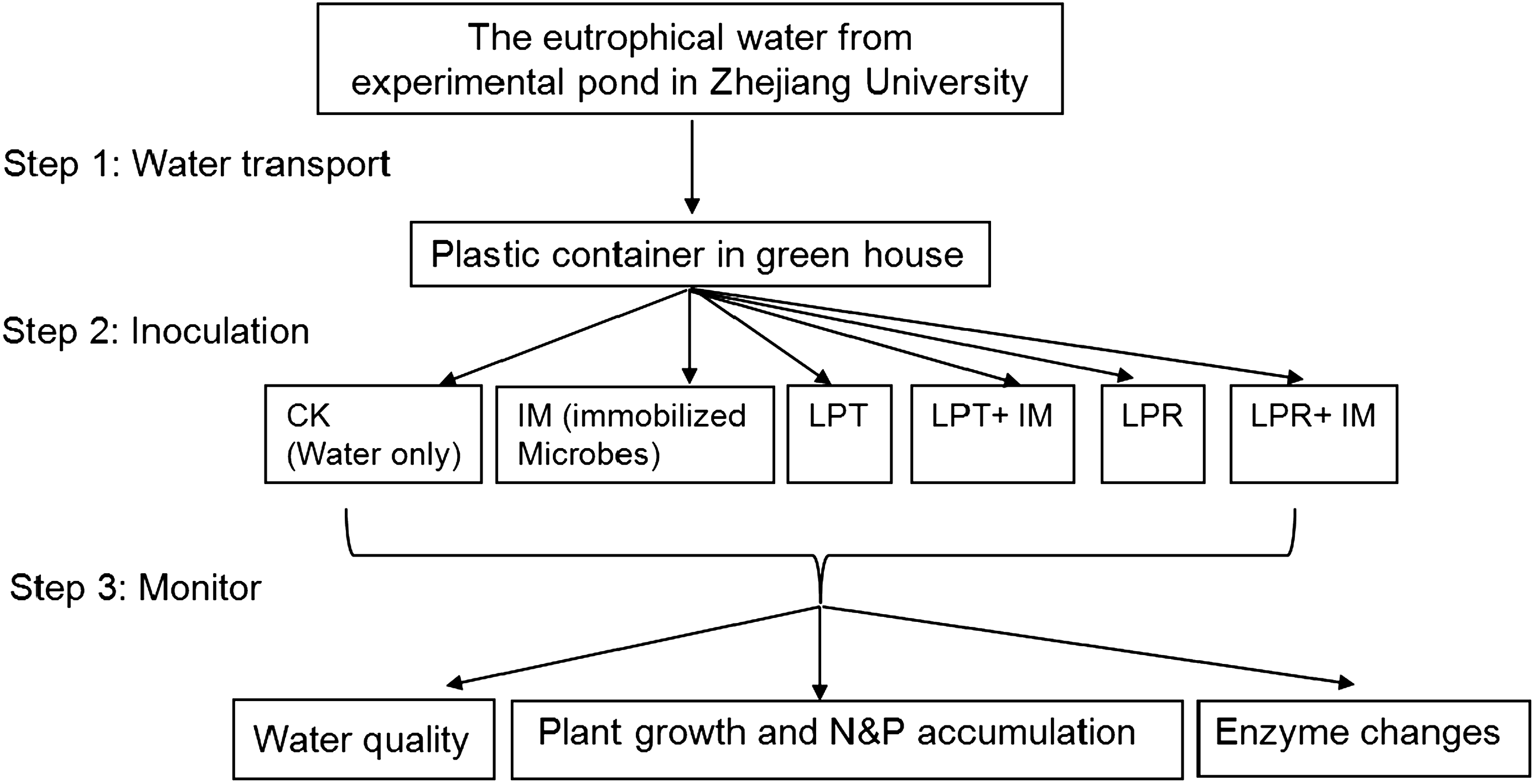
Experimental setup. The investigation was progressed in three steps: water transportation, inoculation, and monitor; the test was carried out during October 2008 to April 2009. Grasses here were artificially germinated, followed by transferring to floating beds. IMs, immobilized microorganisms, CK, without plants or exogenous microorganism; N and P, nitrogen and phosphorus.
