
Abstract
Abstract
This study investigated the influence of water hardness (Mg2+ and Ca2+) on the fate and toxicity of 20 nm citrate silver nanoparticles (AgNPs) and Ag+ toward Nitrosomonas europaea, a model ammonia-oxidizing bacterium. Nitrification inhibition of N. europaea by 1 ppm AgNPs and 0.5 ppm Ag+ was reduced from 80% and 83%, respectively, in the absence of Mg2+ to 2% and 33%, respectively, in the presence of 730 μM Mg2+. Introduction of Mg2+ resulted in the rapid aggregation of the AgNP suspensions and reduced the 3 h Ag+ dissolution rates from 30%, in the absence of Mg2+, to 9%, in the presence of 730 μM Mg2+. Reduced AgNP dissolution rates resulted in decreased concentrations of silver that were found adsorbed to N. europaea cells. Increasing AgNP concentrations in the presence of Mg2+ increased the observed inhibition of nitrification, but was always less than what was observed in the absence of Mg2+. The presence of Mg2+ also reduced the adsorption of Ag+ to cells, possibly due to multiple mechanisms, including a reduction in the negative surface charge of the N. europaea membrane and a competition between Mg2+ and Ag+ for membrane binding sites and transport into the cells. Ca2+ demonstrated similar protection mechanisms, as Ag+ toxicity was reduced and AgNP suspensions aggregated and decreased their dissolution rates. These results indicate that the toxicity of Ag+ and AgNPs to nitrifying bacteria in wastewater treatment would be less pronounced in systems with hard water.
Introduction
S
AgNPs in these applications are released either inadvertently (through the washing of the textiles) or intentionally (as used in sanitizing appliances) into commercial and residential wastewater systems and may bioaccumulate in wastewater treatment plant (WWTP) biosolids (Benn and Westerhoff, 2008; Blaser et al., 2008; USEPA, 2009; Benn et al., 2010). In addition, small but significant fractions of the AgNPs are expected to leave the WWTP through the effluent and enter into the receiving bodies of water (Kaegi et al., 2011). Therefore, AgNPs have the potential to interfere with critical biological functions, such as the nitrification process performed by ammonia-oxidizing bacteria (AOB), in both WWTPs and the natural environment (Choi and Hu, 2009; Giao et al., 2012).
Nitrosomonas europaea, a model AOB, has been used in studies investigating the toxicity of numerous pollutants on AOB, including AgNPs (Arp et al., 2002; Radniecki and Lauchnor, 2011; Radniecki et al., 2011). The relative toxicity of AgNPs toward nitrifying bacteria depended on the size and surface chemistries of the AgNPs, the presence of silver ligands, and the ionic strength of the aquatic environment (Marambio-Jones and Hoek, 2010; Radniecki et al., 2011; Arnaout and Gunsch, 2012; Kostigen Mumper et al., 2013; Yuan et al., 2013). All of these factors influence the rate of release of Ag+ from AgNPs, which has been shown to be a primary toxicity mechanism of AgNPs toward nitrifying bacteria (Radniecki et al., 2011; Arnaout and Gunsch, 2012).
Presence of divalent cations (e.g., Mg2+ or Ca2+) can induce AgNP aggregation, which can result in reduced AgNP dissolution and potentially limit the ability of AgNPs to adsorb onto microbial biomass (Jin et al., 2010; Zhang and Oyanedel-Craver, 2012). The concentrations of Mg2+ and Ca2+ can vary widely from 0.2–0.9 mM and 0.4–1.7 mM, respectively, for pristine surface waters to approximately 10 mM in groundwaters and wastewater impacted surface waters (Smith et al., 1987; Awadallah et al., 1996; DeSimone et al., 1997; van Langerak and Hamelers, 1997; Hammes et al., 2003). Thus, the presence of divalent cations may play a critical role in the expected fate, transport, and toxicity of AgNPs in both natural and engineered aquatic environments.
While recent studies have focused on the influence of divalent cations on AgNP stability and toxicity, few studies have examined how divalent cations influence the fate and toxicity of dissolved Ag+, a major source of AgNP toxicity. This study investigated the effects of Mg2+ and Ca2+ on Ag+ and AgNP toxicity toward N. europaea, including their influence on AgNP stability and dissolution rates. This work also demonstrates that Mg2+ and Ca2+ can lower Ag+ toxicity by preventing the adsorption of Ag+ by N. europaea cells. Thus, we propose that Mg2+ and Ca2+ can reduce AgNP toxicity through multiple mechanisms: (1) The destabilization of AgNPs resulting in a decrease in Ag+ dissolution rates and (2) The prevention of Ag+ from binding to N. europaea cells.
Materials and Methods
N. europaea culturing methods
N. europaea ATCC 19718 cells were grown in minimal growth medium in batch reactors as previously described (Radniecki and Lauchnor, 2011). After 3 d, cells reached mid-exponential growth (OD600 ∼0.070) and were harvested via centrifugation. The cells were washed in a 30 mM HEPES buffer (pH 7.8) to remove the presence of trace metals. The cells were suspended in a 30 mM HEPES buffer (pH 7.8) and were maintained at 4°C until they were used for experimentation.
AgNP and Ag+ inhibition studies
Previously developed methods were used to create dispersed AgNP suspensions in test media (Radniecki et al., 2011). In short, aliquots from 20 nm Citrate BioPure™ silver nanoparticles (nanoComposix, Inc., San Diego, CA; 1,000 ppm) or silver nitrate (AgNO3-10 ppm) stock solutions were placed into batch bioreactors containing deionized (DI) water and were dispersed via shaking. After 15 min of shaking, ammonium sulfate [(NH4)2SO4] and HEPES buffer (pH 7.8) were added to final concentrations of 2.5 mM and 30 mM, respectively. At this time, magnesium sulfate (MgSO4) was also added to Mg2+-containing batch bioreactors to a final concentration ranging from 0 to 730 μM, and calcium nitrate [Ca(NO3)2] was added to Ca2+-containing batch bioreactors to a final concentration ranging from 0 to 730 μM. The batch bioreactors were shaken for an additional 30 min before N. europaea cells were added to a final concentration of 5 mg protein L−1 (OD600≈0.072). All conditions were run in triplicate batch bioreactors.
