
Abstract
Abstract
A formaldehyde-degrading bacterium named BZ-001H was isolated from activated sludge while searching for an efficient strain of bacteria for bioaugmentation during treatment of high-concentration formaldehyde wastewater. It was identified as Bacillus sp. by cell and colony morphologies, biochemical properties, and 16S rDNA sequence analysis. Further study showed that BZ-001H retained its activity with respect to formaldehyde degradation when the pH range was 5–10, temperature range was 5–45°C, and initial formaldehyde concentration was 4,000–20,000 mg/L. BZ-001H exhibited highest removal efficiency and growth rate at pH 7, temperature of 30°C, and 8,000 mg/L of formaldehyde. Moreover, formic acid and methanol were intermediate products. In addition, formaldehyde removal rate was slightly effected by initial substrate concentration, but it was decreased with impact of formaldehyde concentration and intermediate products in the early 9 h. This bacterium might reveal potential with regard to biological formaldehyde treatment in real formaldehyde wastewater due to its strong tolerance to various physicochemical conditions. Overall, this article enriched the study of the biodegradation of high concentration of toxic and refractory organic substrate.
Introduction
F
In general, the formaldehyde in wastewater can be removed by physical, chemical, or biological methods. Compared with biological treatment, the application of physicochemical treatments to remove formaldehyde is often limited by its high treatment cost, inconvenience, and the production of secondary pollutants. However, great performance of formaldehyde biodegradation is difficult to achieve because of the toxicity of formaldehyde at a high concentration on microorganisms and the weak resistance on environmental factors of microorganisms in biological treatment facilities (Vidala et al., 1990; Oliveira et al., 2004; Raja et al., 2009). Some microorganisms capable of degrading formaldehyde have been isolated (Glancer et al., 2001; Yamazaki et al., 2001; Ayako et al., 2005; Mitsui et al., 2005; Kaszycki et al., 2006). However, few of them can tolerate the high concentration of formaldehyde and most have low degradation efficiency. Therefore, it is essential to isolate microorganisms that can tolerate a high concentration of formaldehyde for efficient biodegradation, thereby improving removal efficiency and reducing operating costs.
In this study, a bacterium capable of biodegrading formaldehyde at a high concentration (20,000 mg/L) was isolated. The degradation characteristics and the pathway involved were subsequently investigated. The overall goal of their work and specific objective were to help in providing scientific reference for subsequent application of this bacterium on bioaugmentation of high-load formaldehyde wastewater treatment.
Experimental Protocols
Activated sludge and medium
Activated sludge used for the isolation of a formaldehyde-degrading bacterium was obtained from a suspension aerobic tank at a municipal wastewater treatment plant, using an Anaerobic-Anoxic-Oxic (A/A/O) process in Chengdu, China. The amount of industrial wastewater containing formaldehyde that was injected into this facility was not significant.
Modified synthetic medium was composed of basal medium [0.3 g/L Na2HPO4, 0.5 g/L KH2PO4, 0.5 g/L (NH4)2SO4, 0.61 g/L MgSO4] supplemented with 5 mL/L trace element solution [0.5 g/L EDTA, 0.02 g/L ZnSO4·7H2O, 0.055 g/L CaCl2, 0.051 g/L MnCl2·4H2O, 0.049 g/L FeSO4·7H2O, 0.011 g/L (NH4)2Mo7O24·4H2O, 0.0157 g/L CuSO4·5H2O, 0.016 g/L CoCl2·6H2O, with the pH adjusted to 6.0 using KOH] and a carbon source. Formaldehyde was the sole carbon source and the concentration varied according to the experimental setup (Table 2).
LB medium (Luria Bertani medium): 5 g/L tryptone, 2.5 g/L yeast extract.
Isolation and domestication
Activated sludge (10 mL) was inoculated into 90 mL sterile water. The sample was shaken vigorously for 20 min and then kept static for 30 s. A 1-mL volume of bacterial suspension was removed and inoculated into 100 mL of MS medium containing 500 mg/L of formaldehyde. The mixture was incubated at 30°C with shaking at 160 rpm for 24 h. The culture was diluted, streaked on the modified synthetic agar plate, and then incubated at 30°C for 48 h. The isolate grown on the agar plate was inoculated into modified synthetic media containing formaldehyde, with an increasing concentration from 500 to 12,000 mg/L. The strain was cultivated at 160 rpm and 30°C. Samples were taken every 8 h for the measurement of formaldehyde concentration and Chemical Oxygen Demand (COD) level. The formaldehyde concentration was increased once the COD removal efficiency reached 90%.
