
Abstract
Abstract
Biodiesel has received much attention in recent years. Due to significant lipid and carbohydrate production and the ability to grow using nonpotable water sources, microalgae are being scrutinized as a potential high-yield feedstock for biofuel production. For biodiesel production, selection of suitable strain of microalgae depends on various factors, such as lipid productivity, percentage of lipids, biomass productivity, CO2 tolerance, and nutrient requirements. In both natural and engineered systems, microalgae can be exposed to a variety of environmental conditions that have an effect on cellular composition and growth rate. Furthermore, the amount of carbon fixed in lipids and carbohydrates is highly influenced by nutrient availability and environmental factors. This article reviews the potential of microalgae for lipid production and effects of environmental factors (e.g., light, temperature, and pH) and nutrient availability (e.g., nitrogen, carbon, phosphorus, and trace metals).
Introduction
O
In the production of microalgal biodiesel, the main problems are selection of suitable microalgae for biodiesel production and various environmental factors that affect lipid content and biomass of microalgae. This article reviews the potential of using different types of microalgal species as source of oil and the effects of various environmental factors and nutrient availability on microalgae for biodiesel production.
Biodiesel
Howell (1997) defined biodiesel as “The mono alkyl esters of long fatty acids derived from renewable lipid feedstock such as vegetable oils or animal fats, for use in compression ignition (diesel) engines” as given by the National Biodiesel Board, 1996. In a simple way, biodiesel is a renewable fuel produced from methyl alcohol and animal fats, vegetable oils, and recycled cooking fats (Tyson and McCormick, 2006). Biodiesel properties are very close to those of petroleum diesel.
The most widespread technique to produce biodiesel is through transesterification, a process that involves changing the chemical properties of the oil by using methyl alcohol (Ma and Hanna, 1999), as shown in Fig. 1 (Rajvanshi and Sharma, 2012). The reaction occurs in a stepwise manner: triacylglycerols (TAG) are first converted to diacylglycerols (DAG), subsequently to monoacylglycerols (MAG), and finally to glycerol (Gerpen, 2005).
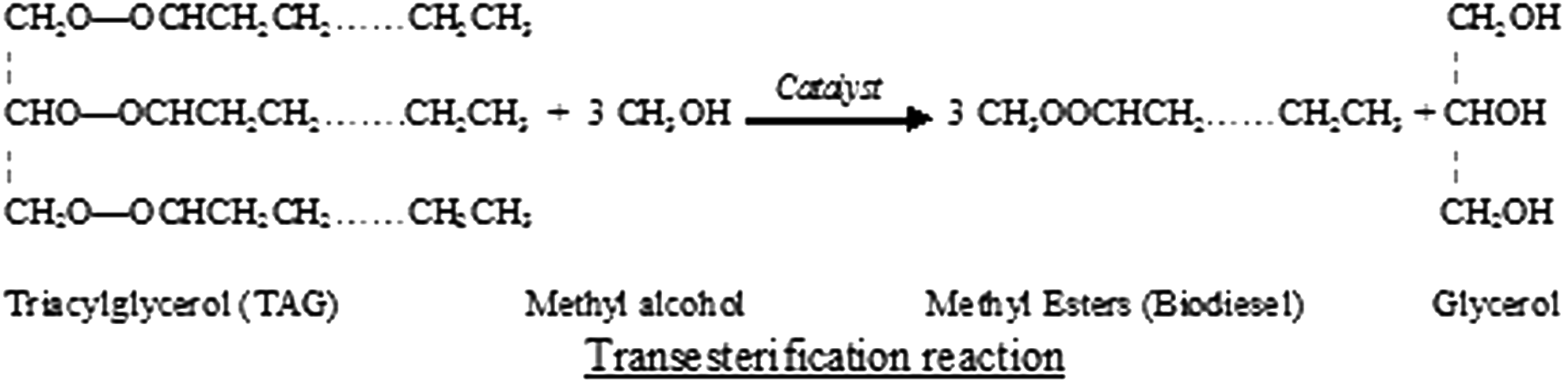
Transesterification reaction, which requires 3 mol of alcohol for each mole of triacylglycerol to produce 1 mol of glycerol and 3 mol of methyl esters.