Sample collection and physicochemical analysis
We collected samples of each treatment by carefully removing plants from the floating beds every 5 days. After washing with tap water, samples were blotted with absorbing paper and their fresh weights were measured. The initial plant weights were measured as start point control. Plant dry weights were measured upon reaching constant weight, after drying at 70°C for 24 h. Chemical oxygen demand (COD), total nitrogen (TN), total phosphorus (TP), ammonium nitrogen (NH4+-N), and nitrate nitrogen (NO3−-N) of water samples were measured according to a former report (Vaillant et al., 2003). TN and TP of plant materials were processed and measured according to methods described by Wang (2004).
Measurements of enzyme activity
Before measuring enzyme activities, we condensed 250 mL water sample to 10 mL by a rotary vacuum evaporator, and assays were performed in triplicate and corrected for a blank.
We carried out the phosphatase assay referring Livia's method (Livia and Uwe, 2005): 10 mL of condensed water sample was mixed with 4 mL of modified universal buffer (pH 11.0), 0.25 mL of toluene, and 1 mL of 25 mM p-nitrophenyl phosphate, and then the solution was incubated at 37°C for 1 h. The reaction was terminated by adding 4 mL of 0.5 M NaOH and the formation of p-nitrophenol was determined at 400 λm with UV-2450 spectrophotometer (Shimadzu, Japan) and enzyme activity was defined as μg p-nitrophenol·100 mλ−1 water·h−1.
We carried out urease activity according to Klose's method (Klose and Tabatabai, 2000). A solution consisting of 10 mL of phosphate buffer (pH 6.7), 0.5 mL of toluene, and 10 mL of 10% aqueous urea was added to 10 mL of condensed water sample and then incubated at 37°C for 48 h. About 20 mL of 1 M KCl was added after incubation, and the solution was thoroughly shaken for 30 min and filtered to use. About 1 mL of filtrate was mixed with 1 mL potassium sodium tartrate, 0.8 mL Nessler reagent, and 4 mL of 1 M NaOH and filled up to 25 mL with distilled water. The released NH4+ was determined at 460 λm using a UV-2450 (Shimadzu, Japan) and the urease activity was defined as μg NH4+ 100 mL−1 water 24 h−1.
We carried out the invertase assay following the method of Michael and Li (Michael and Martin, 1999; Li and Luo, 2008): 10 mL of condensed water sample was mixed with 5 mL of 20% saccharose solution, 5 mL of phosphate buffer (pH 5.5), and 0.25 mL of toluene, and the solution was incubated at 37°C for 23 h and then reacted by shaking in a water bath at 37°C for 1 h. Later reaction was immediately filtered and mixed with 5 mL of Fehling regent, 2 mL of 33% KI, and 1.5 mL H2SO4, filled up with distilled water to 100 mL, and titrated with 0.1 M of Na2S2O3, adding 0.5 mL starch indicator before titration. The units of invertase activity were defined as mL of sodium thiosulfate consumed by 100 mL water at 37°C per 24 h.
Data analysis
Statistical analysis of data was performed using Sigma Plot (version 10.0) and was compared with least significant difference test at 0.05 level using Statistical Package for Social Science (SPSS, version 16.0). Data were expressed as the mean±standard error.
Results and Discussion
Effects of plant–microorganisms combination on nutrient removal
NH4+-N removal had a significant difference among treatments. With LPT treatment, the NH4+-N was removed by 62.8%, which showed significant difference with that of LPR (at p=5% of probability), and when IMs were introduced, the removal efficiency reached 71.3%, which was much more remarkable than other treatments. It is peculiar that with IM treatment, the NH4+-N removal reached 81.3%, which was considerably high than other treatment, especially that combined with plants (Fig. 3). With IM treatment, there were no exogenous ammonia nitrogen supplement to the water, but perennial grasses residues may make up the NH4+-N removed from the water body and thus offset its elimination effect.