The batch bioreactors were shaken in the dark at 250 rpm for 3-h at 30°C. To monitor N. europaea nitrification activity, nitrite production was measured at 45 min intervals throughout the 3-h experiment, via colorimetric assay, as previously described (Hageman and Hucklesby, 1971; Radniecki et al., 2011). The nitrite production rates in both treatment and control bioreactors were normalized to their respective cell densities. The nitrite production rates for all control conditions tested were 1.61±0.07 μmol NO2−/mg protein*min. Percent nitrification activity refers to the nitrite production rates of bioreactors containing AgNP or Ag+ compared with the nitrite production rates of control bioreactors not containing AgNP or Ag+.
Abiotic AgNPs stability studies
The aggregation state of 20 nm citrate AgNP suspensions, created as described earlier, in the presence and absence of either Mg2+ or Ca2+ was quantified using time-resolved dynamic light scattering (TR-DLS) by measuring the z-average diameter at 15 s intervals for 60 min with a 90Plus particle size analyzer (Brookhaven Instruments, Holtsville, NY). Due to the interference of their large size, N. europaea cells were not present. To minimize background noise, the DDI water and HEPES buffer (pH 7.8)/(NH4)2SO4 solution were filtered through a 20 nm syringe filter (Whatman International Ltd., Maidstone, England) to remove small ambient particles before the AgNP were added to a final concentration of 1 ppm.
Abiotic AgNPs dissolution studies
The dissolution of 20 nm citrate AgNP suspensions, created as described earlier, in the presence and absence of either Mg2+ or Ca2+ was quantified using ultra-filtration and inductively coupled plasma optical emission spectroscopy (ICP-OES) as previously described (Radniecki et al., 2011). In short, abiotic batch reactors were shaken at 250 rpm in the dark at 30°C for 3 h, at which time the total silver [both dissolved silver (Ag+) and AgNP] concentrations were measured via ICP-OES using standard methods (APHA, 1998). After 3 h, 15 mL of the test solution was filtered through a centrifugal ultra-filtration device with a 5 kDa molecular-weight cutoff (Amicon Ultra-15; Millipore Co., Billericka, MA) for 30 min at 5,000 g per manufacturer's instruction. The Ag+ content of the filtrate was measured via ICP-OES and may include the sum of numerous silver chemical species [e.g., Ag+, AgSO4, AgNH3+, or Ag(NH3)2+]. All conditions were tested in triplicate abiotic batch reactors.
To determine the amount of Ag+ that may be retained by the filter, 0.5 to 2 mM AgNO3 solutions were filtered using centrifugal ultra-filtration as described earlier. On average, 10%±0.6% of the AgNO3 was retained by the filter for all concentrations tested.
Ag+ and AgNP adsorption studies
The adsorption of either Ag+ or 20 nm citrate AgNP suspensions to N. europaea cells was measured on completion of the 3 h inhibition assays described earlier. Cells were harvested from the batch bioreactors via centrifugation for 20 min at 9,000 rpm (8,324 g). The resulting cell pellet was suspended in 500 μL of DI water and digested overnight at 60°C in 3 mL of an 11.2 N nitric acid and 12% phosphoric acid solution. The digested pellets were diluted twofold in DI water before Ag analysis was conducted via ICP-OES, as described above.
To determine the effect of centrifugation on the AgNP suspensions, abiotic control batch reactors were run for 3 h, as described above. The abiotic control batch reactors contained 2 ppm AgNPs, 30 mM HEPES buffer (pH 7.8), and an absence of both N. europaea cells and (NH4)2SO4. (NH4)2SO4 is a known accelerator of AgNP dissolution and was removed to limit AgNP dissolution to nondetectable rates over the course of a 3-h experiment (Kostigen Mumper et al., 2013). The contents of the abiotic control batch reactors were placed into a centrifuge tube and centrifuged at 9,000 rpm (8,324 g) for 20 min, after which the supernatant was placed into a fresh centrifuge tube.
One milliliter of HEPES buffer (pH 7.8) was added to the centrifuge tube and vigorously shaken and pipetted to dislodge any pellet that may have formed during the centrifugation processes. This is the same procedure that was applied to dislodge the N. europaea cell pellet which formed during the biotic tests before placing the cell pellet in the acid digestion mixture. The 1 mL of HEPES buffer (pH 7.8) was then placed into a fresh centrifuge tube.
The initial AgNP concentration of the abiotic control batch reactors along with the AgNP concentration of the supernatant and “cell pellet” was measured via scanning UV-vis spectroscopy from 300 to 1,100 nm, as previously described (Kostigen Mumper et al., 2013; Ostermeyer et al., 2013).
Results
Effect of centrifugation on the AgNP suspensions
Results of the abiotic control batch reactors indicated that 42%±5% of the AgNPs pelleted out of solution during the centrifugation process. However, the vast majority of the AgNPs that pelleted out of solution remained immobilized on the plastic centrifuge tube, as only 7%±4% of the AgNPs were transferred from the “cell pellet” into a fresh centrifuge tube (Supplementary Fig. S1). The immobilization of the AgNPs to the plastic centrifuge tube is further confirmed by the poor mass balance observed by the AgNP biological tests compared with the excellent mass balance observed during the Ag+ biological tests (Supplementary Fig. S2).
Given the high density of AgNPs compared with the density of bacterial cells, our measurements indicated that the AgNPs fell out of solution first, followed by N. europaea cells with adsorbed Ag+/AgNPs followed by N. europaea cells not containing adsorbed Ag+/AgNPs. Therefore, during centrifugation, about 40% of the AgNPs not attached to N. europaea cells pelleted out of solution first, became immobilized against the plastic centrifuge tube, and were not transferred over with the N. europaea cell pellets into the acid digestion vessel of the total silver assay. Thus, our measurements of the Ag+/AgNPs associated with cells were not significantly biased by the precipitation of nonadsorbed AgNPs.