The first-generation biofuels refer to the fuels that have been derived from sources, such as starch, sugar, vegetable oils, and animal fats. These production systems have considerable economic and environmental limitations. The most general concern related to the current first-generation biofuels is that as production capacities raise, so does their competition with agriculture for land used for food production. The second-generation biofuels are proposed to produce fuels from lignocellulosic biomass that do not compete with food production. Sources consist of agricultural residues and nonedible components of corn or sugarcane. Finally, the third-generation biofuels that are produced from microalgae are considered to be a doable alternative energy source that is devoid of the main problems allied with the first- and second-generation biofuels. On an area basis, microalgae are competent to produce 15–300 times more oil than traditional crops for biodiesel production, although due to high water content harvesting is major (Pragya et al., 2013).
Benefits of biodiesel
Biodiesel is more environmental friendly than petroleum diesel and likewise regarded as “carbon neutral” because essentially all the carbon dioxide (CO2) discharged during consumption had been sequestered from the atmosphere for the growth of oil crops (Barnwal and Sharma, 2005). Biodiesel is renewable, biodegradable, nontoxic, and has low emission profiles compared with petroleum diesel. The biggest advantages of biodiesel compared with many other alternative transportation fuels are that it can be utilized in existing diesel engines with no alteration and may be blended with petroleum diesel in mandatory ratio. Biodiesel performs as well as petroleum diesel, whereas at the same time, it reduces emissions of particulate matter, carbon monoxide (CO), hydrocarbons, and oxides of sulfur (SOx) (Barnwal and Sharma, 2005). Emissions of oxides of nitrogen (NOx) are, however, higher for biodiesel in several engines. Additional environmental benefits of biodiesel include the fact that it reduces emissions of air-borne toxins and carcinogens (comparative to petroleum diesel). Practice of biodiesel will permit a balance to be sought between economic, agriculture development, and the environment.
Microalgae
The word “microalgae” is used to express a large type of prokaryotic (strictly termed Cyanobacteria) and eukaryotic organisms, with a variety of phylogenies and morphologies. They can survive in different environments, such as salt and fresh water. They vary in size, shape, and color (Barsanti and Gualtieri, 2005). Algae have been investigated as a source of energy in various diverse contexts, ranging from direct combustion to the production of hydrogen gas (Golueke et al., 1957; Demirbas, 2011). In contrast to land-based oil crops, where just the seeds are harvested, each algal cell contains lipid, and thus, the yield of product from biomass is far higher (Becker, 1994). Research has focused on microalgae for the mass production of oil owing to their high lipid content compared with macroalgae. Microalgae have higher rates of growth than other feedstocks. As a result of these differences, the yield of oil per area of microalgal cultures potentially exceeds that of the superlative oilseed crops. Comparison of microalgae with other biodiesel feedstocks is given in Table 1.
High oil-containing algal species are particularly attractive as a feedstock for biodiesel production. Because of their simple cellular structure and endurance in an aqueous environment, the intact cell is out there for capturing light by pigments and mass transfer, resulting in relatively high substrate uptake rate and photosynthetic potency (Sheehan et al., 1998; Miao and Wu, 2006).
Most algal species considered for biodiesel production are either green algae (Chlorophyta) or diatoms (Bacillariophyta) (Sheehan et al., 1998). They are generally photosynthetic; however, several species are able to grow heterotrophically or mixotrophically (Barsanti and Gualtieri, 2005; Chen et al., 2011). The mass production of oil is focused mainly on microalgae of 0.4 mm diameter of diatoms and cyanobacteria rather than macroalgae (Horn, 2009). Botryococcus braunii, Navicula pelliculosa, Neochloris oleoabundans, Amphora, Nannochloropsis oculata, and Spirulina maxima are some of the microalgal species currently studied for their suitability for biodiesel production (Li et al., 2008a; Chiu et al., 2009; Gouveia and Oliveira, 2009; Griffiths and Harrison, 2009; Nascimento et al., 2013).
