
Abstract
Abstract
This study is designated to examine the biological activity role of carbon cycling in a tropical lagoon, especially after a natural nutrient pulse. The study took place several days after precipitation. Both physical and chemical hydrographs indicated that significant amounts of dissolved inorganic nutrients had either entered the inner lagoon through runoff drainage or regenerated from the bottom sediments of the inner lagoon. Although the variables (chlorophyll a, bacterial biomass and production, as well as dissolved and particulate organic carbon) had orders of variation, linear relationships were significant between variables. This could have been associated with high phytoplankton growth enhanced by enriched nutrients and its sequential responses. This study considered both phytoplankton and bacterial communities that are the most important components of organic carbon consumption; they accounted for more than 87% of the organic carbon consumption in this ecosystem. A large ratio of primary production to community respiration with a mean of 1.33 suggested that this lagoon was autotrophic during the study period. It also indicated that there was a carbon sink in this ecosystem, and the residual carbon was either exported into the coastal sea or preserved in lagoon sediments resulting in the hypoxic condition of the bottom water. Faster water turnover rate was suggested to improve this low oxygen condition.
Introduction
Dapeng Bay is a tropical lagoon located on the southwestern coast of Taiwan. Previous studies showed this lagoon to be an autotrophic ecosystem, mainly based on a biogeochemical budget (Hung and Hung, 2003; Hung et al., 2008). The objective of this study was to examine the biological role–playing activity in carbon cycling in this lagoon, especially after a natural nutrient pulse. Several variables were measured simultaneously to understand how they associated with the carbon cycling processes in the lagoon. Community respiration (CR) was regressed with biological variables to discriminate the carbon consumption attributed to various components. The production to respiration ratio was used to explain the maintained carbon balance in this ecosystem.
Materials and Methods
Study area and sampling
The study site is located in the Dapeng Lagoon, southwestern Taiwan (22°27′N, 120°27′E), with an area of about 5.32 km2. This lagoon is a shallow (mean water depth of 2.2 m), semienclosed ecosystem with two openings: the seaward opening connected the lagoon to the Taiwan Strait and terrestrial Linpan Dike discharged urban and aquacultural wastewater into the inner lagoon. Samples were collected from 11 stations on June 21, 2000 (Fig. 1). Medium precipitation, 118.5 mm within 3 days, occurred several days prior to this study (Central Weather Bureau, Taiwan). At each station, water was sampled either from the surface or at three depth intervals (surface, middle, and bottom), depending on the water column depth. A custom-made pumping system consisting of a peristaltic pump and a precleaned silicone tube was used for water sample collection. Additional samples were collected from the Linpan Dike to analyze and trace for potential nutrient sources. Dissolved oxygen (DO) was measured in situ with a portable DO meter (YSI 52) calibrated by the direct spectrophotometry of total iodine method (Pai et al., 1993). Temperature and salinity were measured using a portable salinometer (WTW, KF-320) with salinity calibrated by Autosal (Hung and Hung, 2003).
Study area and sampling stations in the Dapeng Lagoon.
Nutrients, chlorophyll a, particulate organic carbon, and dissolved organic carbon
Water samples for chlorophyll a (Chl a) measurement were filtered through a GF/F filter. Concentration of Chl a was determined with fluorometry following acetone extraction (Parsons et al., 1984). For further analyses, a conversion factor of 50 for Carbon to Chl a was used (Chang et al., 2003). Water samples for nutrient, particulate organic carbon (POC), and dissolved organic carbon (DOC) analyses were collected in 4 L polypropylene bottles. Part of the water sample was filtered through a Whatman 25-mm GF/F filter, prebaked at 450°C for 4 h, for POC measurement. Filtered seawater was collected separately for nutrient and DOC analysis. After removal of the inorganic carbon with 2 M hydrochloric acid, the C/N/S analyzer (Fisons NSC 1500) determined POC. DOC was measured using the HTCO (Shimadzu TOC 5000A) method (Hung and Hung, 2003). Nutrients (i.e., NO3−, NO2−, NH4+, PO43−, and H4SiO4) were determined on a colorimeter equipped with flow injection analysis. The trapezoidal method was used to integrate the values of variables over the whole water column, if applied.