Mg2+-induced reduction of AgNP toxicity
N. europaea cells exposed to 1 ppm 20 nm citrate AgNPs showed a nonlinear increase in nitrification activity with increasing Mg2+ concentrations (Fig. 1). At 0 μM Mg2+, exposure to 1 ppm AgNPs reduced nitrification activity to 20%±9%; while the addition of 730 μM Mg2+ increased nitrification activity to 98%±5% of the nonexposed control cells. When 730 μM Mg2+ was present in the abiotic reactors, the AgNPs rapidly aggregated and the 3-h dissolution rate decreased from 30% to 9% (Fig. 2). This reduction in AgNP dissolution can help explain how Mg2+ protects N. europaea cells from AgNP toxicity. However, the protection provided by 730 μM Mg2+ was overcome by increasing the AgNP concentrations to 2 and 4 ppm, which resulted in a reduction of nitrification activities to 30%±5% and 14%±2%, respectively (Fig. 3A).
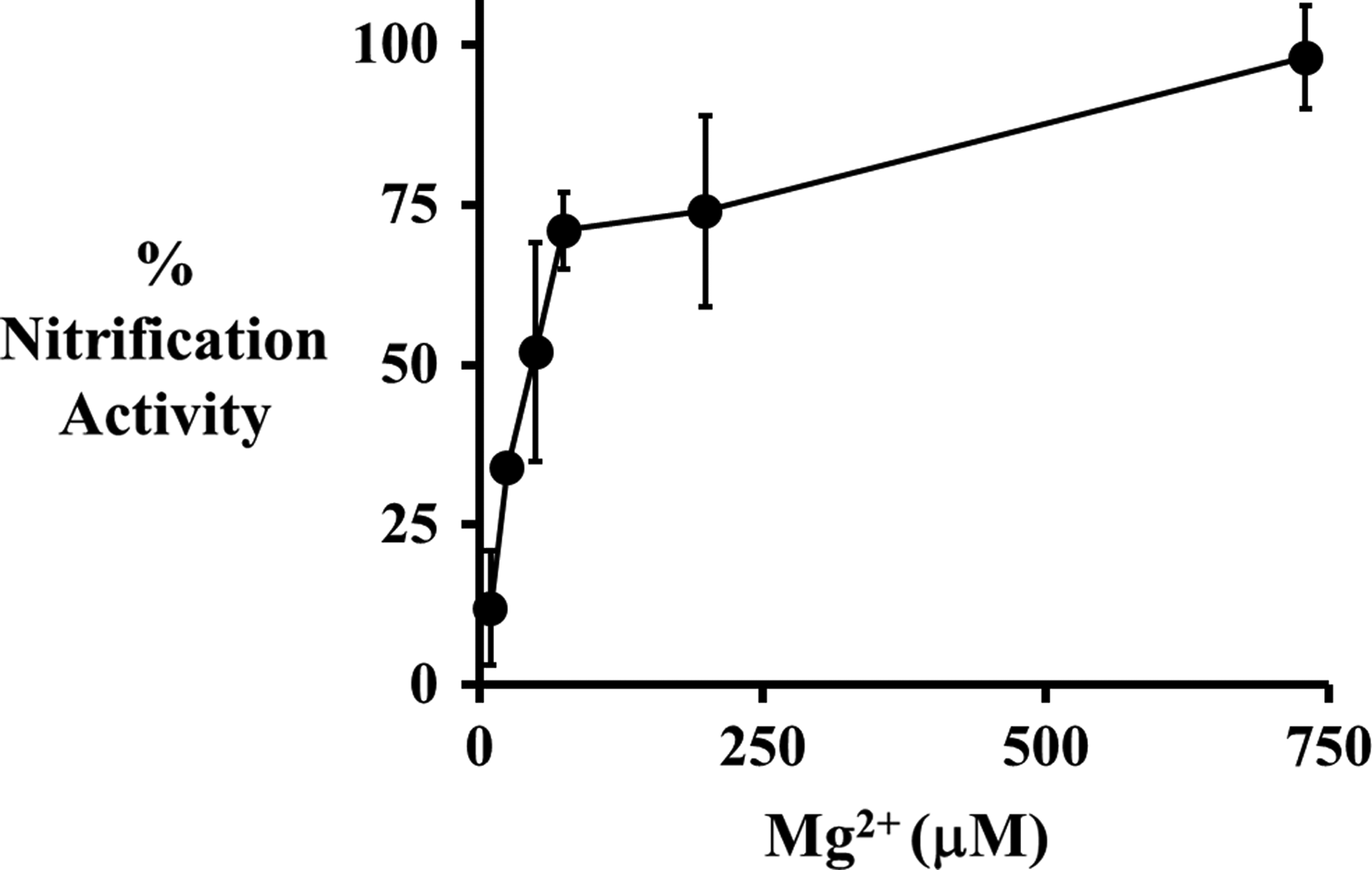
Percent nitrification activity of N. europaea after 3 h exposure to test media containing 1 ppm 20 nm citrate AgNPs (●) concentrations of Mg2+ ranging from 0 to 730 μM (0–17.7 ppm). Error bars represent 95% confidence intervals.