Microalgae lipid
The most important components of algal cells are lipids and carbohydrates for biofuels (Becker, 1994). Microalgae in nature produce lipids as part of the structure of the cell and as a storage compound (Tsukahara and Sawayama, 2005). Storage lipids, TAG, DAG, and MAG forms, accumulate in lipid vesicles called oil bodies in the cytoplasm. Under certain conditions, generally triggered by stress or the cessation of growth, lipid content can increase to over 60% of cell dry weight (DW), mostly composed of TAGs (Becker, 1994).
TAG is the most suitable class of lipids for biodiesel production. Phospholipids are particularly undesirable as they increase consumption of catalyst and act as emulsifiers, impeding phase separation during transesterification (Gerpen, 2005). Phospholipids and several sulfur-containing glycolipids also increase the phosphorous and sulfur content of the fuel, respectively, which must both be below 10 mg/L to meet the European biodiesel standard EN 14214. For biodiesel production, it is therefore essential to maximize not only total lipid production but also TAG content and appropriate fatty acid profile.
Depending on species, microalgae produce many different kinds of lipids, hydrocarbons, and other complex oils. Heterotrophically cultivated Chlorella protothecoides has accumulated higher lipids (about 55% of DW) compared with photoautotrophically (14% of DW) grown cells (Miao and Wu, 2006). A further natural mechanism through which microalgae can alter lipid metabolism is the stress response owing to a lack of nitrogen availability.
Lipid production varies within a vast range among the algal strains (4% to 80% DW basis), and the variation is affected by the environmental conditions. For example, in the case of B. braunii Kützing (Smittenberg et al., 2005), the yield of oil per unit area is estimated to be from 5,000 to 20,000 gallons/acre/year, which is 7–31 times greater than the next best crop, palm oil (635 gallons/acre/year) (Raja et al., 2008). The suitable microalgae strain can be obtained by screening of a wide range of naturally available isolates, and the efficiency of those can be improved by selection, adaptation, and genetic engineering (Singh et al., 2011). Isolation of suitable microalgae from the natural environment is the first critical step in developing oil-rich strains for further exploitation in engineered systems for the production of biodiesel feedstock (Doan et al., 2011).
Microalgae Selection for Biodiesel Production
Around 22,000–26,000 species of microalgae exist, of which only a few have been identified for successful commercial application (Norton et al., 1996). In the US Department of Energy's Aquatic Species Program (ASP) to develop microalgae as a source of biodiesel, more than 3,000 strains of microalgae from ponds and oceans have been isolated (Rosenberg et al., 2008). The selection of suitable microalgae strains is an important factor in the overall success of biofuel production from microalgae (Brennan and Owende, 2010). In the perspective of this review, the ideal algal strain for biofuel production should have high lipid productivity; be robust and able to survive the shear stresses common in photobioreactors; be able to dominate wild strains in open pond production systems; have high CO2 uptake capacity; have limited macronutrient requirements; be tolerant to a wide range in temperatures resulting from the diurnal cycle and seasonal variations; provide valuable coproducts; have a fast productivity cycle (Brennan and Owende, 2010). At present, no known algal strain is capable of meeting all these requirements concurrently. A list of biomass, lipid content, and lipid productivity of microalgae is shown in Table 2.
Source: Rajvanshi and Sharma (2012).
TAG, triacylglycerols.
Another important aspect to consider while selecting the microalgal strains is when microalgae growth is combined with CO2-biomitigation (biologically minimize adverse environmental impacts of CO2), due to the fact that microalgae have much higher growth rates and CO2 fixation abilities compared to conventional agricultural, forestry, and aquatic plants (Li et al., 2008b). Several such algal species, including species of Chlorella and Scenedesmus, are capable of CO2-biomitigation (Li et al., 2008a). Microalgal biofuel production becomes more attractive when combined with fixing industrial exhaust gases (flue gas) and integrating the cultivation of microalgae with wastewater treatment (Chinnasamy et al., 2010).