Bacterial abundance and production
The acridine orange epifluorescence microscopy method and 3H-thymidine incorporation measured bacterial abundance and production, respectively (Hobbie et al., 1977; Fuhrman and Azam, 1982). Bacterial biomass (BB) in carbon units was derived from bacterial abundance using a conversion factor of 20×10−15 g C·cell−1 (Lee and Fuhrman, 1987). Bacterial production (BP) was estimated with a thymidine conversion factor of 1.18×1018 cells mol−1 (Fuhrman and Azam, 1982).
Plankton CR and net primary production
Plankton CR and net primary production (PP) were measured as the change in DO in dark and light incubation, respectively. Dark treatments were performed for samples collected from all water depths in the experimented stations, and light incubations were conducted on samples from surface water at stations 5, 9, and 11. The direct spectrophotometry method measured concentration of DO (Pai et al., 1993). The difference in DO concentration between initial and dark (or light) treatment was used to compute CR (or net PP). To convert respiration and production from oxygen to carbon units, both the respiration quotation and the photosynthetic quotation of 1 were applied. To estimate PP (mg C·m−3·d−1) in the surface water of other stations, an empirical relationship between PP and salinity (Sal) was applied as follows:
with r2 and p linear regression values of 0.99 and 0.015. The estimated PP results were used in this study.
Results and Discussion
Nutrient sources of the semienclosed ecosystem
The Dapeng Lagoon has one seaward opening, and one terrestrial water input via the Linpan Dike into the inner lagoon (Fig. 1). In the seaward end of the lagoon, hydrographic data showed high salinity and low temperature; the inner lagoon exhibited lower salinity and higher temperature (Figs. 2a, b and 3a, b). The water turnover rate was influenced mainly by higher freshwater input via precipitation (Hung and Hung, 2003) that occurred several days prior to this study. Evidence of this phenomenon was the low salinity or high temperature of the surface saltwater of the inner lagoon (Figs. 2a, b and 3a, b).