Isolate identification
To investigate the morphological and phylogenetic characteristics of the bacterial isolate, scanning microscopy, gram stain, polymerase chain reaction (PCR) and 16S rDNA sequencing were undertaken. The total DNA was extracted from cells, and purification of PCR amplicons was conducted using kits from TIANGEN Biotech (Beijing) Co., Ltd. Extracted DNA was amplified by PCR using general bacterial primers (27F: 5′-AGA GTT TGA TCC ATG GCT CAG-3′ and 1495R: 5′-AAG GAG GTG ATC CAG CCG CA-3′), and amplicons were visualized by 1% (w/v) agarose gel electrophoresis. 16S rDNA sequencing was completed with assistance from Beijing SunBiotech Co., Ltd. Phylogenetic analysis was conducted using NCBI's BLAST software.
Optimal degradation conditions
A number of conditions were examined (Table 2). A suspension of the isolate, designated BZ-001H, was inoculated (0.03% dry cell weight) into 200 mL of modified synthetic medium and incubated at 30°C (except the situation where temperature was the variable factor) and 160 rpm under thermostatic and aerobic conditions. The pH of the medium varied during the culturing. Therefore, the pH of the degradation system was maintained by adding appropriate aliquots of 10% NaOH solution every 1 h. Samples from these batch tests were taken regularly for the measurement of formaldehyde concentration and COD.
Gas chromatography–mass spectrometry analysis and determination of enzyme activity
To investigate the degradation pathway of the isolated strain, gas chromatography–mass spectrometry (GC-MS) analysis was performed and attempted to determine the metabolic intermediates of formaldehyde biodegradation. The isolate was inoculated into 200 mL of modified synthetic medium containing 5,000 mg/L formaldehyde and into 200 mL of LB medium. Cultures were incubated at 160 rpm and 30°C. Aliquots were removed from the modified synthetic medium cultures on a regular basis to determine the concentration of formaldehyde and COD. Samples collected after 2 h were subjected to GC-MS analysis using a GCMS-HP6890/5973 and capillary chromatographic column (HP-INNOWAX, 0.25-mm internal diameter, 0.25-μm membrane thickness, 30 μm length) with pure helium as a carrier gas and a column temperature of 40°C. The temperature increased to 120°C at a rate of 10°C/min and was maintained for 5 min. The injection port was kept at 240°C, with a split ratio of 10 and a column flow rate of 0.8 mL/min. Samples from the modified synthetic medium and LB medium after 4 h were used for the determination of formaldehyde dehydrogenase activity. Formaldehyde dismutase activity was assayed with a standard reaction mixture containing 20 mM formaldehyde, 100 mM KCl, and the enzyme in a final volume of 10 mL. The reaction was carried out at 30°C with stirring under N2 gas, and formate formation was followed by means of pH-stat titration (TOA pH-stat model HSM-10A) with 10 mM NaOH at pH 7.0. One unit of enzyme was defined as the amount of enzyme that catalyzed the formation of 1 μmol formic acid/min (Kato et al., 1986). The formaldehyde dehydrogenase activity was assayed spectrophotometrically at room temperature according to the method described by Kaulfers and Marquardt, 1991. The reaction mixture (2 mL) contained 120 μmol Tris-HC1 buffer, pH 8.0, 5 μmol NAD, and 10 μmol reduced glutathione and cell extract. The reaction was started by the addition of 20 μmol formaldehyde. The reduction rate of NAD was detected at 340 nm in 1-cm cuvettes.
Analytical methods and equipment
Gram staining, spore staining, and utilization of carbon source were determined in accordance with previous reports (Juni, 1984; Dong and Cai, 2001). COD was determined using the dichromate method (National Environmental Protection Bureau, 2002). The optical density at 600 nm (OD600) of bacterial suspensions was measured using a spectrophotometer to investigate BZ-001H growth. The concentration of formaldehyde was determined using the Hantzsch method (Nash, 1953). pH was determined using a pH meter. Protein content was determined using Bradford Colorimetry (Bradford, 1976).