Effect of Environmental Factors
During photosynthesis, microalgae produce lipids, carbohydrates, and proteins by using nutrients and light. The relative amounts of those metabolic products are tightly connected to environmental and nutrient conditions, including temperature, pH, light intensity, CO2 levels, nutrients availability, and the existence (or nonexistence) of different organisms (Mata et al., 2013). Nonmineral nutrients, macronutrients, and micronutrients are required for algal growth. Normally, environmental conditions (particularly temperature and light) and the availability of nutrients (i.e., nonmineral nutrients, macronutrients, and micronutrients) significantly influence the microalgal biochemical composition (Fábregas et al., 2004). Further factors such as pH and the presence of toxic metals are also vital factors impacting growth and metabolism of microalgae. Usually, all these factors have an effect on photosynthesis, thus altering carbon fixation and also the distribution of carbon into different types of macromolecules. Thus, environmental factors determine composition of microalgal cell and its importance in biofuel production.
pH
One of the foremost essential factors in algal cultivation is the hydrogen ion concentration, that is, the pH, as it determines the solubility and accessibility of CO2 and required nutrients and since it can have a noteworthy impact on algal metabolism (Goldman, 1973). Hydrogen ion concentration can increase considerably in algal cultures, which is attributable to the uptake of inorganic carbon by microalgae. Most algal growth occurs in the region of neutral pH, although optimum pH is the pH of initial culture in which an alga is adapted to grow (Hansen, 2002). Shifting hydrogen ion concentration, that is, pH, in media might limit the growth of microalgae by metabolic inhibition (Goldman et al., 1982).
Remarkably, pH is one of the key factors to determine relative concentrations of the carbonaceous water species. High pH limits the availability of carbon from CO2, which consecutively suppresses the growth of microalgae (Azov, 1982; Rotatore and Colman, 1991). At higher pH, carbon for microalgae is available in form of carbonates (Rotatore and Colman, 1991). Some microalgae possess the bicarbonate uptake system, for example, Nannochloropsis gaditana (Huertas et al., 2000). The flexibility of the cell wall of microalgae increases at basic pH and prevents their rupture and inhibits autospore release, thus increasing cell cycle completion time (Guckert and Cooksey, 1990). Basic pH indirectly leads to an increase in total triglyceride accumulation in cell; however, basic pH decreases membrane-associated polar lipids due to cell cycle inhibition. In basic pH conditions, membrane lipids in Chlorella were observed to be less unsaturated (Guckert and Cooksey, 1990). Various pH ranges were tested on Tetraselmis suecica CS-187 and Chlorella sp. for growth and lipid productivity. In this study, T. suecica CS-187 showed highest biomass (320±29.9 mg/[L·day]) and lipid productivities (92±13.1 mg/[L·day]) at pH 7.5, and Chlorella sp. showed highest biomass (407±5.5 mg/[L·day]) and lipid productivities (99±17.2 mg/[L·day]) at pH 7 (Moheimani, 2013).
Acidic pH of culture media can alter nutrient uptake or induce metal toxicity and therefore have an effect on algal growth (Visviki and Palladino, 2001). Expenditure of energy is required to pump protons out of the cell for maintenance of neutral intracellular pH in an acidic pH external environment (Terry and Abadía, 1986). Alternatively, acid-tolerant microalgae, such as Chlorella saccharophila (Gehl and Colman, 1985) and Euglena mutabilis (Lane and Burris, 1981), can change intracellular pH in response to varying external pH. Within C. saccharophila, an inner pH of 7.3 was maintained for an outer pH scale of 5.0–7.5; however, decreasing the pH further to 3.0 caused reduction in cellular pH to 6.4 (Gehl and Colman, 1985). Similarly, E. mutabilis shows an inner pH varying from 5.0 (at low outer pH<3.0) to 8.0 (at high outer pH>9.0) (Lane and Burris, 1981). Demand of energy to maintain internal pH in these acid-tolerant microalgae is preserved as the internal pH decreases. This could be a mechanism for maintaining cellular metabolism such that the growth of alga is not drastically affected by an acidic environment (Gehl and Colman, 1985). Such a mechanism would offer acid-tolerant microalgae with the capability to control inner pH in response to outer pH variations, so maintaining an energy benefit over acid-intolerant species at low outer pH. Several acidophilic diatoms, such as Chlamydomonas sp. and Pinnularia braunii, accumulate storage lipids such as triacylglycerides under extremely acidic environment (pH 1) (Tatsuzawa et al., 1996; Poerschmann et al., 2004).