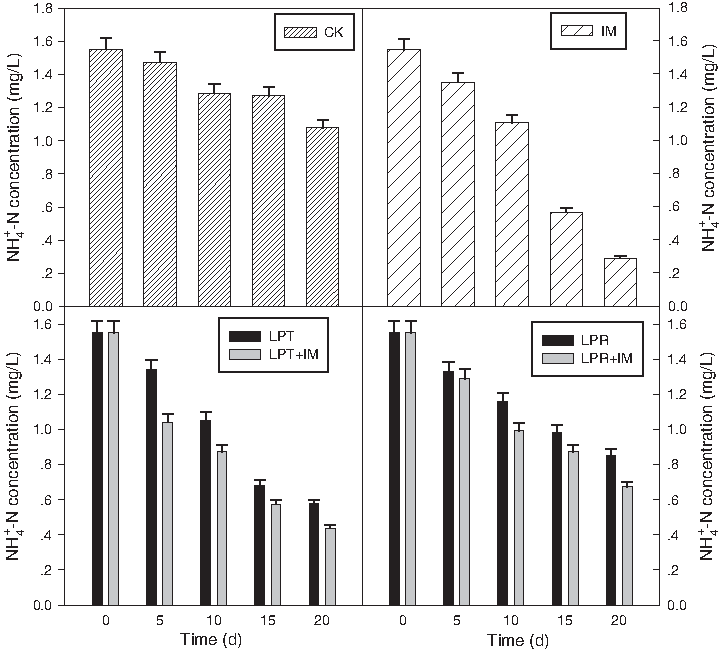
NH4+-N removal during phytoremediation with different treatments.
Similar behavior was observed for the removal of NO3−-N. When the eutrophic water was treated with perennial grasses, the NO3−-N removal efficiency was 52.6% and 45.6% with LPT and LPR treatments, respectively (Fig. 4). On day 10, the NO3−-N was higher in control pot than others, probably because of the fact that the development of an oxygenated zone around the roots would support the growth of nitrifying bacteria, increasing the supply of NO3− for plant uptake (Münch et al., 2005).

NO3−-N removal during phytoremediation with different treatments.
When IMs were added, the situations were quite different and the removal efficiency was 55.4% and 46.9% for LPT and LPR (Fig. 4). Among the two species of perennial grasses, LPT had a more prominent capacity to remove NO3−-N, however, the trend changed when IMs were introduced. When the IMs were added to control pot, the removal efficiency increased by more than 200%, which was the most significant change among all the treatments. The results indicated that plants were not the leading role in the removal of nitrate nitrogen, because when microorganisms were introduced, the removal efficiency sharply increased, whereas the nitrate nitrogen content did not significantly decrease with plant treatment individually.
Nitrogen removal was evaluated and TN removal profiles from the eutrophic water with these cultivars and IM treatment are shown in Fig. 5. In this study, TN decreased over time in both experimental and control sets, but nitrogen removal was significantly higher in the planted system. The removal efficiency was found to be in the order of LPT>LPR, of which the removal efficiencies were 55.6% and 40.1%. During the 20 days of culture, control pots without plants or microbial consortium showed unnegligible nitrogen removal, mostly because of the self-purification of waterbody. The maximum TN removal rate was attained at 20 days for LPT combined with IM treatment (59.8% removed); this value was 3.6 times higher than that obtained from control (16.4%). It is evident that the treatment with plants was more efficient when compared with the control, and the increase in efficiency becomes more evident with the addition of microorganisms, for instances, the plant+IM treatment decreased TN concentration to 59.8% and 45.3% for LPT and LPR, respectively, and the ability of IM to remove TN in the tested water was 32.1%. It can be evaluated that significant differences exist between LPT and LPT+IM (at p=5% of probability) as well as LPR and LPR+IM (Fig. 5). As nitrogen removal is primarily dependent on the nitrification/denitrification activity of root-associated bacteria (Farahbakhshazad and Morrison, 1997), the presence of aquatic macrophytes and their associated microorganisms can greatly enhance the ability of the floating-bed system to reduce nitrogen levels.
