
Abstract
Abstract
Microalgae-based systems for wastewater treatment, biofuel production, and nutrient recovery require effective solid/liquid separation processes that frequently include coagulation and flocculation steps where discrete particle (i.e., microalgae) suspensions are destabilized to initiate particle agglomeration, followed by conglomeration into larger flocs, respectively. The choice of coagulant affects process economics and sustainability. This study investigated the use of natural cationic polymers (chitosan and cationic starches with varying degrees of substitution) and the synthetic cationic polymer (polydiallyldimethylammonium chloride [pDADMAC]) to flocculate Chlorella protothecoides and Nannochloropsis salina. Algae surface charge and flocculation efficiency were examined in response to variable pH, coagulant dosage, cell concentration, and salinity. Overall effectiveness of coagulation, flocculation, and sedimentation is referred to as flocculation efficiency and is determined by measuring the percentage decrease in optical density of the microalgae suspensions. Cationic starch with a degree of substitution (DS) of 0.5 and chitosan neutralized the charge on C. protothecoides at lower dosages compared with pDADMAC. C. protothecoides flocculation efficiencies >95% were achieved with dense cultures (1 g/L algae dry weight) at cationic starch (DS 0.5) and chitosan dosages of 0.02 g/g algae dry weight. Zeta potential measurements indicated that complete charge neutralization was not necessary for maximum flocculation. Of the polymers tested, zeta potential analysis indicated that only chitosan was sensitive to pH. Maximum flocculation efficiency of N. salina was achieved through chitosan precipitation and subsequent sweep floc at pH 8.0. However, cationic starch (DS 0.5) and pDADMAC optimum doses were lower than that required for N. salina flocculation with chitosan.
Introduction
S
One or more solid/liquid separation steps are used to dewater algae on a large scale. Energy and cost-efficient harvesting operations frequently apply flocculation combined with sedimentation or flotation as primary steps, followed by centrifugation or filtration (Schenk et al., 2008). Flocculation involves two processes, destabilization of a particle suspension, followed by particle aggregation to form large flocs that settle easily. Algal destabilization can be induced by neutralizing the cell surface charge through treatments with cationic chemicals or polymers, interparticle bridging using long-chain polymers, enmeshment of algal cells in precipitates, and double layer compression (Tenney et al., 1969; Henderson et al., 2008; Schlesinger et al., 2012).
Several flocculation methods, including bioflocculation, autoflocculation, and flocculation using inorganic salts or polymeric coagulants exist. Bioflocculation is induced by extracellular polymeric substances, secreted by microorganisms under stress or during lysis, which cause aggregation through adhesion, bridging, and hydrophobic interactions (Tian et al., 2006). Autoflocculation occurs when CO2 consumption by algae during photosynthesis results in high pH, subsequently initiating precipitation of calcium and magnesium salts. When cations (i.e., Ca or Mg) are present in excess, these precipitates may carry a positive charge making conditions favorable for algae aggregation and coprecipitation (Sukenik and Shelef, 1984; Vandamme et al., 2012). Both bio- and autoflocculation processes require no chemical additives; however, they are not reliable on a large scale and may cause undesirable changes in cell composition due to stress (Schenk et al., 2008; González-Fernández and Ballesteros, 2011). Alum, ferric sulfate, and ferric chloride are inorganic salts of polyvalent cations commonly used to induce coagulation by charge neutralization, but can be toxic and can contaminate the algal biomass (Tripathy and De, 2006).
Polymeric coagulants include both synthetic and natural polymers. In comparison with inorganic salts, synthetic polymers such as polyacrylamides are more efficient, but not preferred due to their low biodegradability (Krentz et al., 2006). Natural organic polymers, including chitosan and cationic starches, are biodegradable and less toxic than synthetic polymers, and thus preferred for wastewater treatment and microalgae separation processes (Lavoie and de la Noue, 1983; Divakaran and Pillai, 2002; Sun et al., 2007; Vandamme et al., 2010; Anthony and Sims, 2013). Both synthetic and natural polymers can facilitate algae aggregation and flocculation through attractive electrostatic interactions and bridging between cells (Tripathy and De, 2006). In brackish and marine algae applications, flocculation effectiveness can be limited by high salt concentrations that can alter the molecular configuration and charge density of cationic polymers (Bilanovic et al., 1988; Sukenik et al., 1988; Molina Grima et al., 2003).
Natural cationic polymers, including chitosan and cationic starches, are intrinsically well suited for microalgae coagulation and flocculation. Chitosan, derived from alkaline deacetylation of chitin has a high cationic charge density in acidic conditions due to the presence of protonated amino groups (Renault et al., 2009). At neutral and alkaline pH, chitosan can precipitate with coprecipitation of algae (Renault et al., 2009). Lavoie and de la Noüe (1983), Morales et al. (1985), and Divakaran and Pillai (2002) reported >90% biomass removal, determined using optical density (OD) measurements of both freshwater and saltwater algae cultures by flocculation with chitosan, followed by gravity separation. These studies showed microalgae flocculation with chitosan was sensitive to pH, algae strain, and cultivation methods.