Temperature
Temperature is one of the foremost vital environmental factors that influence algal growth rate, cell size, biochemical composition, and nutrient requirements. Microalgae grow in a variety of temperature ranges, relying upon strain, region, and time of year. Temperatures providing optimum growth rates are stated between 20°C and 25°C for mesophilic species (Plectonema calothricoides and Phormidium luridum) but can increase for thermophilic strains, for instance 55°C for Mastigocladus laminosus or decrease down for psychrophilic strains, for instance 4°C for Mastigocladus vaginatus (Chen and Berns, 1980). Several algal species can tolerate temperatures as much as 15°C below their optimum, with low growth rates; however, a temperature of only some degrees higher than optimal can lead to cell death (Mata et al., 2010). At optimum temperature, the growth of microalgae leads to minimum cell size, and therefore, the efficiency of carbon and nitrogen utilization decreases at nonoptimal temperatures (Barsanti and Gualtieri, 2005). Usually, carboxylase activity decreases at low temperatures, and if the light environment remains unchanged, the energy produced will be higher than the required one. This disproportion creates light saturation conditions. Chlorella vulgaris, for example, is cultivated efficiently at 5°C with a lower chlorophyll content compared to cultures at 27°C (Ras et al., 2013). Temperature may play a main role in photoinhibition. Low temperature leads to reduced electron transport at a given photon flux rate owing to slower rate of CO2 fixation (Richmond, 2008).
One of the most frequently observed variations with temperature change is an alteration in the level of unsaturation of fatty acids within the lipid membrane (Moazami-Goudarzi and Colman, 2012). During a study on eight marine plankton species, fatty acids (14:0) increased from ∼4% at 10°C to >20% at 25°C, whereas PUFA (polyunsaturated fatty acids) were constantly higher at lower temperature (10°C) (Thompson et al., 1992). Dunaliella salina has revealed a substantial enhancement in fatty acid unsaturation when temperature is decreased from 30°C to 12°C (Thompson et al., 1992). Fluidity in the cell membrane decreases at low temperature. Cells then compensate by raising the levels of unsaturated fatty acids to enhance fluidity, although this addition makes the membranes more prone to damage by free radicals (Nishida and Murata, 1996). With greater fluidity, enhanced levels of unsaturated fatty acids tend to enhance the cellular membranes stability. This, sequentially, at low temperature protects the photosynthetic machinery from photoinhibition (Nishida and Murata, 1996).
However, temperature failed to have an effect on the secretion of extracellular lipids (Kalacheva et al., 2002). Similarly, N. oculata and C. vulgaris show an optimum growth at 25°C. An increase in temperature from 20°C to 25°C doubled the lipid content (from 7.90% to 14.92%) in N. oculata. Increasing the growth temperature from 25°C to 30°C reduced the lipid content in C. vulgaris from 14.71% to 5.90% (Converti et al., 2009). Increasing the temperature beyond the optimum decreases protein synthesis and consequently leads to a decrease in growth rate (Konopka and Brock, 1978).
Light
Light is the energy source in the photoautotrophic growth phase, and organisms utilize light energy to convert CO2 to organic compounds—sugars. Microalgae need light and dark regimes for photosynthesis. The light phase provides adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide phosphate-oxidase (NADPH) (Atta et al., 2013; Kim et al., 2014). The intensity of light affects the growth of microalgae through its impact on photosynthesis (Stockenreiter et al., 2013). The efficient production of algal biomass relies on the optimal provision of light energy to all cells within the culture. The majority of algal growth systems become light limited at high cell densities. Light penetrates only a few centimeters into a dense algal culture due to absorption and shading by the cells (Richmond, 2008). The average provision of light is linked to reactor depth or diameter, mixing, and cell concentration. A larger surface area to volume ratio, generally achieved through areas of thin paneling or narrow tubing, results in higher light provision. At low light intensities, photosynthetic efficiency is highest. At high light levels, although the photosynthetic rate may be faster, there is a lesser amount of efficient use of absorbed light energy.