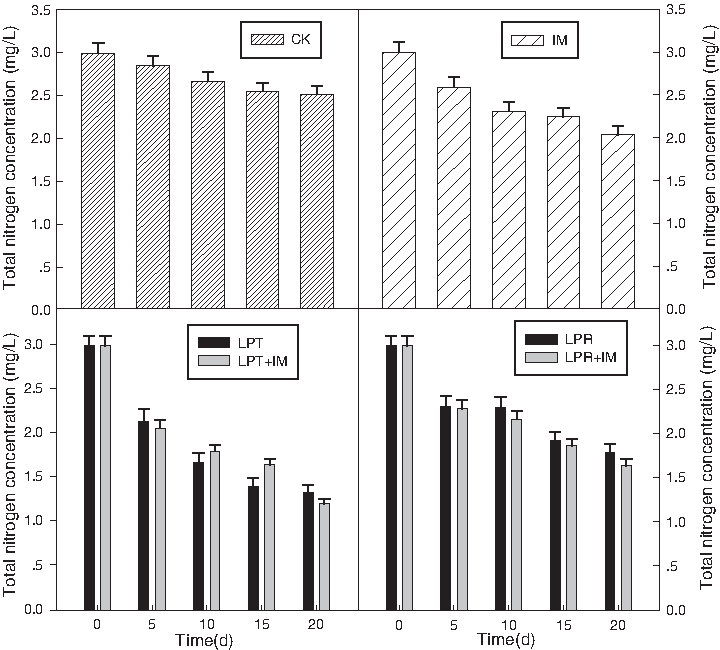
Total nitrogen removal during phytoremediation with different treatment.
The use of IM to purify water is influenced by many factors. After IMs are introduced to the water, plant rhizosphere environment is changed, which could influence the biological activity of these microorganisms and thus influence removal rate (Jiang et al., 2004). On one hand, the transport of O2 from leaves and stems to roots along with the associated rhizosphere microorganisms is important for nutrient removal (Good and Patrick, 1987), on the other hand, oxygen transport can be particularly important in the denitrification process, because it is required for nitrate formation, whereas nitrate is the limiting factor for denitrification. As nitrification occurs in aerobiotic environments and denitrification in anaerobic environments, the juxtaposition of the oxidized zone around the root with the reduced environment of nonrhizosphere zone is very important for the treatment of N in plant–microorganisms system (Hunt et al., 2002). However, the removal efficiency was not so good in the later period of the study, which may be due to the fact that perennial grasses growth was suppressed when the temperature was not feasible for its growth, and consequently, plant root activity was depressed. Kazunoti et al. (1995) also reported that phosphate uptake rate and the amount taken up were higher at 15°C and 25°C than at 5°C and 35°C.
The results from the work by Tanner et al. (1999) showed that phosphorus can be removed by sorption, complexation, precipitation, and assimilation into microbial and plant biomass. In the present study, TP was not completely removed by the system, but the average TP removal in the planted system, especially combined with IM, was higher than in the unplanted system. In addition to high removal efficiencies of TP, the two species of perennial grasses also had a quite good capacity to remove nitrogen, as well as perennial grasses+IM integrated system. However, phosphate removal with IM treatment was also very good when it was not kept in anaerobic condition. During the experiment, it removed more than 73.2% phosphate after 20 days (Fig. 6) with IM treatment—this response was similar to the results reported by Gopal (1999).

Total phosphorous removal during phytoremediation with different treatments.
The more effective perennial grass to remove phosphate was LPT. On day 20, the TP level dropped from its initial 0.56 mg/L level to 0.47 mg/L (15.5% removal) in the control and from 0.56 mg/L down to 0.07 mg/L (87.1% removal) in the presence of LPT.
When LPT was put together with IM, the situation was quite different. In the first 15 days, the phosphate removal efficiency reached more than 80%. Considering the aforementioned results, it is important to emphasize that the maximum phosphorus removal takes place not with IM treatment but with IM and LPT, suggesting that phosphorus removal could be associated to the root exudates of the plant.
The experiment was designed to cover the container with a floating bed, which cut off the oxygen transportation from air to the eutrophic water; on the other hand, organic pollutant degradation consumed oxygen as well, leading to the anaerobic condition in early period. When the plant had adapted the aquatic environment, the transport of O2 from leaves and stems to roots was enhanced, combined with oxygen exudation from plant root; as a result, aerobic condition was created and the anaerobic and aerobic conditions enhanced phosphorus-accumulating microorganisms' activity.