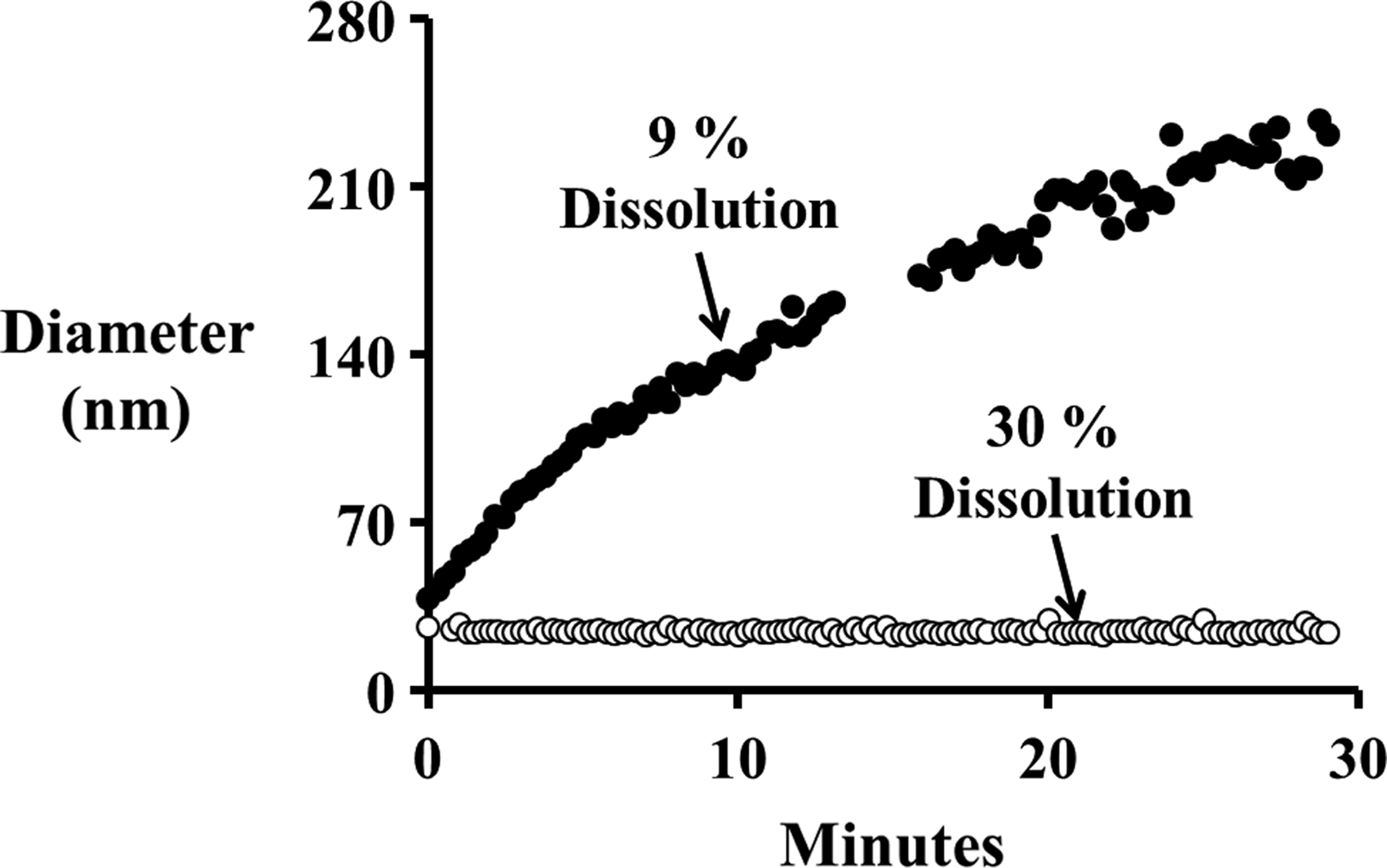
Average diameter of 20 nm AgNPs in test media containing 0 μM Mg2+ (○) and 730 μM (17.7 ppm) Mg2+ (●). Percent dissolution rates were calculated after 3 h of exposure to the test media.
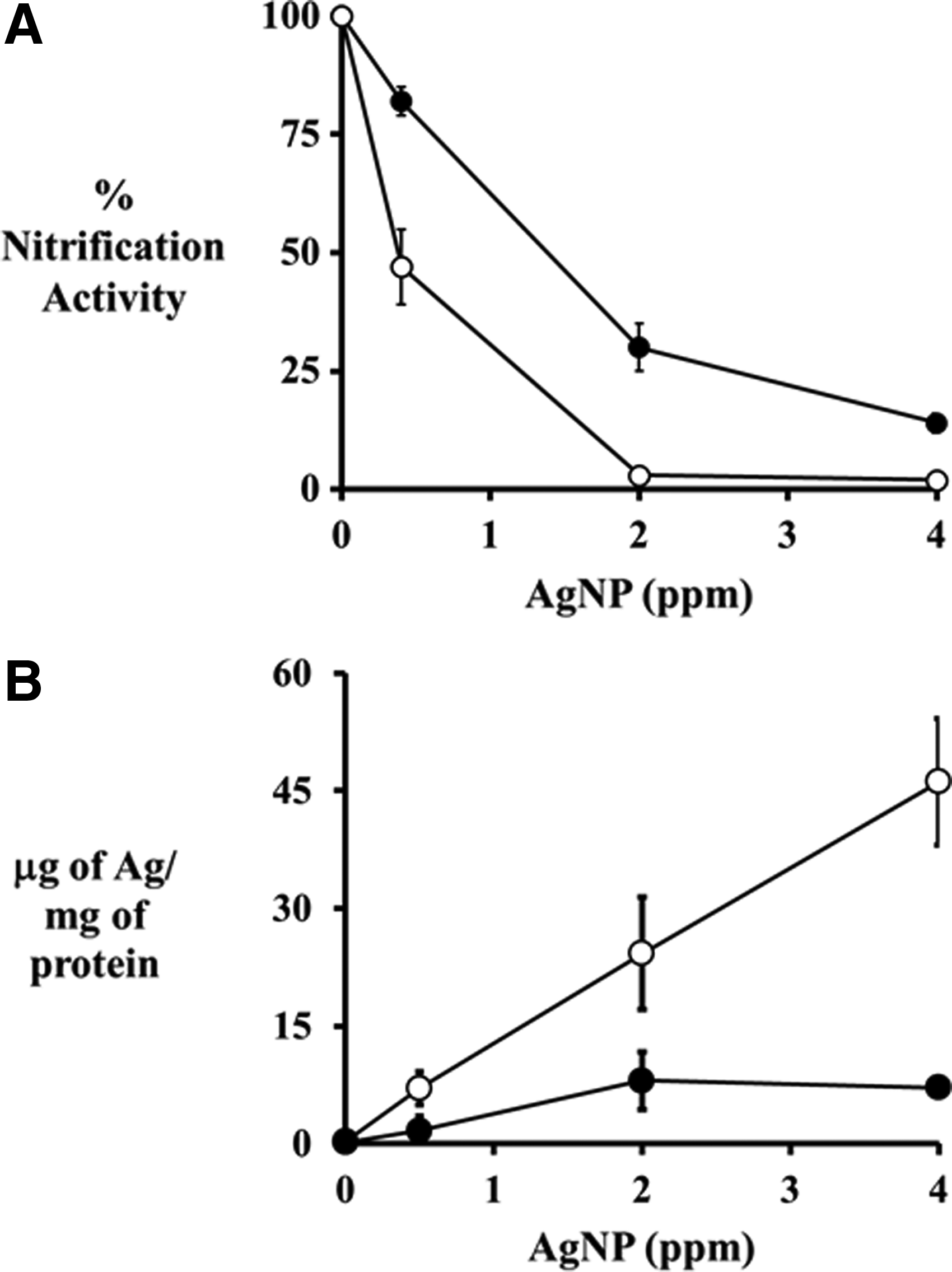
Percent nitrification activity of N. europaea cells (
When Mg2+ was absent from the test medium, N. europaea cells accumulated higher total silver concentrations as the AgNP concentrations increased (Fig. 3B). When exposed to 0.5 ppm AgNPs, N. europaea cells accumulated 7±2 μg of Ag/mg of protein; while when the AgNP concentration was increased to 4 ppm, 46±8 μg of Ag/mg of protein accumulated. Interestingly, when 730 μM Mg2+ was present in the test medium, N. europaea cells did not accumulate additional total silver when AgNP concentrations increased from 2 to 4 ppm. Rather, the cell-associated silver concentrations were 8±3 and 7±0.5 mg of Ag/mg of protein, respectively (Fig. 3B). However, the overall trend showed that nitrification activity tracked strongly with the amount of total silver associated with the cells for all AgNP concentrations and conditions tested (Fig. 3).