Above the saturation point, damage to photosynthetic machinery can occur in a process known as photoinhibition (Scott et al., 2010). Within a dense culture exposed to direct sunlight, cells at the surface are likely to be photoinhibited, whereas those at the middle of the reactor are in the dark. The rate of light–dark cycling has been reported to affect algal productivity (Grobbelaar, 2000). When the light is supplied in short bursts, microalgae are less likely to become photoinhibited because the photosystems have time to recover during the dark period. Additionally, a recent study has shown that blue LED was the effective source for the biomass production and the accumulation of lipid content owing to its high potency for photosynthesis. C. vulgaris exhibited its highest growth rate and lipid content under the influence of blue LED (Atta et al., 2013).
Salinity
Salinity is another significant factor that changes the biochemical composition of algal cells (salinity refers to sodium chloride concentration, unless otherwise specified). Culturing microalgae at lower or higher salinity levels than their natural (or adapted) levels can alter growth rate and vary composition. Microalgal growth is retarded under salinity stress condition owing to the buildup of compatible solutes, such as glycine and proline, to balance the outer salt concentrations (Hellebust et al., 1989). Compatible solutes are produced inside the cells under osmotic stress environment, and they act as osmoprotectants to stabilize enzymes. The level of saturated fatty acid in microalgae is reduced as the concentration of NaCl is increased, whereas the level of highly unsaturated fatty acid is enhanced (Kirroliaa et al., 2011). Reduction in photosynthetic activity usually is examined as a possible consequence of salt stress due to limitations in photosynthetic transport of electron and partial stomatal closure (Zhang et al., 2010). For instance, in microalgae, lipid content increases at higher salinity (Fabregas et al., 1984; Zhila et al., 2011). In Dunaliella tertiolecta, an increase in intracellular lipids (60% to 67%) and triglyceride concentrations (40% to 56%) with an increase in NaCl concentrations from 0.5 to 1.0 M was studied (Takagi and Yoshida, 2006). Increasing the salinity level in cultures of B. braunii resulted in an increase in growth rate, carbohydrate content, and lipid content; however, the utmost biomass concentration was accomplished at the lowest salinity level (Rao et al., 2007). An additional study with a similar alga reported a reduction in protein content with unaffected carbohydrate and lipid content, with an increase in salinity (Vazquez-Duhalt and Arredondo-Vega, 1991). Lipid content was increased up to 34% dry cell weight in Nannochloropsis salina when N. salina grew at 22 practical salinity unit (PSU) until stationary phase is reached and then increased to 34 PSU (Bartley et al., 2013).
Nutrients
Nonmineral nutrients required for algal growth include carbon, hydrogen, and oxygen. Macronutrients contain nitrogen, phosphorus, sulfur, and metallic elements such as magnesium and potassium. Micronutrients, including manganese and iron, are also essential in little amounts, whereas the additional components, such as zinc, boron, molybdenum, cobalt, and copper, are crucial trace components (Mata et al., 2013). Considerable variation within the biochemical composition during conditions of nutrient limitation can be studied in microalgae, depending upon which nutrient is limited and to what level. For growth and metabolism of algal cells, nitrogen and phosphate are two vital macronutrients. Phosphorus is the main element of phospholipids. Limitation of fundamental nutrients shifts the metabolic pathway of the microalgae (Richmond, 2008). For instance, nitrogen and phosphorus starvation transfers the lipid metabolism from membrane lipid synthesis to neutral lipid storage. This enhances the entire lipid content of Chlorophyta. Specific effects of key nutrients are discussed below.