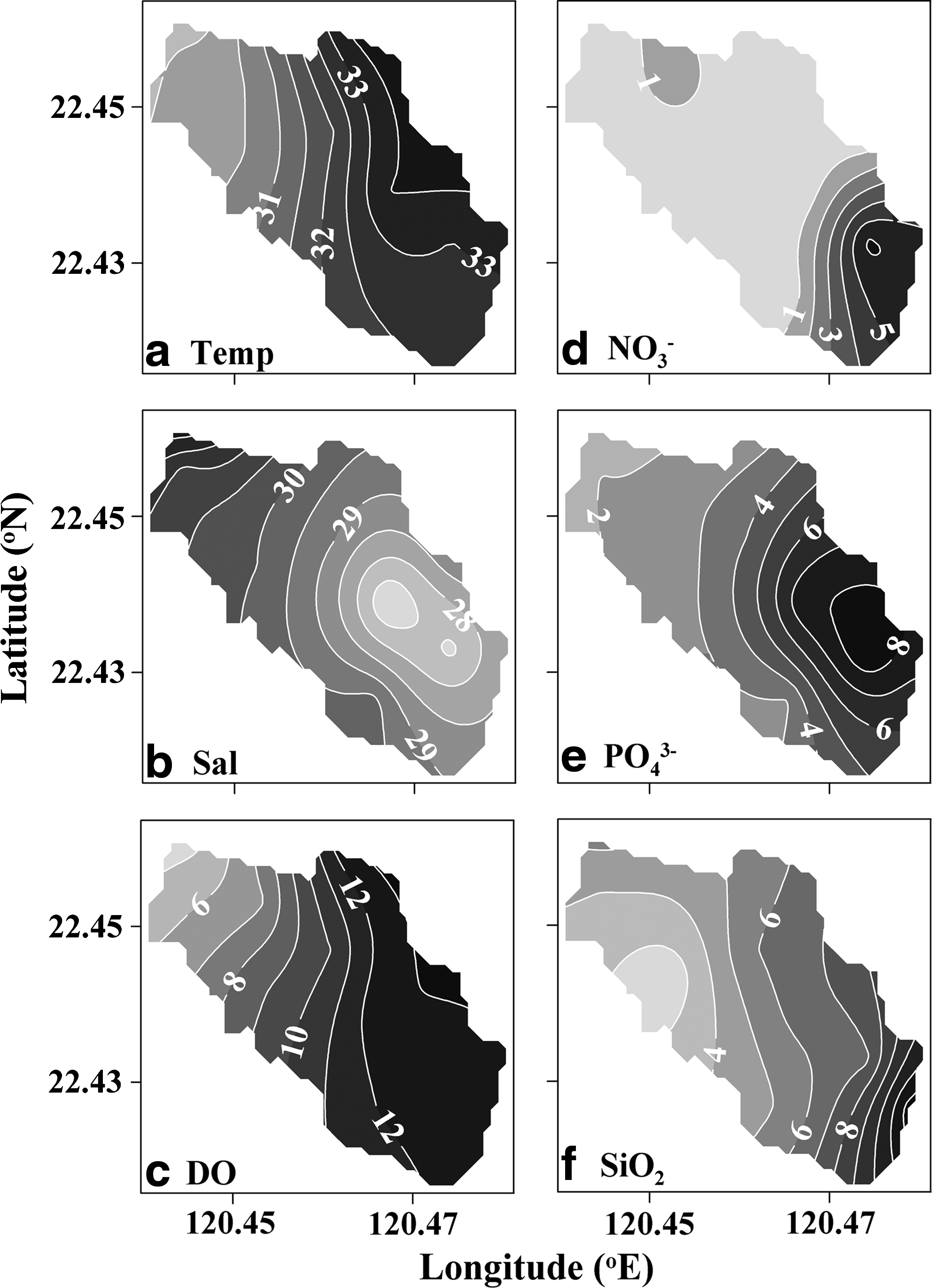
Contour plots of

High concentration of dissolved inorganic nutrients, including nitrate, phosphate, and silicate, was found in the surface water of the inner lagoon with values of 11.2, 9.0, and 12.3 μM, respectively (Fig. 2d–f). This could be attributed to nutrient input via the freshwater runoff from the Linpan Dike where high nutrient concentrations (i.e., nitrate=40.0 μM, phosphate=39.4 μM, and silicate=86.3 μM) were measured. Furthermore, this assumption was indirectly supported by the significant relationships between phosphate concentrations and salinity (r2=0.76, p<0.001) and phosphate concentration and temperature (r2=0.46, p=0.02) in surface water. These results indicate that one of the major nutrient sources of this lagoon was fluvial runoff from the Linpan Dike, especially after precipitation.
Besides freshwater runoff, the water–sediment interface might be another important source for nutrients diffusing through to the water column (Kemp and Boynton, 1984). In this shallow ecosystem, results showed a high concentration of ammonium (79.9 μM; data not shown), phosphate, and silicate in the bottom water of the inner lagoon (Fig. 3e, f). The nitrate patch, with a value of 6.19 μM, observed in the middle depth of the water column might be due to nitrification of ammonium from the bottom water (Fig. 3d). A high rate of nutrient regeneration in the sediment could explain the high nutrient concentrations observed in the bottom water; unfortunately, this study did not perform a diffusion rate measurement. One can infer that diffusion occurs from the low DO concentration (0.96 mg O2·L−1; Fig. 3c) and the high H2S concentration (2–14 mg·L−1) observed in the bottom water of the inner lagoon (Hung and Hung, 2003). These results suggest that nutrient regeneration in the sediment might be another important nutrient source for this shallow lagoon. Nutrient input from the ocean via the tidal inlet appears to be less important.