Mg2+-induced reduction of Ag+ toxicity
N. europaea cells exposed to 0.5 ppm Ag+ showed a nonlinear increase in nitrification activity with increasing Mg2+ concentrations (Fig. 4A). At 0 μM Mg2+, exposure to 0.5 ppm Ag+ reduced nitrification activity to 17%; while the addition of 730 μM Mg2+ increased nitrification activity to 67%. The increase in nitrification activity correlated with a decrease in total silver accumulated on the cells (Fig. 4B). At 0 μM Mg2+, N. europaea cells accumulated silver to a concentration of 9±1 μg of Ag/mg of protein, while the addition of 730 μM Mg2+ decreased the accumulation of silver to 1.5±1.6 μg of Ag/mg of protein.
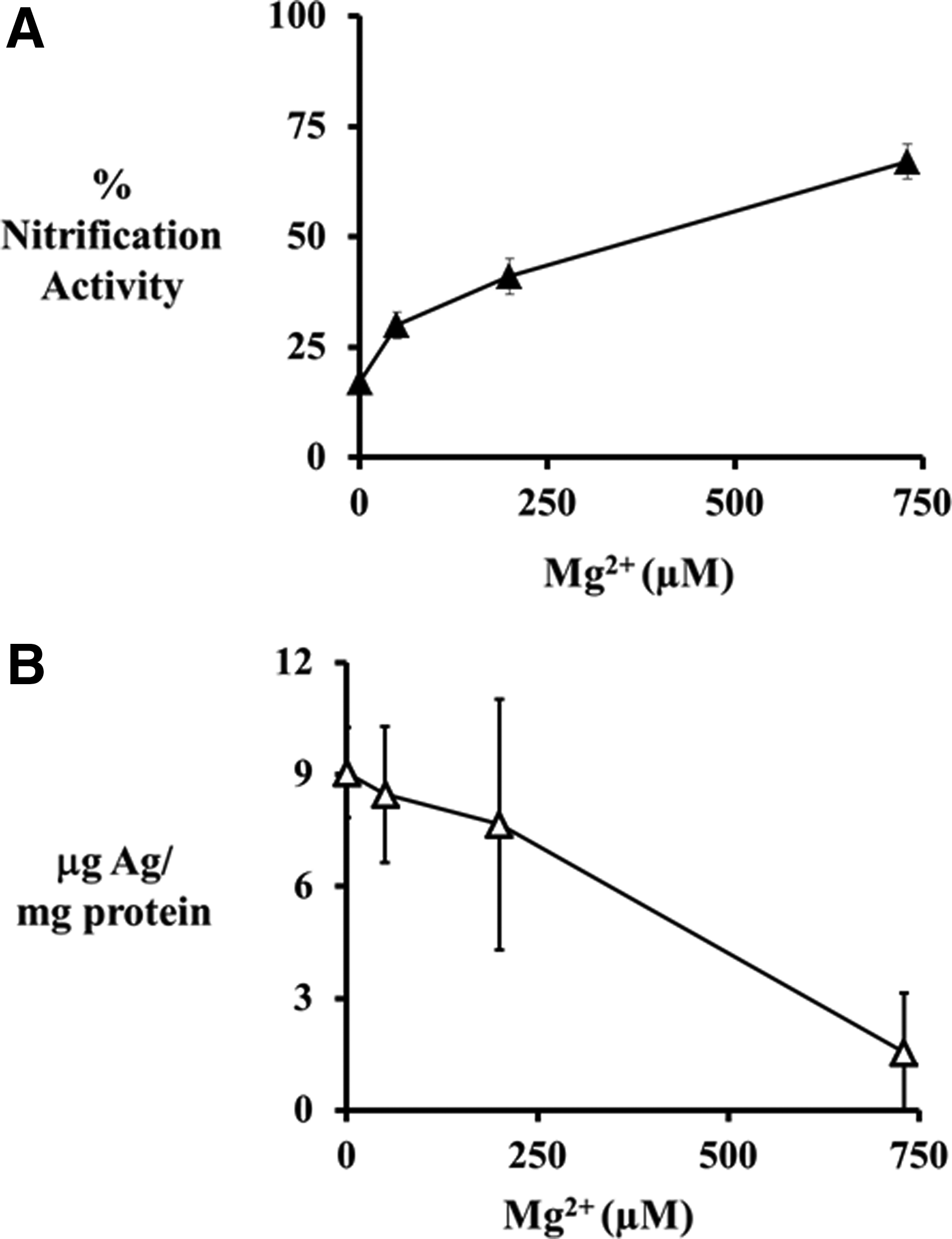
Percent nitrification activity (▴) of N. europaea cells (
Similar to AgNPs, increasing the Ag+ concentration resulted in a decrease in nitrification activity in the presence and absence of 730 μM Mg2+ (Fig. 5A). In cell exposures to Ag+ in medium containing 730 μM Mg2+, the nitrification activity was consistently 20–22% higher compared with cell exposures to Ag+ in medium not containing Mg2+ across all Ag+ concentrations tested. The increased protection is due to a decrease in Ag+ adsorbed in the N. europaea cells, which was 67–78% lower in exposures containing Mg2+ compared with those lacking the Mg2+ (Fig. 5B). It is interesting to note the similarities in the results obtained with the AgNP exposures and the Ag+ exposures, indicating similar inhibition processes.