Monod's noninhibitory kinetics equation [Eq. (1)] was used to fit the half-saturation coefficient (Ks) in 3.3 (optimal degradation conditions) (Alireza and Farzaneh, 2013c). Where μ was the specific removal rate of formaldehyde, mg/(L·h); μmax was the maximum removal rate of formaldehyde, mg/(L·h); S was the specific concentration of substrate, mg/L; and Ks was the half-saturation coefficient, mg/L. Calculations and statistical analyses were conducted with Microsoft Excel 2007.
Equipments used in this study included a UV-visible spectrophotometer (TU-1901; PERSEE), a high-speed freezing centrifuge (5430; Eppendorf), an upright incubator with a shaker and temperature controller (IS-RDV1; Crystal), and a pH meter (SG2; Mettler-Toledo).
Results
Isolation and domestication
Following cultivation in the modified synthetic medium, the activated sludge was capable of utilizing formaldehyde as the sole carbon and energy source for growth. From this activated sludge, a formaldehyde-degrading bacterium named BZ-001H was isolated. Subsequently, domestication of BZ-001H by increasing concentration of formaldehyde in the modified synthetic medium was completed, and the performance of COD removal was shown in Figure 1. The adaptation of BZ-001H required 70 cycles. Effluent COD increased from 1.3 to 1,073.21 mg/L as the influent concentration of formaldehyde increased from 500 to 12,000 mg/L. The COD removal efficiency during adaptation remained above 84.07%. An isolate that was capable of degrading formaldehyde at a high concentration was eventually obtained.

COD removal in domestication of BZ-001H with increasing influent concentration of formaldehyde. COD, Chemical Oxygen Demand.
Identification of strain
BZ-001H isolate was able to grow on the plate of solid LB medium, producing an off-whit and opaque colonies with a coarse surface with the expanded edge and a diameter of 3–5 mm after cultivation for 24 h. The isolate was a gram-positive bacterium and its spores were 1–1.3×3–5 μm, with smooth ends (Fig. 2). Amplification by PCR yielded a 1543-bp amplicon. Sequencing analysis revealed that the isolate BZ-001H was a Bacillus species (GenBank Accession No. KP769781).
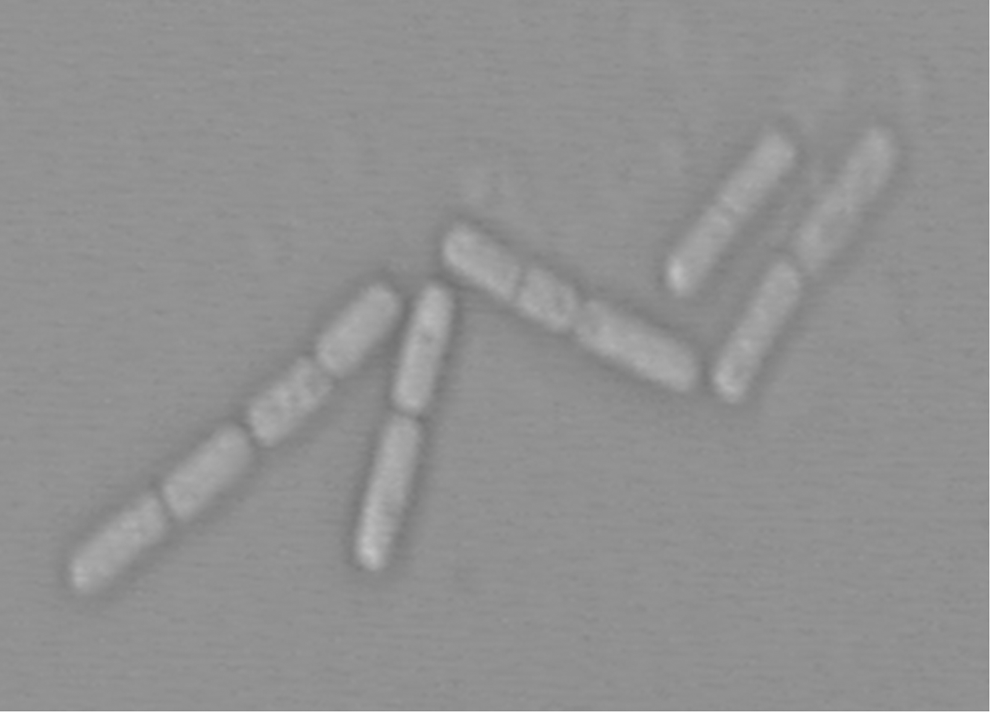
Scanning electron micrograph of strain BZ-001H.