Spatial distribution and regulating factors of planktonic communities
Integrated values of biological and physical variables over the entire water column were used to examine the relation between variables. Several orders of variation were observed for the integrated values of Chl a, BB, BP, POC, and DOC (Fig. 4; Table 1). Even though the integrated values varied over a wide range, significant correlations were evident between the integrated variables (Table 2), indicating that distributions of biological variables are in accordance with physical and chemical hydrographs (Figs. 2 and 4).
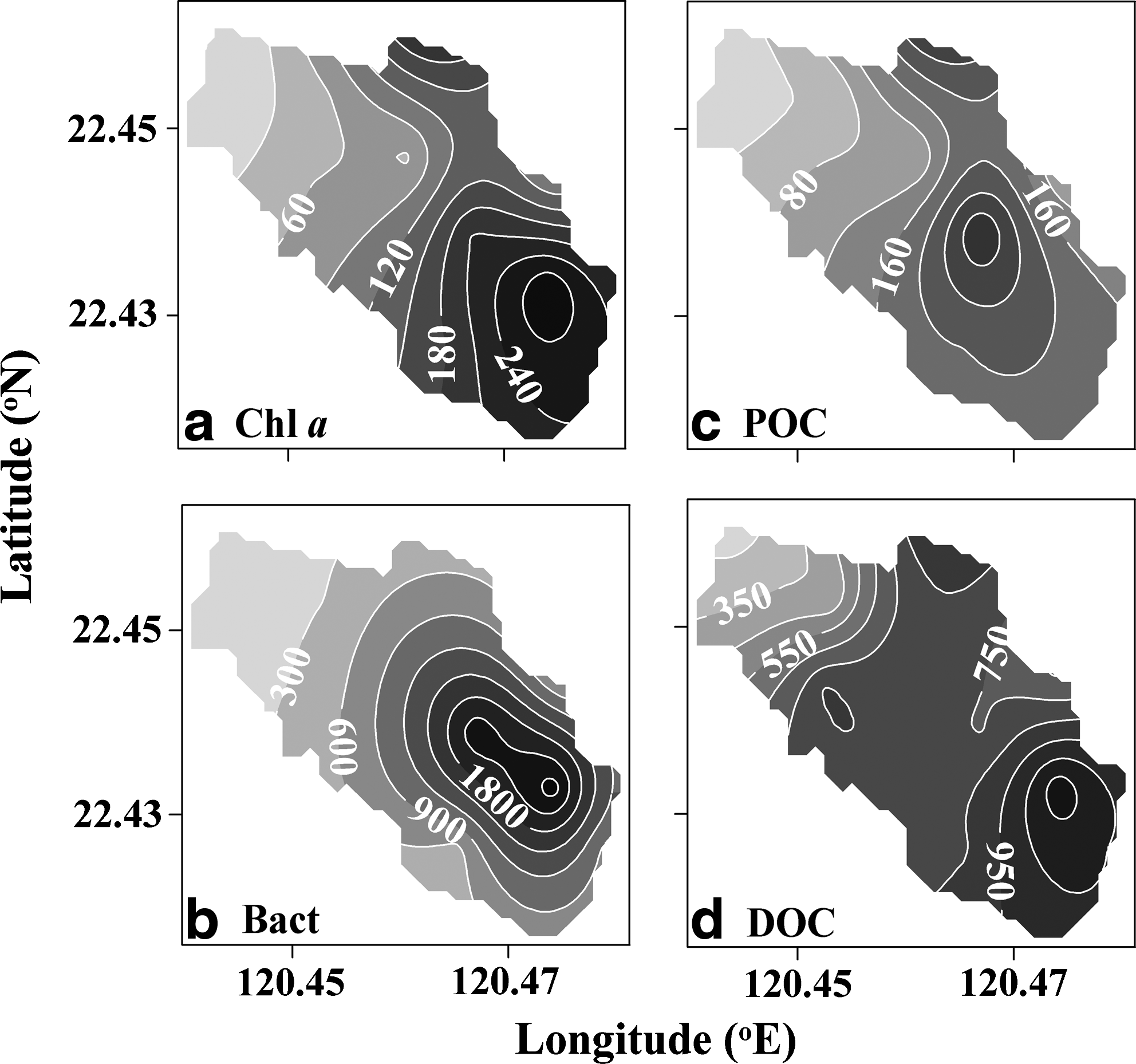
Contour plots of integrated values of
Values were integrated over whole water column depth for each variable.