CODMn was another pollutant for water eutrophication, although it is inferior to nitrogen and phosphorus. Among the two species of perennial grasses, LPT as well as LPT+IM had much prominent removal efficiencies than LPR (Fig. 7). The decrease of COD in the control pot, and in the presence of LPT, COD increased from 11% (on day 5) to approximately 31.2% (on day 20) and from 48% (on day 5) to approximately 66.8% (on day 20), respectively. When IMs were added, the removal was more significant. In the first 20 days, the CODMn removal rate had reached more than 66% and 70% with LPT treatment and LPT+IM, respectively. However, in the last 5 days, the removal efficiencies slowed down. This indicated that when IMs were added, the immobilized beads as well as plant root absorbed organic pollutant and thus biofilm was formed, on which a large amount of microorganisms inhabited. As a result, the pollutant removal efficiency was greatly improved (Henze, 1996), but with increase in absorption, the biofilm aged and abscission occurred, making pollutants removal efficiency discounted.
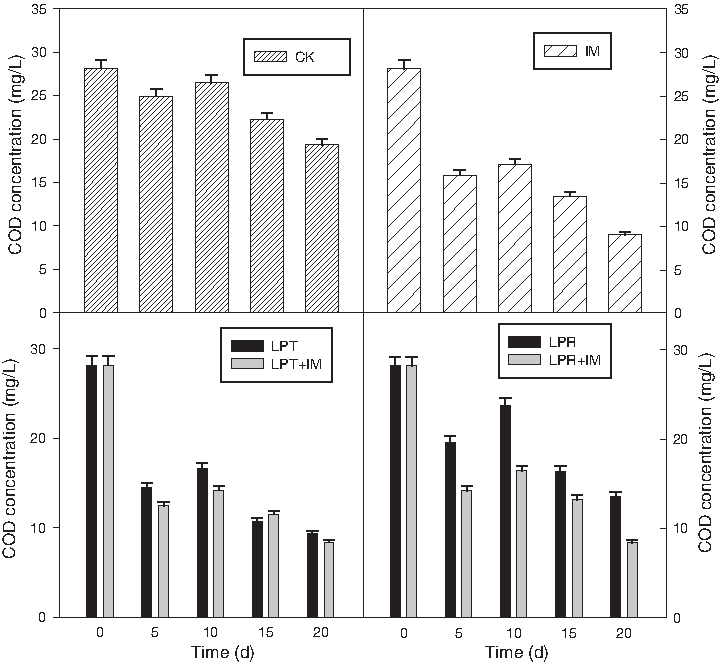
CODMn removal during phytoremediation with different treatments.
It has been reported that the utilization of aquatic macrophytes to remove nutrient from wastewater is one feasible phytoremediation way (Jayaweera and Kasturiarachchi, 2004; Li et al., 2007). However, most researchers consider that the potential removal of N and P by plants depends upon their growth rate and tissue N and P content (Li et al., 2007), but the function of some relevant microorganisms was ignored. Some researchers found that water collected close to the roots of Nymphaea amazonum, a species of aquatic macrophytes, had the highest fungal growth (Josy et al., 2009), but their role was unclear. Our study further revealed that microorganisms played the leading role in nutrients removal and plant uptake of some nutrients for its growth, on one hand, and simultaneously creating a proper environment for microorganisms, on the other hand.