Cationic starches are prepared by addition of cationic functional groups (e.g., quaternary ammonium cation) to the hydroxyl groups on starch molecules. The level of cationic substitution is defined as degree of substitution (DS). Vandamme et al. (2010) and Anthony and Sims (2013) have evaluated cationic starches at both low DS (0.11, 0.15) and high DS (0.82, 1.32) to separate freshwater algae cultures (Scenedesmus obliquus) at initial biomass concentrations ranging from 0.1 to 0.5 g/L. Their studies showed that biomass separation effectiveness was proportional to the DS with biomass removal efficiencies of 80–90%. Vandamme et al. (2010) further showed that cationic starch with low DS was ineffective for flocculating the marine algae Nannochloropsis sp. Cationic starches are increasingly being considered in wastewater and algae harvesting processes due to their effectiveness. However, polymer production costs are proportional to the DS and need to be taken into consideration for sustainable process designs (Vandamme et al., 2010).
These earlier studies have demonstrated the use of natural polymers as effective coagulants for algae flocculation. However, these studies are limited and more research is required for a more comprehensive understanding of algae-polymer charge interactions, salinity effects on flocculation efficiency, and natural and synthetic coagulant performance. This study evaluates the use of chitosan and cationic starches (DS 0.5, 0.2) to flocculate microalgae suspensions of Chlorella protothecoides (freshwater species) and Nannochloropsis salina (brackish-water species). The effectiveness of cationic starches to aggregate algae depends on the DS; therefore, cationic starches with a DS between the range tested by Vandamme et al. (2010) and Anthony and Sims (2013) were chosen for this study. Flocculation characteristics of natural cationic polymers are compared with a commonly used synthetic cationic polymer, polydiallyldimethylammonium chloride (pDADMAC). Experimental observations of the zeta potential and flocculation efficiency were used to characterize cell aggregation mechanisms and provide insight regarding the coagulant choice. Results show that cationic starch (DS 0.5) and chitosan are potential coagulants for microalgae flocculation where efficiencies of cationic starch and chitosan are dependent on the DS and pH, respectively.
Materials and Methods
Microalgae growth
C. protothecoides and N. salina were chosen to study the effectiveness of cationic polymers to flocculate algae grown in freshwater and brackish-water environments, respectively. C. protothecoides (UTEX 256) cells were cultured in Bolds' basal media (Bischoff and Bold, 1963). N. salina obtained from Texas AgriLife Research (College Station, TX) was grown in 15 g/L saltwater prepared with Crystal Sea Marinemix (Marine Enterprises International, Baltimore, MD) and supplemented with Guillard's f/2 nutrients obtained from Florida Aqua Farms, Inc. (Dade City, FL). The f/2 nutrient solution was added using sterile filtration at a dose of 0.4 mL for 1 L saltwater. The cultures were grown in 250-mL Erlenmeyer flasks and transferred into 20-L batch reactors. The batch reactors were exposed to continuous light from fluorescent bulbs (100 μmol/[m2·s]) to maximize cell growth. The reactors were sparged with 0.2 μm filtered air enriched with 5% CO2 at a flow rate of 1 L/min. Algal growth was monitored by measuring total suspended solids, and flocs were qualitatively characterized using a microscope. Biomass concentrations are expressed as dry weight (DW) and ash-free dry weight (AFDW) for freshwater (C. protothecoides) and brackish-water (N. salina) cultures, respectively (Steinman and Lamberti, 1996). Algal growth phase affects cell surface charge, hence the effective coagulant dosage will vary accordingly. Lower optimum coagulant doses have been reported for the stationary phase compared with the log growth phase (Tenney et al., 1969; Edzwald and Wingler, 1990). To minimize growth phase effects on cell surface charge, cultures in the stationary phase were used for zeta potential and flocculation experiments.
Zeta potential measurements
Zeta potential is a measure of electric potential in the interfacial double layer and is dependent on the particle surface charge, adsorbed layer at the particle interface, and the nature and composition of the surrounding medium in which the particle is suspended (Atkins, 1990). Zeta potential measurements were made using a zetaPALS (Brookhaven Instruments, Holtsville, NY) to assess cell suspension stability as a function of pH, coagulant dosage, and salinity. Dilute algal cultures were used to prevent formation of large flocs that could settle during zeta potential measurements. C. protothecoides dilutions were made with appropriate growth media to attain 0.020 g/L initial cell concentration. Sample pH and coagulant dosage were adjusted to desired levels and vortexed. All measurements were taken 30 min following pH adjustments and coagulant additions to allow sufficient time for the zeta potential to stabilize (López-León et al., 2005). C. protothecoides zeta potentials were evaluated as a function of media pH and coagulant dosage. All zeta potential experiments were performed in triplicate.
N. salina zeta potentials were evaluated as a function of the coagulant at three salt concentrations (0.05, 0.25, and 0.5 g/L). Samples were prepared by centrifuging 50 mL aliquots of the culture media at 3,500 rpm for 15 min. Following centrifugation, the supernatants were decanted. The concentrated cultures were then resuspended in seawater at the respective experimental salinities. The zeta potential was measured at low salt concentrations, relative to the flocculation experiments, as a surrogate measure of the cell's electric potential to elucidate coagulation mechanisms. The low salinity modification to the zeta potential protocol was needed to minimize double layer compression that can invalidate zeta potential measurements through a charge shielding effect at higher ionic strengths. It can be shown that the reciprocal Debye length (κ−1) or characteristic double layer thickness is inversely proportional to ionic strength as shown by
where ɛ0=permittivity of free space, ɛr=relative permittivity of water, R=gas constant, T=absolute temperature, F=Faraday's constant, and I=ionic strength (Sterling et al., 2004). Furthermore, adverse effects to zeta potential measurements may include joule heating due to high conductivity (Corbett et al., 2012).