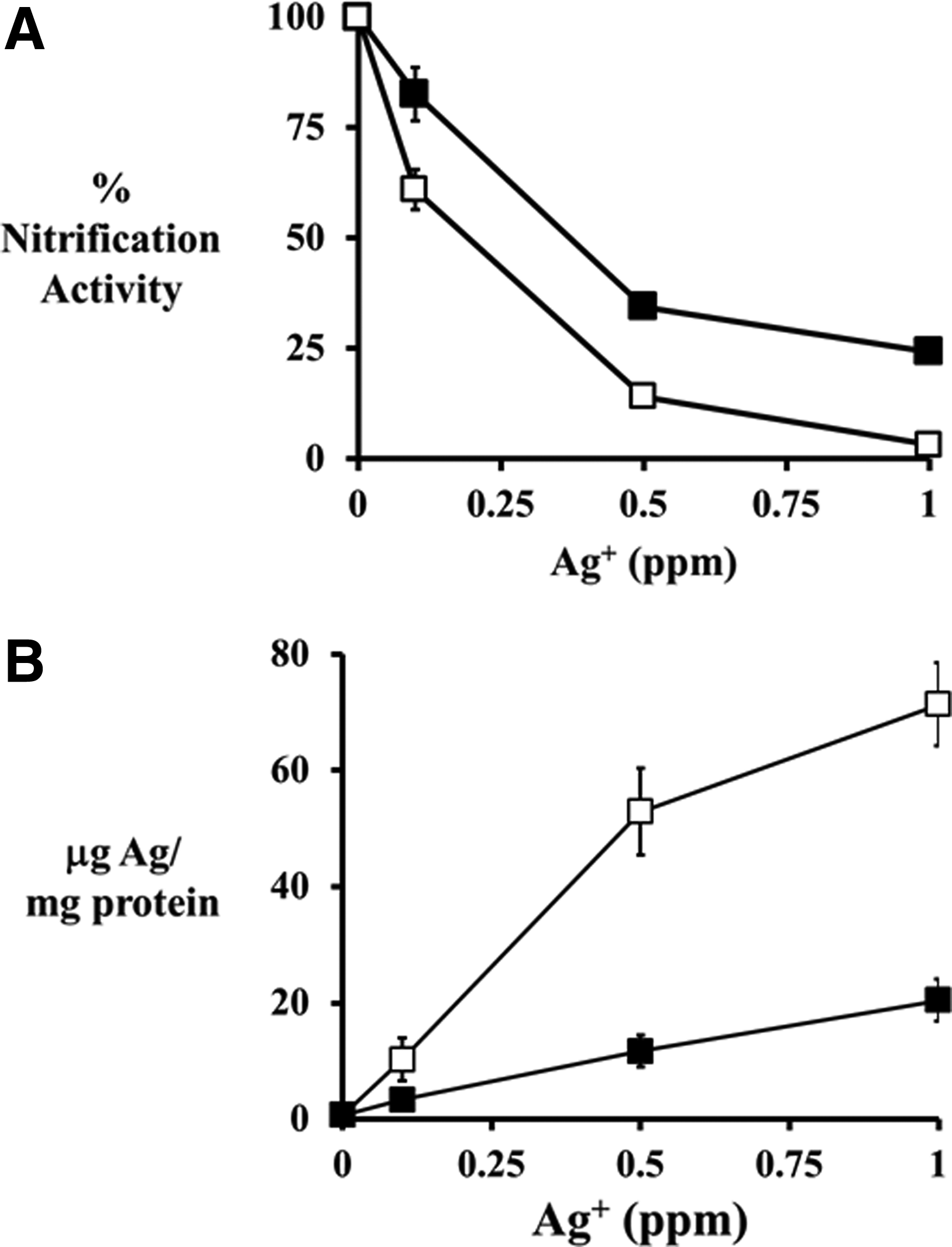
Percent nitrification activity of N. europaea cells (
Ca2+-induced reduction of Ag+ and AgNP toxicity
In the presence of 200 μM Ca2+, 20 nm citrate AgNPs aggregated slightly and the 3-h dissolution rate decreased from 30%±1.6% to 19%±2% (Fig. 6A). This decrease is dissolution corresponded with a decrease in toxicity with nitrification activity rising from 20%±9% to 82%±11% (Fig. 6B).