Carbon
Carbon, hydrogen, and oxygen are three crucial nonmineral nutrients. Abundance of hydrogen and oxygen within the media for microalgae cultures means that their availability is not a challenge to growth or cellular metabolism. Carbon is a key nutrient that has to be provided. It is essential for photosynthesis and consequently growth and reproduction of microalgae. Carbon fixed by the microalgae can finish up in three destinations; it will be used (1) as an energy source, (2) for respiration, or (3) as a raw material within the formation of further cells (Berman-Frank and Dubinsky, 1999). Reduction in the carbon fixation rate results in a decrease in growth rate of microalgae. Carbon can be used in the form of glucose or acetate for heterotrophic growth and in form of CO2, carbonate, or bicarbonate for autotrophic growth. Relying upon pH, temperature, and nutrient content, CO2 in water could also be there in any of these forms:
The addition of organic carbon into heterotrophic cultures of microalgae is able to enhance both microalgal biomass productivity and lipid content (Xin et al., 2011; Silaban et al., 2014). A major effect of CO2 on the composition of the alkenones and polyunsaturated fatty acids was reported. Specifically, lower CO2 concentrations led to an increase in PUFA. Increasing CO2 concentration was additionally observed to enhance the amount of fatty acid accumulation in D. salina (Muradyan et al., 2004). Similarly, the study conducted on N. salina shows that the growth rate and lipid content in microalgae increases when the culture medium was supplied with CO2-enriched air than air alone (Arudchelvam and Nirmalakhandan, 2012). Two Chlorophyta, T. suecica and Chlorella sp., when grown using CO2 as inorganic carbon source achieved highest biomass and lipid productivity: 51.45±2.67 mg/[L·day] biomass and 14.8±2.46 mg/[L·day] lipid productivity for T. suecica, and 60.00±2.4 mg/[L·day] biomass and 13.70±1.35 mg/[L·day] lipid productivity for Chlorella sp (Moheimani, 2013). Highest algae productivity of 24.7 g/[m2·day] was measured when wastewater treatment high rate algal ponds combined with CO2 (Park and Craggs, 2010).
Nitrogen
Nitrogen is a key constituent of all structural and functional proteins within the algal cells and accounts for 7% to 20% of cell DW (Richmond, 2008). Foremost effects of nitrogen deficiency in algal culture consist of the enhanced biosynthesis and accumulation of lipids (Muradyan et al., 2004; Demirbas, 2010; Surendhiran and Vijay, 2014) and triglycerides (Takagi et al., 2000; Stephenson et al., 2010; Yang et al., 2014) with a concurrent decrease in protein content. Biomass concentration and lipid productivity was reached at highest level with a nitrogen concentration; it was raised ten times that of the standard medium in D. tertiolecta (Mata et al., 2013). Attempts to enhance lipid concentration through nitrogen limitation should be carefully estimated to confirm high lipid productivity. Nitrogen starvation additionally affects the lipid synthesis and reduces chlorophyll synthesis, resulting in excess carotenoids within the cells. It had been observed that Haematococcus pluvialis accumulate large amounts of carotenoids, astaxanthin, and its acylesters (up to 13 w/w) during growth under nitrogen-depleting conditions (Harker et al., 1996). Huang demonstrated the effect of nitrogen on Tetraselmis subcordiformis SHOU-S05, N. oculata SHOU-S14, and Pavlova viridis SHOU-S16; growth rates were lowest for all three microalgae in the medium without nitrogen and highest lipid contents were observed at 0.22 mM (Huang et al., 2013).
Phosphorus
Phosphorus is a considerable component needed for normal growth and development of microalgae (Richmond, 2008). Limitation of phosphorus results in lipid accumulation. It was observed that total lipid content in Scenedesmus sp. was increased from 23% to 53% with a decrease in initial total phosphorus (as phosphate) concentration of 0.1 from 2.0 mg/L (Xin et al., 2010). When grown on phosphorus-deficient culture media, the lipid content in Picochlorum sp. increased by 2.5-fold (El-Kassas, 2013). Phosphatidylglycerol (PG), which is one among the four main glycerolipids comprising membrane lipids in chloroplasts, was examined to decrease with phosphorus limitation in Chlamydomonas reinhardtii (Sato et al., 2000).