Cationic polymer coagulants
Coagulants investigated were natural (chitosan, cationic starch) and synthetic cationic polymers (pDADMAC). Chitosan (molar mass 190–300 kDa, degree of deacetylation 75–85%) and pDADMAC (molar mass 200–350 kDa, 20 wt.%, density 1.04 g/mL) were obtained from Sigma-Aldrich (St. Louis, MO). Chitosan stock solution was prepared in 1% acetic acid and was left overnight to dissolve. Two commercially available cationic starches commonly used in the paper making industry were used. Cationic corn starches with 0.2 and 0.5 DS were obtained from Hestion Industry (Liaocheng, China) and Grain Processing Corporation (Muscatine, IA), respectively. Cationic starch (DS 0.2) stock solution was prepared by boiling the starch powder in deionized water. Cationic starch with DS (0.5) was supplied as 40% by weight starch solution in water. The chemical composition of the commercial cationic starches used is proprietary, but in general cationic starches are prepared by substituting hydroxyl groups present on starch with amine or quaternary ammonium groups (Anthony and Sims, 2013; Khalil and Aly, 2013).
Flocculation experiments
Jar tests were performed in a Phipps and Bird (Richmond, VA) six-paddle stirrer using 1-L beakers, each containing a liter of algal suspension. Culture pH was adjusted using 0.1 M HCl or 0.1 M NaOH and the required coagulant dosage was added. The culture was mixed at 120 rpm for 1 min, followed by 30 min of slow mixing at 60 rpm. The flocs were then allowed to settle undisturbed for 30 min. Samples of the supernatant were taken from a fixed height in the jar corresponding to the 0.8 L volume mark on the beaker. For the purpose of this article, flocculation efficiency is defined as the overall effectiveness of coagulation, flocculation, and sedimentation. Flocculation efficiency was estimated by measuring the OD of cell culture before flocculation and OD of the supernatant sample after settling at 680 nm (Lee et al., 1998). Optical density (OD680) of the sample was measured using a spectrophotometer (Ocean Optics, Dunedin, FL). Flocculation efficiency was calculated by the following equation (Papazi et al., 2010):
where A is OD680 of supernatant liquid after settling and B is OD680 of initial culture.
The effect of pH, coagulant dosage, and initial algae biomass concentration on flocculation efficiency was studied. Jar tests were performed in triplicate. Statistical analysis of results was done using one-way analysis of variance (ANOVA) and Tukey's test (OriginPro 8, OriginLab, Northampton, MA).
Results and Discussion
Effect of pH and polymer dosage on the zeta potential of C. protothecoides
Zeta potentials of C. protothecoides cells coagulated with cationic starches (DS 0.5, DS 0.2), chitosan and pDADMAC, were evaluated in response to variable pH (Fig. 1) and polymer dosage (Fig. 2). The effect of pH was measured at a constant polymer dose of 0.075 g/g algae DW. The zeta potential of C. protothecoides cells without any polymer addition was measured as control. Figure 1a–c shows that C. protothecoides cells (control) are negatively charged in the pH range 4.0–8.0, and the charge is independent of pH in this range (ANOVA p=0.83). Of the two cationic starches tested, positive zeta potentials (>25 mV) were measured at all pH levels in C. protothecoides suspensions coagulated with DS of 0.5 (Fig. 1a). Conversely, suspensions treated with cationic starch (DS 0.2) resulted in negative zeta potentials that were not significantly different from the control suspensions. These observations demonstrate that the charge neutralization capacity of the cationic starches is proportional to the DS. Statistical analysis indicates that the zeta potential of C. protothecoides–cationic starch aggregates was not significantly dependent on solution pH for both starches (Tukey's test p>0.05), suggesting the presence of quaternary ammonium groups on the starch molecule (Anthony and Sims, 2013). The zeta potential of C. protothecoides suspensions treated with chitosan varied with solution pH, with zeta potentials shifting toward neutral in acidic media (Fig. 1b). The positive charge on chitosan can be attributed to protonation of −NH2 groups on chitosan (pKa ∼6.5) in an acidic environment and change in the degree of protonation with [H+] concentration (Rinaudo et al., 1999). The protonated amine groups allow chitosan to strongly adsorb on negative regions on the algal surface, resulting in less negative zeta potentials of aggregates in acidic pH. In basic pH, the charge on the chitosan polymer is low; therefore, no significant difference in the zeta potential of control and cells coagulated with chitosan was observed at pH 7.0 and 8.0. In the pH range 4.0–8.0, the zeta potential of aggregates with pDADMAC was not significantly influenced by solution pH (ANOVA p=0.78) (Fig. 1c). As pDADMAC is a homopolymer of diallyldimethylammonium chloride, a quaternary ammonium compound, a positive charge on the polymer is maintained even at high pH.