Optimal degradation conditions
BZ-001H was highly resistant to acidic (pH 5) and alkaline (pH 10) conditions (Fig. 3A). Its ability to remove formaldehyde was optimal at a pH range of 6–9, with removal efficiency above 90% and 78% when the formaldehyde concentration in the medium was 4,000 and 8,000 mg/L, respectively. In addition, BZ-001H was more sensitive to pH when the formaldehyde concentration was high, possibly because of the double stress caused by extreme pH and high formaldehyde concentration. BZ-001H at an unaccommodated pH must deal with another stress shock and exhibited a reduced ability to degrade formaldehyde.
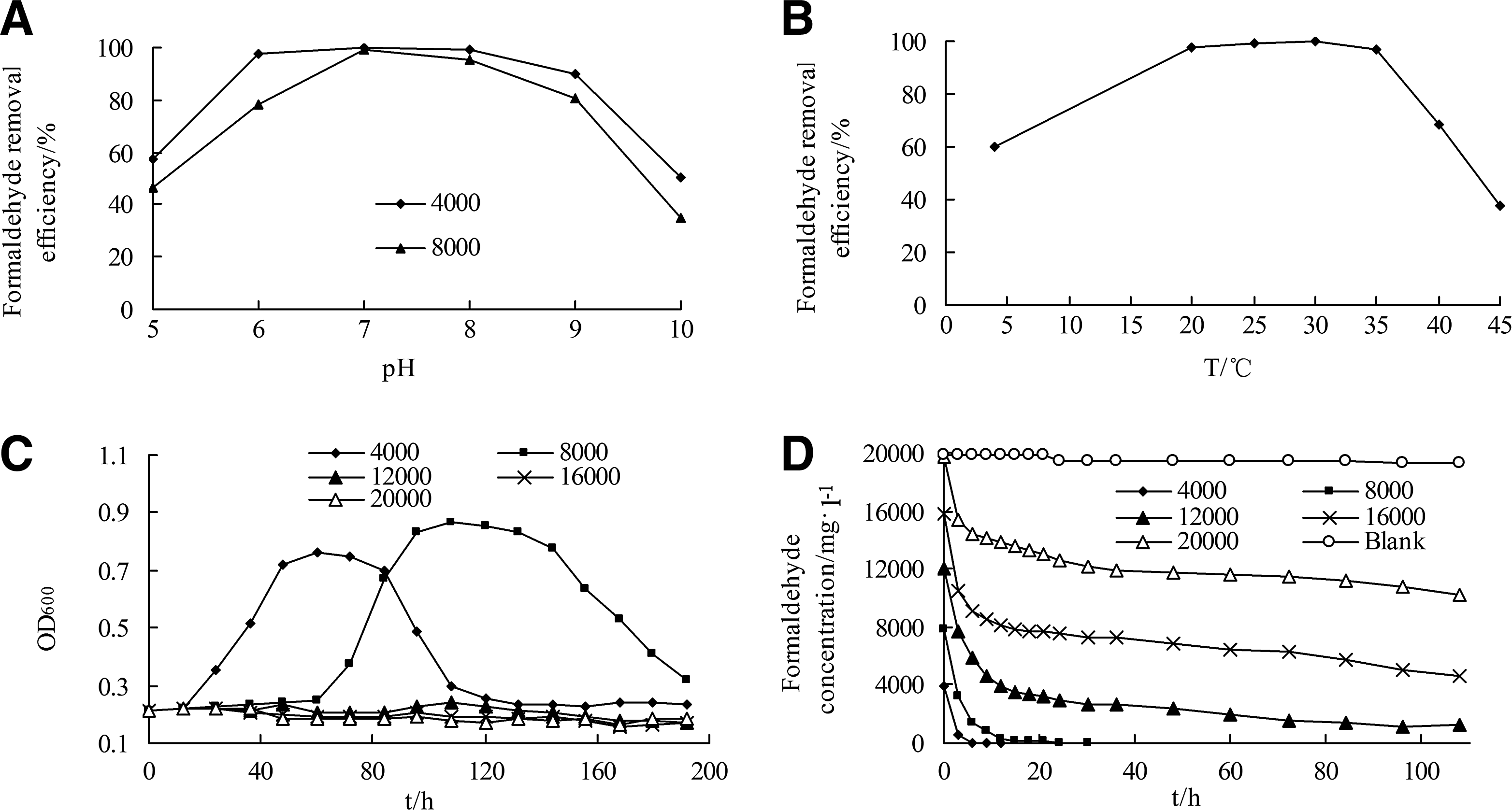
Formaldehyde removal and growth of BZ-001 under different conditions:
Formaldehyde at 4,000 and 8,000 mg/L was completely degraded by BZ-001H within 6 and 14 h, respectively. The maximum growth rate of BZ-001H during the exponential growth phase was 0.014 and 0.024 per hour in the medium containing 4,000 and 8,000 mg/L of formaldehyde, respectively. The BZ-001H isolate was able to degrade 20,000 mg/L formaldehyde; however, the rate was slow, and only 48.15% removal efficiency was achieved. The growth of BZ-001H could not be determined, even after 108 h.