Trace metals
Trace metals are metals present in algal cells in tremendously small quantities (<4 ppm), and yet they are vital elements of phycophysiology. Iron (Fe), magnesium (Mg), potassium (K), manganese (Mn), and zinc (Zn) are the five most vital trace metals essential to microalgae for varied metabolic functions (Mata et al., 2013). Deficiencies in trace metals can limit the growth of microalgae, whereas high metal concentrations (above the toxicity threshold) could inhibit growth, impair photosynthesis, deplete antioxidants, and damage cell membrane.
Iron is a significant trace metal for normal growth and functioning of photosynthesis and respiration in microalgae (Richmond, 2008). Iron limitation considerably lowers photosynthetic electron transfer, leading to reduced NADPH formation. An increase in lipid content was observed in C. vulgaris culture containing high concentrations of iron (Liu et al., 2008). D. tertiolecta has revealed a substantial enhancement in the maximum lipid productivity to almost the double, from 14.6 to 28.0 mg/[L·day], when raising the Fe concentration 10 times in the culture medium (Mata et al., 2013).
Although there are some nonessential metals (e.g., cadmium, lead, and chromium), which might inhibit several metabolic processes even in minute quantities, there are some essential elements (e.g., zinc and copper) that when in excess can cause toxicity (Campanella et al., 2001). In comparison with the standard medium biomass, productivity of D. tertiolecta is reduced when zinc and manganese are added to the culture medium (Mata et al., 2013). Cadmium (Cd) is mostly toxic to algal cells. Although cadmium has no biological importance in a living cell, it is taken up by marine algae in the form of complexes with organic matter and is absorbed on both organic matter and inorganic matter in ionic form (Wong et al., 1979).
Summary of environmental factors on biochemical composition of microalgae is given in Table 3.
FA, fatty acid.
Conclusions
Selection of resilient and productive species and optimizing cultivation conditions for enhancing lipid content in microalgae are key considerations in biodiesel production. High lipid-producing microalgal strains are suitable for biodiesel production. Various nutrient and environmental factors affects on lipid content and biomass productivity of microalgae, such as pH, temperature, light, nitrogen, and phosphorus. Depending on the microalgal strain, the percentage of lipid content can be increased under stress conditions, for example, low temperature or reduced nitrogen and phosphorus in the medium.
Among all the environmental factors studied, nitrogen plays an important role and widely studied for increasing the lipid accumulation in microalgal cell. Besides nitrogen, temperature is also a crucial environmental factor for enhancing lipid content. However, along with temperature, other factors, such as pH and light, are difficult to maintain while cultivating microalgae at large scale. Although exact combination of different environmental factors (such as temperature, pH, photoperiod, and nitrogen) for commercial scale cultivation for biodiesel production will differ from species to species, this depends on nutrient availability and environmental conditions.
Future Potential
With advances in biotechnology and bioinformatics, a huge volume of genetic information and genetic manipulation tools are now available, rendering studies of interaction effects tractable. Continued research will lead not only to an enhanced understanding of basic algal cell biology, but it will also aid in the development of more accurate predictive models for microalgae growth. Predictive models can be used for the development of automated optimal control systems for managing microalgae growth in large-scale production systems.
Further research is focused on cost reduction of cultivating microalgae on small and large scale. Besides growing microalgae under well-defined conditions and provided with ideal growth substrates, microalgae can be grown on waste water, swage, industrial waste, CO2 emitted by industries, or industrial byproducts. In recent research, it was shown that the alga Schizochytrium limacinum could be grown on crude glycerol, which is a main byproduct of commercial biodiesel production. The composition of microalgae produced from biodiesel-derived glycerol was similar to that of commercial microalgae.
A cultivation system is essential for large-scale cultivation, depending on strain and suitable parameters required for lipid productivity, photobioreactors, or other cultivation systems that should be designed.
Footnotes
Author Disclosure Statement
No competing financial interests exist.