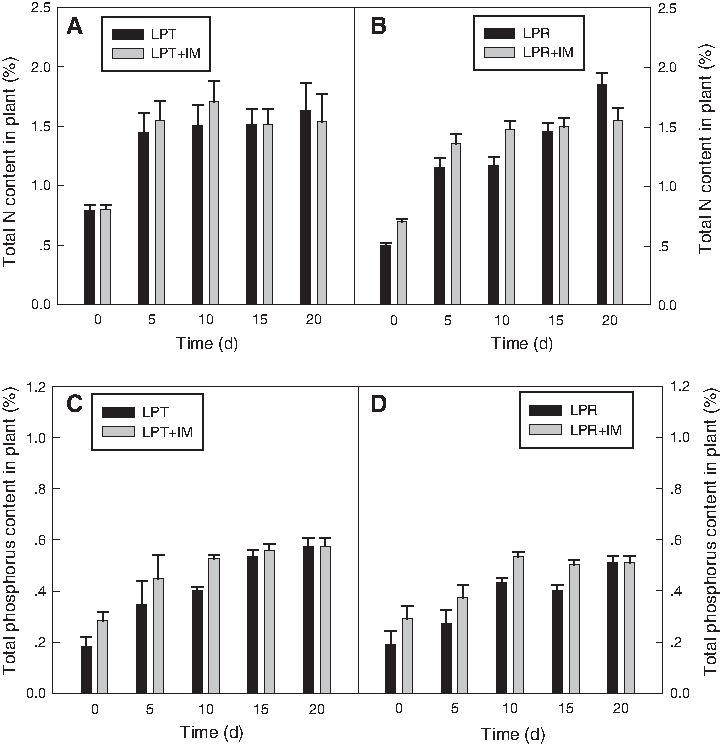
Variation of tissue nitrogen and phosphorus contents
Nitrogen and phosphorus accumulation by these grasses under different treatments during the study was estimated. It can be found that the addition of the microbial consortium had little influence on nitrogen uptake, especially in the early period (Fig. 8A, B), suggesting that the denitrification by microorganisms resulted in the loss of nitrogen in the system.
Total nitrogen
Maximal plant phosphorus content was obtained in LPT (Fig. 8C, D). Plant phosphorus content was enhanced by adding the microbial inoculums for both LPT and LPR, especially in the first 10 days of the present study; however, the rate slowed down as the research proceeded (days 15–20). These results show that the addition of the microbial consortium enhanced phosphorus removal from water, resulting in the deficiency of it for the plant in the later period. Therefore, a positive effect of inoculums was observed on the phosphorus removal and phosphorus accumulation in the early period, whereas a negative effect was found on plant growth later.
Aquatic macrophytes played an important role in constructed wetlands; however, the utilization of them is limited in some aspects. E. crassipes and P. stratiotes Linn. are the last choice in bioremediation, although they had significant nutrients removal capacity; some plants suffered from the embarrassment of plant residues. The utilization of perennial grasses showed great advantages such as high nutrient removal efficiency and convenient residue disposal (disposed as animal feeds) (Li et al., 2011).
Enzyme activity
Some research reported that enzymes in the wetland system played important roles in material transformation (Kang and Freeman, 1998). Few researchers have focused on substrate microorganism and enzymatic activities and there remains a lack of information about the relationship between microorganisms and pollutants removal efficiency (Liang et al., 2003). The production of enzyme in constructed wetlands is due to the activities of substrate microorganisms, animals, and plant roots.
Urease was involved in the nitrogen cycle (Megharaj et al., 1999; Marcote et al., 2001). The activity of urease was minimum in the control pot and was higher with the plant treatment and further enhanced when IMs were added (Fig. 9). The increase of urease activity coincided with high nitrogen removal rates with time. This might indicate a positive effect of LPT and LPR on microbial growth and N assimilation. The changes of enzyme activities in the control plot may be due to the change of temperature and the source water quality (Zhou et al., 2005).