Formaldehyde between 4,000 and 20,000 mg/L was rapidly degraded from 0 to 3 h (Fig. 3D). Yet, After 6 h, degradation proceeded slowly when the concentration of formaldehyde was greater than 12,000 mg/L. Linear regression analysis was conducted based on formaldehyde degradation data of from 6 to 15 h (Table 3). The absolute value of k parameter was the formaldehyde removal rate within the examination period.
μ was the formaldehyde removal rate in 3 h; k and R2 were the slope and regression coefficients in the linear regression equation based on the data of formaldehyde degradation in 6–15 h at the substrate concentration range of 12,000–20,000 mg/L.
A high concentration of formaldehyde did not considerably affect the formaldehyde removal rate during the early 9 h of degradation (Table 3). Increasing formaldehyde concentration stimulated substrate degradation; however, at a certain concentration, the process would be inhibited. The highest removal rate was 1,754.67 mg/(L·h), which was achieved when the initial formaldehyde concentration was 16,000 mg/L. From 6 to 15 h, the removal rate decreased as formaldehyde concentration increased. The removal rate was 253.00 mg/(L·h) when the formaldehyde concentration was 12,000 mg/L, which was 2.90-fold higher than that at 20,000 mg/L. This indicated that the removal rate of formaldehyde by BZ-001H after 9 h was both affected by the initial substrate concentration and the accumulation of intermediate products. In addition, substrate inhibition did not occur when the concentration of formaldehyde was less than 16,000 mg/L. Therefore, the Monod model was suitable. The value of the half-saturation coefficient was 787.79 mg/L, which was much higher than that observed for Ralstonia eutropha (47.6 mg/L). The maximum allowable formaldehyde concentration for R. eutropha was 900 mg/L, indicating a greater resistance by BZ-001H at high formaldehyde concentrations (Alireza and Farzaneh, 2013c).
Despite the highest removal rate during the early 9 h when the initial formaldehyde concentration was 16,000 mg/L, the complete removal of formaldehyde was difficult to accomplish. A removal efficiency of 90.01% was observed at 108 h; however, there was a lag in bacterial growth. Hence, the optimal concentration of formaldehyde enabling its degradation and bacterial growth was determined as 8,000 mg/L, which was obviously higher than that previously reported for other formaldehyde-degrading strains (Mirdamadi et al., 2005; Alireza and Farzaneh, 2013c; Tanida et al., 2013).
Intermediate products and key enzyme activity
Growth of BZ-001H and its ability to degrade formaldehyde were optimal at pH 7.0 and 30°C, when the initial concentration of formaldehyde was 5,000 mg/L (Fig. 4). The maximum formaldehyde degradation and COD reduction rate were 1,005.65 mg/(L·h) within 0–4 h and 203.59 mg/(L·h) within 36–60 h, respectively. These parameters did not occur simultaneously, implying that many intermediate products were produced during the degradation of formaldehyde. Such a large disparity between formaldehyde degradation and COD removal rate suggested that the formaldehyde degradation ability of BZ-001H was greater compared with the intermediate products.