POC, particulate organic carbon (mmol·m−2); DOC, dissolved organic carbon (mmol·m−2); Chl a, chlorophyll a (mg Chl·m−2); BB, bacterial biomass (mg C·m−2); BP, bacterial production (mg C·m−2·d−1); PP, primary production (mg C·m−2·d−1); ICR, integrated community respiration (mg C·m−2·d−1).
p≤0.01.
p≤0.05.
DIN, dissolved inorganic nitrogen;
As stated previously, the biomass of phytoplankton was highly variable in this small lagoon (Table 1). Spatially, the lowest values for the phytoplanktonic biomass were found in the seaward opening, with the values increasing toward the inner lagoon (Fig. 4a). The growth of phytoplankton can be regulated by any combination of light intensity, nutrient availability, and grazing pressure (Howarth, 1988; Kirk, 1994; Calbet and Landry, 2004). In this study, significant linear relationships were evident between integrated Chl a and the measured nutrients (Table 2). The data suggest the important nutrient impact on the spatial distribution of phytoplankton in this lagoon. Furthermore, molar ratios of nitrogen (i.e., dissolved inorganic nitrogen) to phosphate in the water column were low with a mean±standard deviation (SD) value of 2.23±2.23. This indicated that growth of phytoplankton in this lagoon might be limited by nitrogen, and this has been suggested in a previous study (Hung and Hung, 2003). The low nitrate concentrations (<1 μM) observed in most of the surface