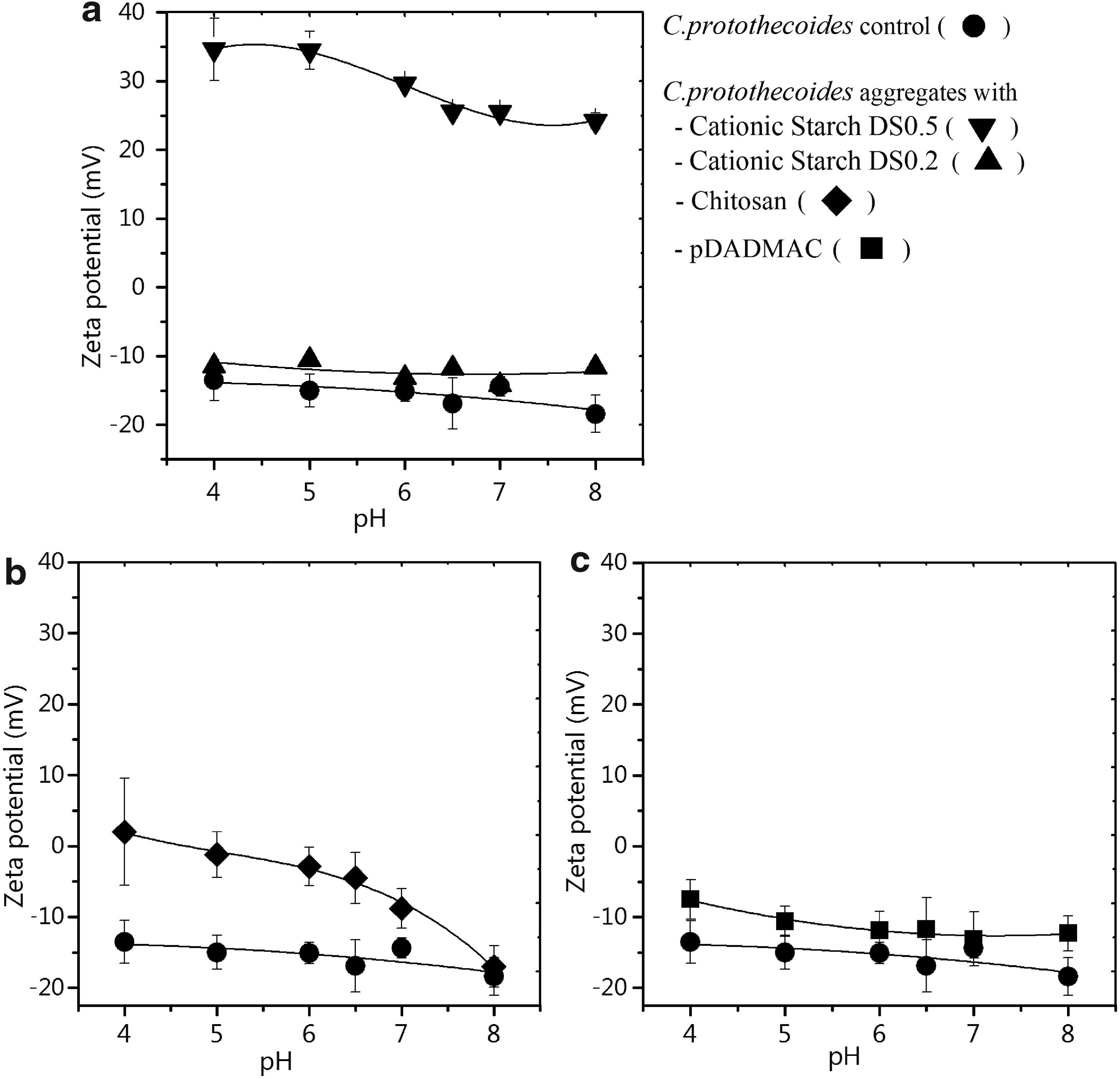
Zeta potential of Chlorella protothecoides aggregates with 0.075 g/g dosage of cationic polymer as a function of pH.

Zeta potential of C. protothecoides at pH 6.0 as a function of polymer dosage.
Comparison of Figure 1a–c indicates that the zeta potential of C. protothecoides flocculated with the cationic polymers tested was not significantly dependent on solution pH, except for chitosan. Zeta potentials show that in acidic media, chitosan are more cationic and able to reduce the cell surface charge magnitude (Fig. 1b). However, the use of acid to destabilize algal suspensions may contaminate the algal biomass and add to the overall processing cost. To minimize acid use and still maintain pH below pKa of chitosan, pH 6.0 was selected for dosage studies with C. protothecoides.
The zeta potential of C. protothecoides was evaluated as a function of cationic polymer doses between 0.001 and 0.500 g/g algae DW (Fig. 2). Zeta potential measurements show that charge neutralization of C. protothecoides at pH 6.0 occurred at doses of 0.030, 0.080, and 0.100 g/g for cationic starch (DS 0.5), chitosan, and pDADMAC, respectively (Fig. 2). The doses at which charge neutralization occurred are indicative of the relative capacity of the polymers to destabilize the algal suspensions. Complete charge reversal, where the zeta potential shifted from negative to positive, was observed at increased polymer doses. In the range of 0.001–0.500 g/g dosage, the zeta potential of C. protothecoides cells coagulated with cationic starch (DS 0.2) remained negative, indicating that the level of cationic substitution of the DS 0.2 polymer was insufficient to neutralize the algal suspension under the conditions tested.