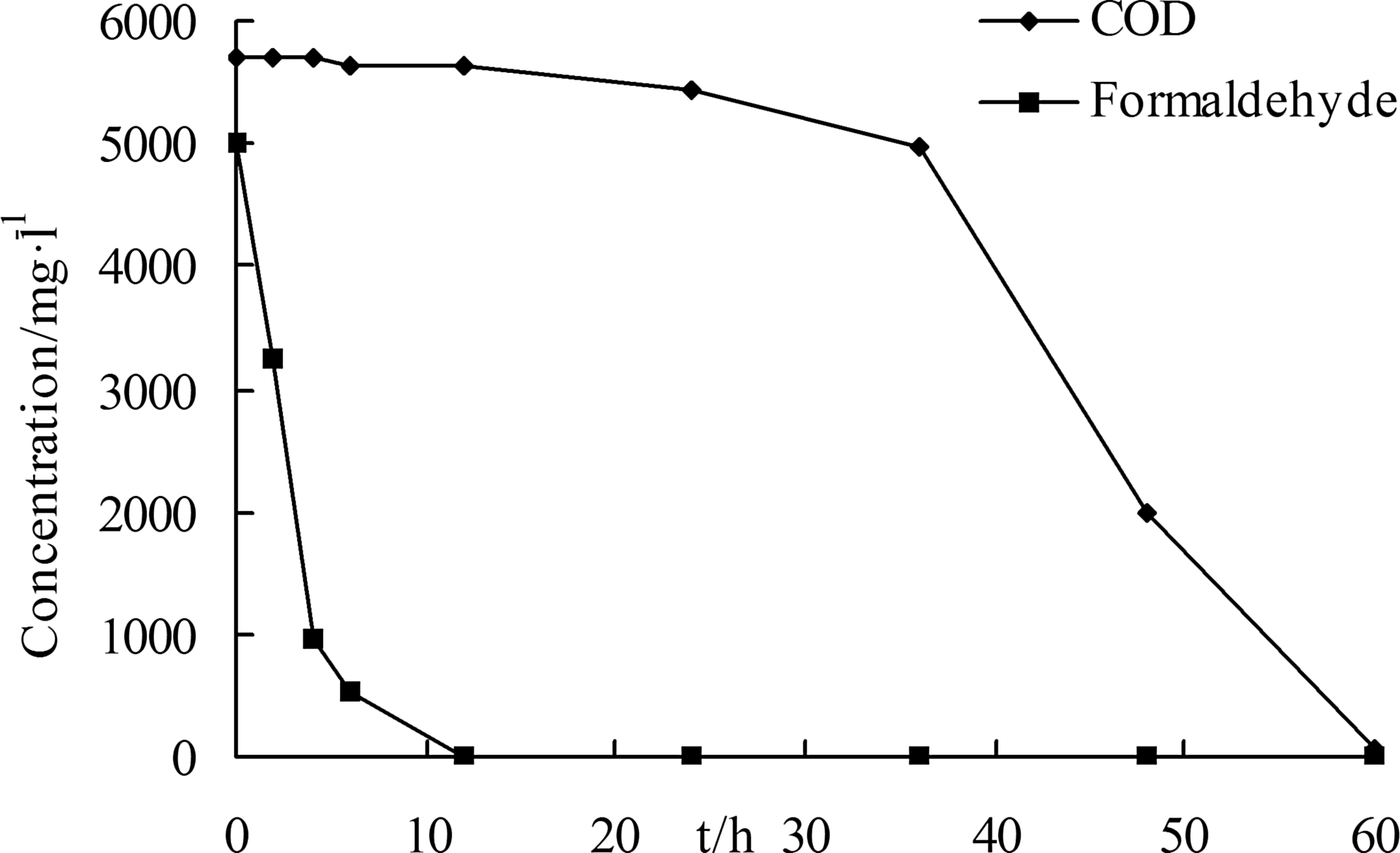
Degradation of formaldehyde and COD by strain BZ-001H.
GC-MS analysis of samples collected after 2 h was used to investigate the intermediate products that appeared during formaldehyde degradation. The three components identified in the reaction system were formaldehyde, methanol, and formic acid, whose retention times were 1.35, 2.56, and 7.07 min, respectively. These compounds comprised 12.37%, 61.59%, and 26.04% of system components, respectively (Fig. 5), and revealed that methanol and formic acid were intermediate products, which were concordant with the results in the research by Marta et al. (2006) that 1,360 mg/L of formaldehyde could degrade under the anoxic condition. In addition, the ratio of methanol to formic acid was 2.37 higher than the theoretical value of 2, which was consistent with previous reports stating that formic acid was utilized first (Adroer et al., 1990).