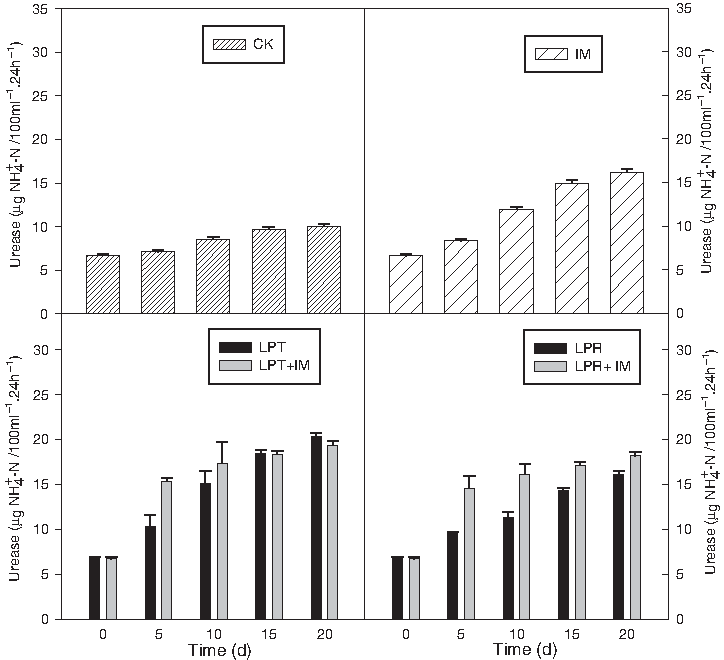
Urease activities of eutrophic water with different treatments, displayed as μg NH4+-N 100 mL−1 sample 24 h−1.
Conclusively, perennial grasses+IM had an obvious performance than either perennial grasses or IM individually. Among the two species of perennial grasses, LPT were more conspicuous than LPR in enhancing urease activity in rhizosphere.
The activity changes of invertase had a similar pattern as urease, which showed higher activity with plant and plant+IM treatment than control pot, especially with LPR (Fig. 10). The pattern of invertase activity increasing also showed difference between LPT+IM and LPR+IM, of which the activity showed a sharp increase at day 5 for LPT+IM, whereas a rapid increase appeared at day 20 with LPR+IM. In the end of the experiment, the growth of perennial grasses slowed down, and consequently, the CODMn removal was not remarkable compared with LPT and LPT+IM, as well as LPR.

Invertase activities of eutrophic water with different treatments, displayed as mL Na2S2O3·100 mλ−1 sample·24 h−1.
Phosphatase is responsible for hydrolyzing organic phosphorus compounds to inorganic P (Marcote et al., 2001). The relationship between phosphatase activity and soil phosphorus concentration has been shown in several studies (Allison et al., 2007). Kang and Freeman (1998) also indicated that soil enzyme activities are partly responsible for improving water quality. When the water was treated with IM, the activity of alkaline phosphatases significantly increased at day 5 and remained stable till the end. The alkaline phosphatases activity showed huge difference between treatments (Fig. 11). The most remarkable changes happened on LPT, of which the activity of alkaline phosphatases increased by 53.5% when combined with IM. The reason for this phenomenon might be because the root exudates enhanced the activity of microorganisms in rhizosphere. Dalal (1982) also reported that the addition of organic substances to the soil served as a carbon source, which enhanced microbial biomass and phosphatase activity. When treated with plants, the phosphatase activities were much higher because of ample nutrients and substrate for microorganisms.

Phosphatase activities in eutrophic water with different treatments, displayed as μg p-nitrophenol 100 mλ−1 sample h−1.
Summary
The present study demonstrated that concentration of residual TN and TP and the extent of COD remaining in eutrophic water in which two cultivars of perennial grass were planted were significantly lower than that without plants, indicating that it is effective for rapid removal of nutrients. Two strains of denitrifying phosphate-accumulating organisms, which were isolated and cultured by ourselves, acted well when they were immobilized and introduced to eutrophic water, especially combined with perennial grasses. LPT performed better than LPR either alone or combined with IM. The addition of these microorganisms improved plant growth and nutrients removal when compared with those not inoculated. TN, TP, as well as the extent of CODMn removal were higher in the presence of LPT and microorganisms than LPR.
Our results indicate that urease, phosphatase, and invertase play important roles in nitrogen, phosphorus, and organic matter removal in plant-floating bed system. However, how this microorganism acted in the nutrients removal process and how the microorganism community changed with perennial grasses treatment need further investigation.
Footnotes
Acknowledgments
This work was supported by the Key CN-USA International Cooperative Project (No. 2010DFB33960), a seed project for control and remediation of water pollution (No. 2008ZX07101-006-08-B-2) from the State Ministry of Science and Technology of China, and a project from Science and Technology Bureau of Zhejiang Province (No. 2010C13G2010132).
Author Disclosure Statement
No competing financial interests exist.