Effect of salinity on the zeta potential of N. salina cells
To study the effect of salinity on flocculation characteristics of cationic polymers, experiments were performed with N. salina cultures grown in brackish water. Zeta potential measurements were taken at three salinites (0.05, 0.25, and 0.50 g/L), two pH values (6.0 and 8.0 representing below and above pKa of chitosan), and at a constant cationic polymer dosage of 0.075 g polymer/g algae AFDW (Fig. 3a–c). The zeta potential of N. salina cells without coagulant was measured as control. Zeta potentials of N. salina control suspensions and suspensions treated with cationic starch (DS 0.2) were negative under all experimental conditions and were not influenced by pH. Increasing the salinity significantly reduced the magnitude of negative zeta potentials at the highest salt concentration of 0.50 g/L (Fig. 3a). Figure 3b shows that zeta potentials obtained with cationic starch (DS 0.5) at pH 6.0 and 8.0 were similar; reconfirming that pH had no significant effect on the cationic starch zeta potential. Zeta potentials of N. salina cells coagulated with chitosan and cationic starch (DS 0.5) were positive, and the potentials reduced significantly (p<0.05) for both these polymers with increasing salt concentration (Fig. 3b). At pH 8.0, chitosan was observed to precipitate, resulting in erratic zeta potential measurements; therefore, the data are not presented. The magnitude of the zeta potential reduced by ∼10 mV as salinity increased from 0.05 to 0.50 g/L for N. salina suspension prepared with both chitosan and cationic starch (DS 0.5) (Fig. 3b), suggesting the occurrence of double layer compression and the subsequent shielding of surface potential that can occur at higher ionic strengths (Crittenden et al., 2012). In contrast, the zeta potential of N. salina cells treated with pDADMAC was not significantly influenced by the salt concentration in the range of 0.05–0.50 g/L (ANOVA p=0.21) (Fig. 3c). The minimal effect of salt on the pDADMAC zeta potential at the salinities tested may be due to cyclopentane rings in the polymer backbone providing it a stiff structure (Gao et al., 2001). In addition, the zeta potential of N. salina treated with pDADMAC was not influenced by pH (Fig. 3c).

Effect of salinity on zeta potential of Nannochloropsis salina cells and cells coagulated with cationic polymers at 0.075 g polymer/g algae ash-free dry weight (AFDW).
Flocculation of C. protothecoides with cationic polymers
Algae biomass concentrations can vary largely from 0.5 to 5 g/L depending on the method used for cultivation (Molina Grima et al., 2003), it is therefore important to evaluate flocculation efficiency in response to variable initial cell concentrations. C. protothecoides culture was diluted with Bolds' basal media to attain initial biomass concentrations of 0.95, 0.50, and 0.10 g/L algae DW concentrations. The biomass concentrations of the prepared dilutions were within ±0.05 g/L. At each biomass concentration, the effect of the cationic polymer dose on flocculation efficiency was studied at pH 6.0. The results for the four polymers tested are presented in Figure 4a–d.
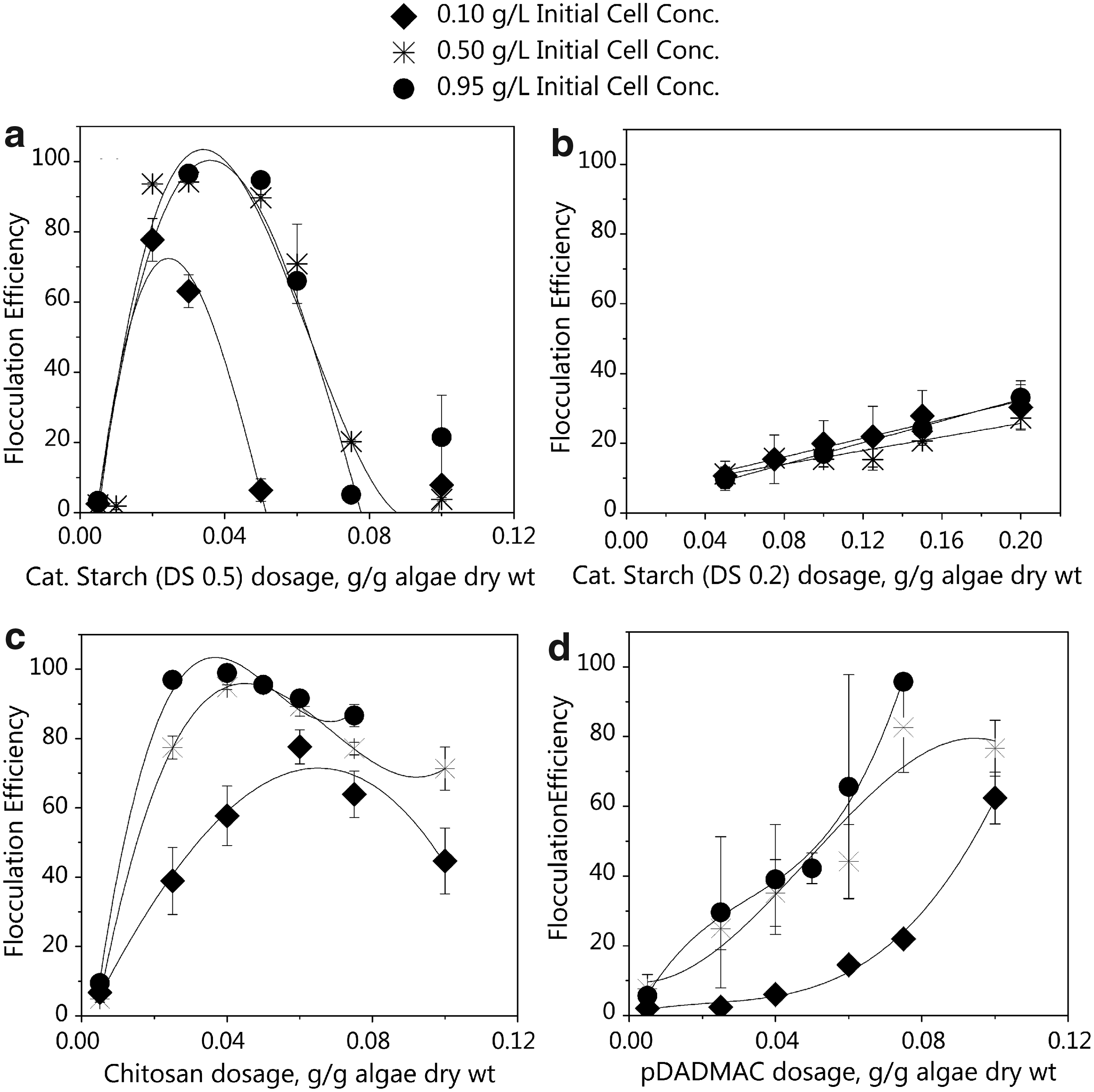
Effect of cationic polymer dosage and initial cell concentration on flocculation of C. protothecoides at pH 6.0.