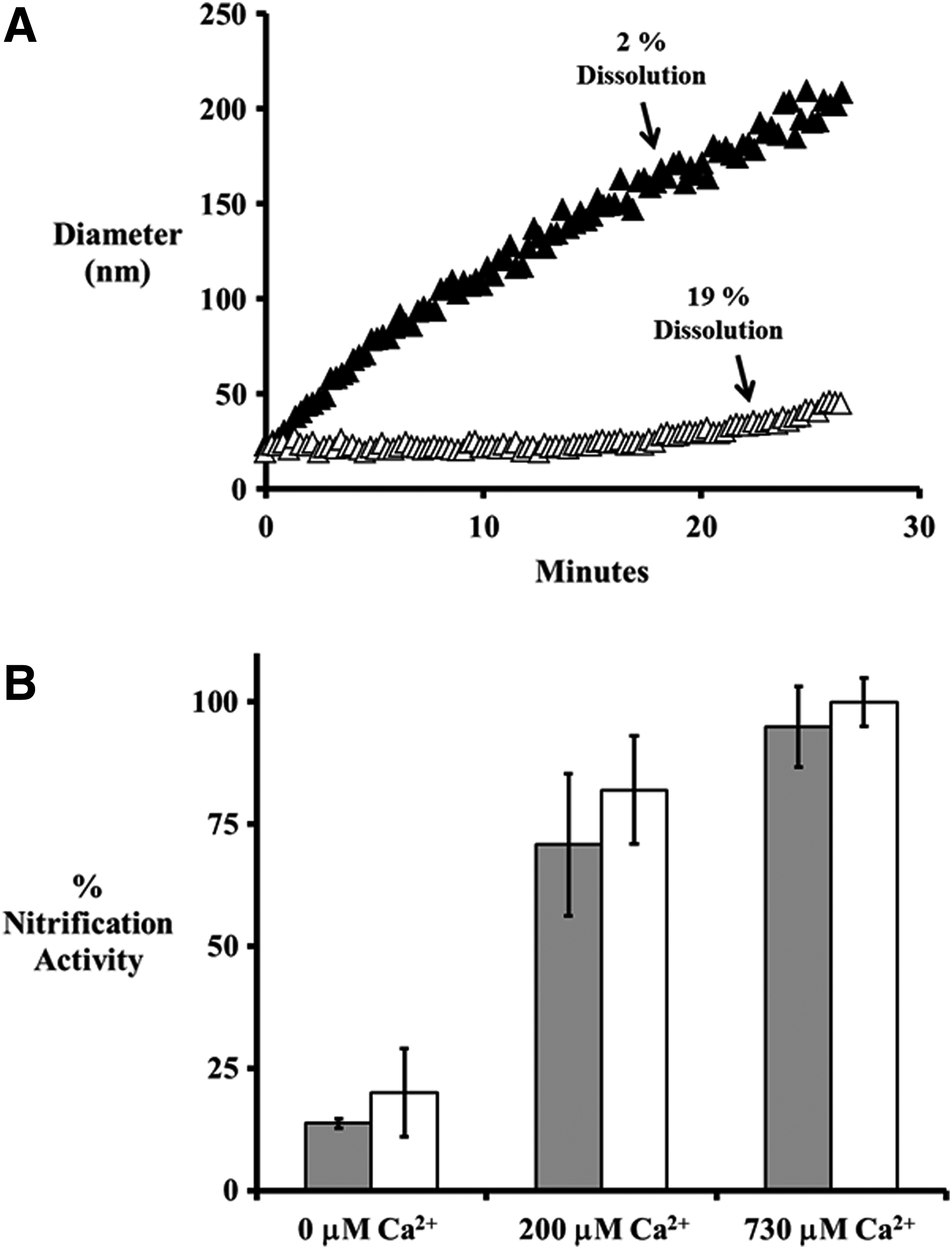
(
AgNPs exposed to 730 μM Ca2+ rapidly aggregated and the 3-h dissolution rate drastically decreased from 30%±1.6% to 2%±1% (Fig. 6A). This extreme decrease in AgNP dissolution corresponded with a complete absence of toxicity with nitrification rates measured at 100%±5% (Fig. 6B).
Similar to Mg2+, increased Ca2+ concentrations resulted in increased nitrification rates in N. europaea cells exposed to Ag+. The nitrification activity increased from 13.8%±1% when exposed to 0.1 ppm Ag+, to 70.8%±14.6% when 200 μM Ca2+ was present, and to 94.9%±8.3% when 730 μM Ca2+ was present (Fig. 6B).
Discussion
This work demonstrates that Mg2+ and Ca2+ have a protective effect on N. europaea cells exposed to AgNPs with increasing concentrations of these divalent cations, resulting in increased protection (Figs. 1, 3A and 6B). Similar trends have been observed in other gram-negative bacteria exposed to AgNPs and varying concentrations of Mg2+ and Ca2+ (Marambio-Jones and Hoek, 2010; Zhang and Oyanedel-Craver, 2012; Zhang et al., 2012).
One possible mechanism for the observed protection from AgNP toxicity is the Mg2+- or Ca2+-induced aggregation of the AgNPs, which resulted in significant decreases in the dissolution of Ag+ (Figs. 2 and 6A) and a decrease in silver associated with the N. europaea cells (Fig. 3B). Several studies have previously shown that Mg2+ can neutralize the stabilizing electrostatic double layer of AgNPs, which results in aggregation of the AgNPs. The aggregation leads to a reduction in AgNP dissolution rates due to a smaller surface area-to-volume ratio (Jin et al., 2010; Marambio-Jones and Hoek, 2010; Huynh and Chen, 2011; Zhang and Oyanedel-Craver, 2012; Zhang et al., 2012). This reduction in dissolution rates is critical, as the dissolution of Ag+ has been shown to be responsible for the majority of the inhibition observed in N. europaea cells exposed to AgNPs (Radniecki et al., 2011; Arnaout and Gunsch, 2012; Kostigen Mumper et al., 2013).
However, even in a highly aggregated state, AgNPs can still be toxic at high concentrations (≥2 ppm), though less toxic than their nonaggregated counterparts at the same concentration (Fig. 3A). It should be noted that AgNP concentrations ≥2 ppm are not likely to occur in most natural environments outside of an accidental spill site.
Similar to Mg2+, previous research has demonstrated that Ca2+ can induce aggregation of AgNPs through a reduction in the electrostatic double layer, and this aggregation results in a decrease in Ag+ dissolution as the surface area-to-volume ratio decreases (Huynh and Chen, 2011; Zhang and Oyanedel-Craver, 2012). This was observed in AgNP suspensions exposed to 730 μM Ca2+ as they rapidly aggregated, decreasing their dissolution rates by 93% compared with nonaggregated AgNPs (Fig. 6A). This decrease in dissolution eliminated their observed toxicity toward N. europaea (Fig. 6B).
Toxicity of the AgNPs was also greatly reduced, by 77% (from 80% inhibition to 18% inhibition), when 200 μM Ca2+ was present (Fig. 6B). However, dissolution rates only decreased by 37% (from 30% to 19%) on exposure to 200 μM Ca2+ (Fig. 6A). Thus, the observed protection from AgNP-induced inhibition afforded by 200 μM Ca2+ cannot be solely explained by a decrease in AgNP dissolution rates.
To investigate what may be causing the observed decrease in toxicity of the AgNPs in the presence of 200 μM Ca2+, the protective potential of Ca2+ against Ag+ toxicity was explored. Similar to the AgNP experiments, increasing concentrations of Ca2+ resulted in increased protection from Ag+ toxicity (Fig. 6B). Since the Ca2+ was added as Ca(NO3)2, equilibrium models do not predict a difference in silver speciation between media containing Ag+ without Ca(NO3)2 compared with media containing Ag+ with Ca(NO3)2 (Supplementary Table S1). Thus, it is unlikely that the protection observed is related to potential changes in silver speciation.