water could also support this assumption (Fig. 2d). In addition, light availability was another important factor regulating the growth of phytoplankton, especially in the inner lagoon, which had high Chl a concentration (Figs. 4a and 5a). Low light intensity could be attributed to a shading effect caused by a high concentration of plankton and DOC in the water column (Fig. 5a, c, d) (Kirk, 1994). Grazing by zooplankton seemed to be a minor factor in the regulation of the phytoplankton biomass, since a low grazing impact (3.5%) had been observed in this lagoon (Lo et al., 2004).
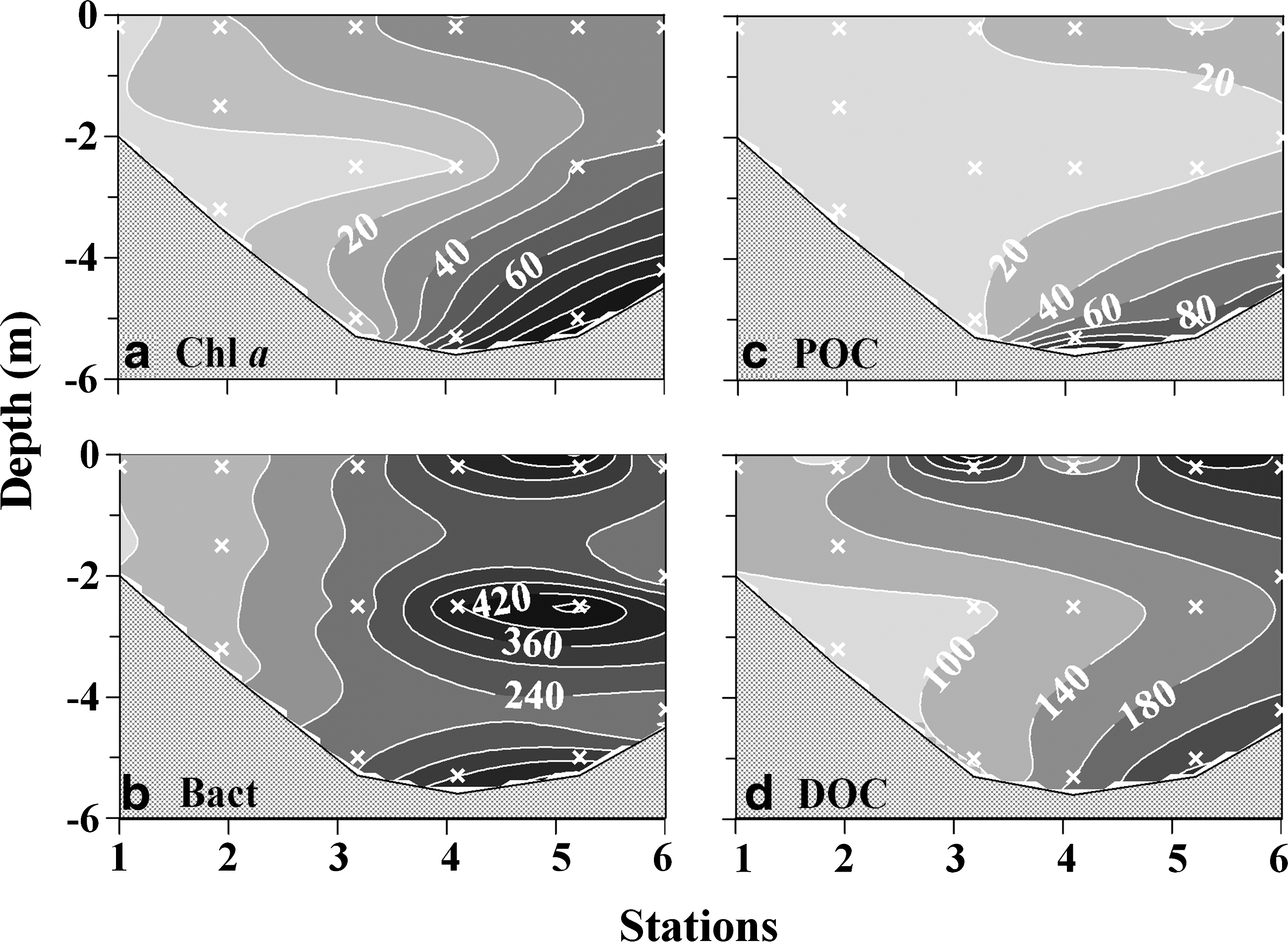
Depth profiles of
The spatial distribution of bacterioplankton was similar to that of phytoplankton (Fig. 4b). A similar trend was also evident in the linear relationship observed between BB and the integrated Chl a value (Table 2). This suggests that the growth of bacterioplankton is enhanced by a DOC flux associated with phytoplanktonic exudation or sloppy grazing of herbivores. DOC exudation via phytoplankton can be observed indirectly from the significant relationship observed between Chl a and DOC (Table 2). Integrated BB also regressed linearly with nutrients and BP (Table 2). All of these facts suggest that nutrients and organic carbon play important roles in regulating bacterioplankton, and that growth of bacterioplankton is most likely a bottom-up phenomenon in this lagoon.