Observed flocculation efficiency with cationic starch (DS 0.5) increased as the polymer dose increased from 0.005 to 0.020 g/g algae DW, with a maximum flocculation efficiency at 0.020 g/g algae DW dosage (Fig. 4a). At dosages above 0.030 g/g algae DW, flocculation efficiency declined. At the optimum cationic starch (DS 0.5) dosage (0.020 g/g algae DW), the zeta potential of C. protothecoides aggregates was −10 mV. At higher doses (>0.03 g/g), zeta potentials were positive (>+5 mV) (Fig. 2), resulting in low flocculation efficiencies due to particle restabilization. At a fixed polymer dose, flocculation efficiency was proportional to the initial algae biomass concentration. For initial biomass concentrations of 0.95, 0.50, and 0.10 g/L, the observed maximum flocculation efficiencies were 96%, 94%, and 78%, respectively. Efficient flocculation at the higher initial biomass concentration can be explained using Smoluchowski's coalescence equation, which shows flocculation kinetics are proportional to the initial particle concentration (Lee et al., 2000; Sterling et al., 2004). Flocculation efficiency with cationic starch having a lower DS (0.2) increased with increase in polymer dosage (Fig. 4b). In comparison with cationic starch (DS 0.5), flocculation efficiencies obtained with cationic starch (DS 0.2) were much lower, with a maximum efficiency of 33% at the highest dosage (0.20 g/g algae DW), and biomass concentration (0.95 g/L). Effectiveness of cationic starch with a higher DS is consistent with zeta potential observations where cationic starch with DS of 0.5 effectively neutralized the cell surface charge compared with DS of 0.2 (Fig. 2). Apart from DS, flocculation properties of cationic starch are influenced by other factors, such as location of substitutions, molecular weight of polymers, steric configuration, and the amylose-to-amylopectin ratio (Vandamme et al., 2010).
The effect of chitosan dosage on C. protothecoides flocculation was similar to cationic starch (DS 0.5) where either underdosing (<0.040 g/g algae DW) or overdosing (>0.060 g/g algae DW) of the coagulant resulted in reduced flocculation efficiencies (Fig. 4c). At optimum chitosan dosages (between 0.040 and 0.060 g/g algae DW) for the three initial biomass concentrations, zeta potentials were between −10 and −5 mV. Similar to cationic starch (DS 0.5), flocculation efficiency of chitosan was proportional to the initial biomass concentration with maximum flocculation efficiencies of 99%, 95%, and 78% being observed at initial biomass concentrations of 0.95, 0.50, and 0.10 g/L, respectively. Likewise, pDADMAC flocculation efficiency was proportional to both the coagulant dosage and initial biomass concentration (Fig. 4d). At 0.95 and 0.50 g/L biomass concentrations, optimum pDADMAC dosage was 0.075 g/g DW and achieved flocculation efficiencies were 96% and 83%, respectively. At lower biomass concentrations of 0.10 g/L, maximum of 63% efficiency was achieved at the pDADMAC dose of 0.100 g/g DW. Zeta potentials at optimum pDADMAC dosage of 0.075 and 0.100 g/g algae DW were −10 and −8.75 mV, respectively.
Analysis of flocculation efficiency and zeta potential observations demonstrate that the zeta potential must be close to neutral, not necessarily zero, for optimum algae flocculation. A similar observation that a zeta potential close to zero is sufficient for successful algae removal using alum-based coagulants has been reported (Edzwald and Wingler, 1990; Henderson et al., 2008; Garzon-Sanabria et al., 2012). According to these studies, at moderately low surface charge, cells can overcome the repulsive forces and aggregate on collision due to particle motion. C. protothecoides flocs generated with chitosan in flocculation experiments were sticky and larger compared with flocs formed by cationic starch and pDADMAC (Fig. 5a–c). During the 30 min settling period, chitosan aggregates settled quickly due to their large size, whereas pDADMAC flocs were much smaller and did not settle well. These aggregates in the supernatant resulted in large variability in measured OD values for pDADMAC–C. protothecoides aggregates (Fig. 4d).