Further studies were conducted to determine whether Mg2+ has a similar protective effect against Ag+ toxicity. Surprisingly, results from these studies demonstrated that the presence of Mg2+ can greatly reduce Ag+ toxicity toward N. europaea with higher Mg2+ concentrations, resulting in lower toxicity levels (Fig. 4A). The Mg2+ were added to the system, as MgSO4 and equilibrium models do not predict a difference in silver speciation between media containing Ag+ with MgSO4 and media containing Ag+ without MgSO4 (Supplementary Table S1). Therefore, similar to the results with Ca2+ protection, the observed protection against Ag+ toxicity is not likely to be due to changes in silver speciation.
However, the protection mechanism of Mg2+ against Ag+ toxicity becomes more evident when the amount of silver adsorbed to the N. europaea cells is measured. Increased Mg2+ concentrations led to decreased accumulated silver adsorbed to the N. europaea cells with the addition of 730 μM Mg2+ resulting in an 83% reduction in absorbed silver compared with N. europaea cells exposed to Ag+ without Mg2+ present (Fig. 4B). It can, therefore, be concluded that this lack of silver accumulation is responsible for the observed protection of Mg2+ against Ag+ toxicity. These results are consistent with our original observation that AgNP toxicity toward N. europaea is correlated with the silver adsorbed to the cells (Radniecki et al., 2011).
One possible explanation of the decrease of Ag+ accumulation in the presence of Mg2+ is the reduction of the bacterial membranes' negative surface charge. Gram-negative bacteria have a negative net surface charge due to the negatively charged functional groups of the lipopolysaccharide (LPS) layer (Costerton et al., 1974). Mg2+ acts to stabilize bacterial membranes through the bridging of the negative charges of LPS functional groups, including phosphates, phosphatidic acid, and 2-keto-3-deozyoctulosonic acid (Schindler and Osborn, 1979; Rivera et al., 1988; Amro et al., 2000). Mg2+ will also bind to other membrane constituents, including inorganic phosphates and teichoic acids (Heptinstall et al., 1970; Naumann et al., 1989; van Veen, 1997). Thus, the presence of Mg2+ has been shown to reduce the net surface charge of gram-negative bacteria (De Kerchove and Elimelech, 2008; Zhang and Oyanedel-Craver, 2012; Zhang et al., 2012). This lower surface charge may reduce Ag+ interactions with N. europaea by decreasing the electrostatic attraction between the positively charged Ag+ and the negatively charged N. europaea.
In addition to reducing attractive electrostatic forces, excess Mg2+ may reduce Ag+ ability to bind to N. europaea cells by competing with Ag+ for these binding spots on these membrane constituents. Excess Mg2+ may also reduce Ag+ ability to enter into the N. europaea cells through Mg2+ transporting proteins, as Ag+ have been shown to be capable of entering into cells through Mg2+ transporter proteins (Nies, 1999). Previous studies have demonstrated that increased Mg2+ will out compete other heavy metals, including Cd2+ and Zn2+, for entrance into N. europaea cells through these transporters (Radniecki et al., 2009a, 2009b). Thus, higher concentrations of Mg2+ may reduce Ag+ transport through these proteins.
Conclusion
Mg2+ and Ca2+ are the primary constituents of water hardness and may play a critical role in the fate, transport, and toxicity of AgNPs in both engineered and natural water systems. While it has been previously established that divalent cations can decrease AgNP toxicity by inducing aggregation and decreasing the rate of Ag+ release, the effect of divalent cations on the toxicity of the released Ag+ has not been as thoroughly characterized.
The work presented here is among the first to demonstrate that Mg2+ and Ca2+ utilize multiple mechanisms to protect N. europaea from AgNPs and Ag+. At relatively high concentrations (e.g., 730 μM), both Mg2+ and Ca2+ induced rapid aggregation of the AgNP suspensions, which led to highly reduced dissolution rates, which accounted for the vast majority of the protection observed. However, at lower Ca2+ concentrations (e.g., 200 μM), the AgNP suspensions only slightly aggregated, which led to a moderate decrease in dissolution rates and could not account for the high level of protection observed. This suggested that the Ca2+ are not only decreasing the dissolution rates of the AgNPs but also reducing the toxicity of the dissolved Ag+ which are released.
Both Ca2+ and Mg2+ were shown to be able to protect N. europaea from Ag+ toxicity. Mg2+, specifically, was shown to reduce Ag+ adsorption to the N. europaea cells. This reduction in silver accumulation could account for the observed decrease Ag+ toxicity. Therefore, these studies suggest that Mg2+ and Ca2+ can have an important impact on the ultimate fate and toxicity of the dissolved Ag+ from AgNPs and should be carefully considered during AgNP toxicity testing.
Based on the results of this study, it can be expected that in hard water environments, the toxicity of AgNPs and dissolved Ag+ would be significantly muted compared with soft water environments. In addition, in hard water environments lacking stabilizing silver ligands (e.g., natural organic matter), AgNPs would be expected to aggregate and begin to fall out of solution. This may have important implications in determining the fate and toxicity of AgNPs in WWTPs, where AgNPs may precipitate out of solution in primary clarifiers, or attach to microbial biomasses in the secondary clarifier, thus limiting their release from the WWTP through the effluent.
Footnotes
Acknowledgments
The authors would like to thank the Dan Arp lab at Oregon State University for kindly supplying the N. europaea culture. They would like to thank Jeff Nason and Dylan Stankus for conducting the DLS measurements used in this study. They would also like to thank J.R. Giska and Margaret Schneider for help with inhibition experiments and ICP analysis. Funding was provided by a grant from the National Science Foundation's Division of Chemical, Bioengineering, Environmental, and Transport Systems–Environmental Health and Safety of Nanotechnology program (# 1067572). Joseph Anderson was also funded through the Pete and Rosalie Johnson Internship Program at Oregon State University.
Author Disclosure Statement
No competing financial interests exist for the authors of this work.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.