Organic carbon consumption in the lagoon
In this study, a large variation in integrated CR (ICR) was observed, ranging from 818.6 to 4146.8 mg C·m−2·d−2 with a mean±SD value of 2243.3±986.1 mg C·m−2·d−2 (Table 1; Fig. 6a). Exploring further, the relationships between ICR and other biological variables provide evidence for understanding the planktonic components that are the most important contributors to organic carbon consumption. Different organisms might contribute CR, including phytoplankton, bacterioplankton, protozoa, and zooplankton, and the relative importance is system dependant (Biddanda et al., 1994; Chen et al., 2009). Indeed, significant linear relationships were evident between ICR and several integrated biological variables including Chl a, PP, and BB, as well as BP (Table 3). ICR also multiply regressed against the integrated values of Chl a and bacterioplankton, as represented by the following equation:
The same test was also conducted for average values (per m3) over the water column. Slopes, intercepts, and r2 values of linear regression are shown only for relationships with statistical significance.
p<0.01.
with an r2 value of 0.86 (p<0.001). However, significant linear relationships were observed only between CR and Chl a or PP (data expressed as average values; Table 3). In all cases, this suggests that organic carbon consumption was associated more with phytoplankton and, to some extent, with bacterioplankton in the lagoon. In this ecosystem, phytoplankton is responsible, on average, for about 53% of organic carbon consumption, assuming dark respiration is 40% of PP (Weger et al., 1989). In addition, bacterioplankton contributed about 34% of the CR if a 20% growth efficiency is assumed (del Giorgio et al., 1997). This should be an expected result for an ecosystem with a significant growth of phytoplankton induced by a large amount of nutrients. Thereafter, one should also expect the growth of bacterioplankton to be enhanced by high nutrient concentrations and by rich organic materials.
To understand the fate of organic carbon, the ratio of PP to CR (PP:ICR ratio) was used to examine whether there was a carbon sink or source in the lagoon (Chen et al., 2006). In this lagoon with high organic carbon production (≥2000 mg C·m−2·d−1; Table 1), ratios of PP:ICR were higher than 1 at almost all stations with a mean value of 1.33. A high PP:ICR ratio suggests the autotrophic highly productive ecosystem; a similar conclusion was derived from a biogeochemical model estimation (Hung and Hung, 2003). This evidence indicates a carbon sink in this ecosystem; Hung and Hung (2003) also estimated that the annual CO2 invasion from air to water is about 10.3 mol C·m−2·year−1 in this lagoon. The residual organic carbon is either exported through tidal exchange or preserved in the sediment. One can expect bottom hypoxia, therefore, in sediments with a high organic carbon content and low physical turbulence (Chen et al., 2007). Low DO concentration (Fig. 3c) and high H2S concentrations were observed in the bottom waters of the inner Dapeng Lagoon (Hung and Hung, 2003).
The results of the current study showed that significant amounts of nutrients were flushed into the lagoon from the fluvial input of the Dike after precipitation (Figs. 2 and 3). A large quantity of organic carbon was produced in situ via phytoplankton enhanced by enriched nutrients (Figs. 4 and 5; Table 1). A large ratio of PP to CR indicates a carbon sink in this ecosystem. The residual carbon could be preserved in lagoon sediments resulting in the hypoxic condition of the bottom water (Fig. 3c).
During the study, oyster culture racks and caged fishery structures largely occupied this lagoon; these maricultural structures were removed completely around January 2003. After the structures' removal, the net ecosystem production was even higher, and the low bottom DO water distributed widely in this lagoon (Hung et al., 2008). The slow water exchange rate has been suggested as the main factor influencing the carbon and nutrient biogeochemical processes and budgets of this lagoon (Chung et al., 2011). To manage and improve the water quality of this coastal lagoon, the authors suggest a faster water turnover rate or water circulation, especially in the inner lagoon.
Footnotes
Disclosure Statement
No competing financial interests exist.
Acknowledgments
This research was supported by the National Science Council, Taiwan, under contract Nos. NSC-89-2621-Z019-008 and NSC-89-2621-Z110-016. We would like to thank Dr. H.-M. Su at Tungkang Branch, Taiwan Fisheries Research Institute for generously providing facilities for conducting our incubation experiments. We also thank Miss P.-Y. Hung for her assistance with the analysis of nutrients, POC, and DOC samples. The authors are also grateful to five anonymous reviewers for providing valuable and constructive comments to improve the manuscript.