Images of Chlorella aggregates with cationic polymers.
Flocculation of N. salina with cationic polymers
Flocculation efficiencies of N. salina measured as a function of the cationic polymer dosage, at initial cell concentration of 0.1 g/L AFDW, growth media salinity of 15 g/L, and pH 8.0 are presented in Figure 6a–c. N. salina control cultures in the high ionic strength media (i.e., 15 g/L salinity) produced stable biomass suspensions, indicating that repulsive energy between discrete particles (i.e., cells) was greater than the attractive Van Der Waals forces. Addition of the coagulant resulted in algal flocculation with maximum efficiencies of 89%, 97%, and 94% being obtained with cationic starch (DS 0.5), chitosan, and pDADMAC at 0.005, 0.050, and 0.020 g/g algae AFDW dosages, respectively. For cationic starch (DS 0.5), flocculation efficiency remained ∼90% in the dosage range of 0.005 to 0.120 g/g algae AFDW. Further increase in the polymer dosage reduced the flocculation efficiency (Fig. 6a). A similar trend was observed for pDADMAC where flocculation efficiency was reduced at doses above 0.100 g pDADMAC/g algae AFDW (Fig. 6b). Flocculation with cationic starch (DS 0.2) had the lowest flocculation performance with maximum flocculation efficiency of 51% being observed at the highest polymer dose of 0.2 g/g algae AFDW.
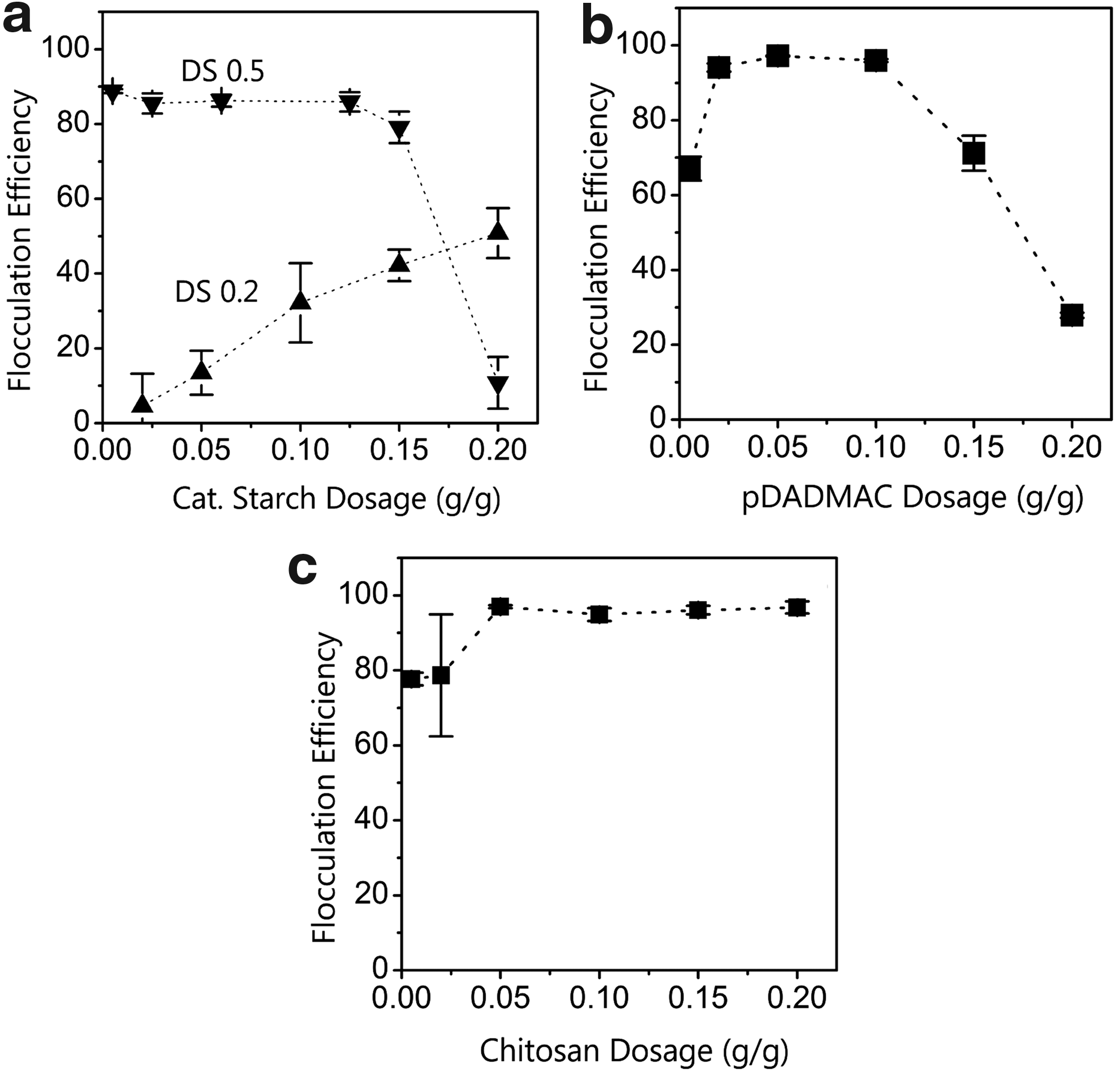
Effect of dosage on flocculation of N. salina at pH 8.0. (Initial cell concentration of 0.10 g/L AFDW).
The energy of repulsion between two spherical particles in a dilute system can be estimated using the Derjaguin approximation
where a1 and a2 are radii of particles 1 and 2, respectively, nα is bulk ion density, k is the Boltzmann constant, T is absolute temperature, γ is the surface potential, and h is the interparticle distance (Liang et al., 2007). Inspection of Equation (3) shows that repulsive energy is proportional to particle surface potentials (γ) and ionic strength (inversely proportional to κ). Thus, occurrence of effective flocculation following polymer treatments (Fig. 6a, c) implies a reduction in the surface potential and subsequent reduction in VR. In addition, N. salina control and cells treated with cationic starch (DS 0.2), zeta potentials at all salinities (0.05, 0.25, and 0.5 g/L), and pH 6.0 and 8.0 were all negative with no significant difference with respect to variable salinity. In contrast, Figure 3b and c shows strongly positive zeta potentials in all cases when N. salina was treated with cationic starch (DS 0.5) and pDADMAC. These zeta potential flocculation studies suggest that particle destabilization is predominantly a charge neutralization mechanism. In contrast with cationic starch and pDADMAC, flocculation efficiency did not decrease with chitosan doses above 0.050 g/g (Fig. 6c). Chitosan precipitated at pH 8.0, resulting in N. salina removal by sweep flocculation, thus addition of more chitosan did not have any impact on flocculation. Similarly, Morales et al. (1985) observed marine algae floc precipitation with chitosan at pH 8.0.
Effect of pH on algae flocculation using chitosan
Of the cationic polymers studied, only chitosan was sensitive to pH where the zeta potential magnitude was reduced at acidic pH compared with basic pH (Fig. 1b), indicating a pH effect on suspension stability and flocculation efficiency. Therefore, it was deemed important to investigate the effect of pH on chitosan flocculation with both C. protothecoides and N. salina. Flocculation efficiencies of C. protothecoides and N. salina at a constant chitosan dosage of 0.075 g/g algae DW and initial cell concentration of 0.1 g/L are shown in Figure 7a and b, respectively. Maximum C. protothecoides flocculation efficiency of 64% occurred at pH 6.5, with modest reductions in the range of 45–55% at pH 4.0–6.0, and a significant (p<0.001) reduction to 20% at pH 7.8 (Fig. 7a). Low flocculation efficiencies may be expected at high pH where negative zeta potentials showed the greatest magnitude (>−10 mV) for C. protothecoides–chitosan aggregates (Fig. 1b). At pH 6.5 and below, the zeta potentials at the chitosan dosage of 0.075 g/g DW were less negative than −5 mV (Fig. 1b), resulting in better separations.

Effect of pH on chitosan flocculation at 0.075 g/g dosage and initial cell concentration of 0.1 g/L.
Similarly, the effect of pH is prominent in N. salina flocculation with chitosan at 15 g/L salinity (Fig. 7b). The flocculation efficiency dropped from 75% to 25% when pH was increased from 6.0 to 7.0. However, further increase in pH showed an increase in N. salina flocculation. Flocculation efficiencies of 93% and 85% were achieved at pH 8.0 and 9.0, respectively, due to chitosan precipitation. With proper pH control, chitosan can be an effective coagulant for both freshwater and brackish-water algae. The results are in agreement with studies conducted by Xu et al. (2013) and Morales et al. (1985) showing that chitosan's cationic activity is increased in acidic pH, and hence pH below 7.0 is desirable for flocculation with freshwater algae species, whereas precipitation at basic pH is ideal for flocculation of marine species.
Conclusions
Cationic starch (DS 0.5) was the most effective coagulant for both C. protothecoides and N. salina separation. Compared with chitosan and pDADMAC, the cationic starch (DS 0.5) dose required for maximum flocculation was significantly lower. Cationic starch (DS 0.2) required very high doses and was ineffective. The difference in the flocculation performance of two cationic starches (DS 0.5, 0.2) demonstrated that flocculation efficiency was proportional to the degree of substituted cationic groups on the polymer. The starches used in this study are widely used and designed specifically for the production of paper, indicating their immediate availability for industrial applications and the potential for enhanced efficiency through optimization of the starch properties for use in algal flocculation processes. Chitosan flocculation was pH dependent with a higher cationic activity in acidic pH. However, efficient flocculation of brackish alga N. salina was achieved at pH 8.0 through chitosan precipitation and subsequent sweep flocculation. Zeta potential measurements demonstrated the charge neutralizing ability of cationic polymers and provided insight regarding coagulant effectiveness and mechanisms. Relatively high flocculation efficiencies were achieved when algal surface charge was sufficiently reduced and complete charge neutralization was not required.
Footnotes
Acknowledgments
The authors would like to acknowledge their funding agency, National Alliance for Advanced Biofuels and Bioproducts (NAABB). The authors thank Dr. Andreescu and Mr. Alkasir (Clarkson University, NY) for providing help with zeta potential measurements.
Author Disclosure Statement
No competing financial interests exist.