GC-MS analysis of the formaldehyde degradation products by BZ-001H. GC-MS, gas chromatography–mass spectrometry.
It has been reported that formaldehyde dehydrogenase plays an important role in formaldehyde resistance (Paul-Michael and Annette, 1991). Enzyme activity results revealed that the formaldehyde dehydrogenase activity of BZ-001H in the LB medium was 0.0645 U/mg protein, while that in modified synthetic medium containing 5,000 mg/L of formaldehyde was 1.4 U/mg protein.
Discussion
Microorganisms are at the core of biological wastewater treatment. The complexity and instability of wastewater are widely recognized and are a stressful environment for microorganisms. Consequently, the ability of microbes to remove toxic organic compounds from wastewater is reduced. Thus, microorganisms that exhibit high levels of resistance to toxic compounds could be advantageously employed in biological wastewater treatment.
Generally, strains that are highly resistant to formaldehyde are enriched in mixed cultures after domestication (Whyte et al., 1997; Tian et al., 2012; Alireza and Farzaneh, 2013c), with stepwise adaptation rarely reported. Our results indicate that the resultant resistance of the isolated strain could be promoted by increasing the stress in culture. In addition, Pseudomonas sp. are generally regarded as the model microorganisms with respect to the degradation of organic compounds (Jisun and Woojun, 2014). However, it has also been shown that Bacillus sp. also played an extremely important role in the biodegradation of organic compounds in the natural environment (Manabu et al., 2005; Das et al., 2007; Chang et al., 2008; Fuchedzhieva et al., 2008).
Luis et al. showed that the expression of the growth-related gene was negatively accommodated along with the expression of the stress-related gene. The stress-related gene expression increases the level of self-protection against injury because microorganisms face environmental stress, while the expression of the growth-related gene decreases accordingly. This resulted in a low efficiency of growth and substrate degradation (Luis et al., 2008). The results showed that formaldehyde removal efficiency of 100% and 99.54% was achieved when the initial formaldehyde concentration was 4,000 and 8,000 mg/L, respectively, at an optimum pH of 7. This pH value is the same as the optimal value reported for the formaldehyde-degrading bacteria, SGFA1 and FD3; however, the adapted pH range was broader (Haoyu et al., 2013; Diansi et al., 2014).
Recently, many formaldehyde-degrading bacteria have been isolated and investigated with respect to their performance of formaldehyde removal (Alireza and Farzaneh, 2013c; Haoyu et al., 2013). In previous studies examining degradation conditions, the optimal initial formaldehyde concentration was usually used (Alireza and Farzaneh, 2013b; Tanida et al., 2013). Some bacteria can tolerate high concentrations of formaldehyde; in particular, Paracoccus sp. is able to degrade 8,000 mg/L formaldehyde (Haoyu et al., 2013).
However, other parameters during formaldehyde degradation, such as pH and temperature, have not been widely explored. The BZ-001H isolate exhibited high levels of resistance over a broad pH and temperature range, and for the initial substrate concentration tested. BZ-001H was able to degrade formaldehyde in the pH range 5–10, and a temperature range of 5–45°C, when the initial concentration of formaldehyde was 4,000–20,000 mg/L. The findings revealed that removal efficiency and bacterial growth rate were highest at pH 7 and 30°C in the presence of 8,000 mg/L of formaldehyde. Moreover, BZ-001H could tolerate a formaldehyde concentration approaching 20,000 mg/L, implying that BZ-001H could potentially be applied to the biodegradation of formaldehyde in wastewater.
It has been reported that there were two aerobic degradation pathways for formaldehyde (Margareta et al., 2001). In the first pathway, formic acid is generated through the actions of formaldehyde dehydrogenase and then oxidized into CO2 by formate dehydrogenase. In the second pathway, formic acid and methanol are generated through the actions of formaldehyde dismutase, which are then utilized by microorganisms as carbon sources (Adroer et al., 1990; Crowther et al., 2008). Four characteristics of formaldehyde degradation by BZ-001H could be concluded as below: (1) formaldehyde was rapidly degraded; (2) the removal rate for intermediate products was much lower than that for formaldehyde; (3) formic acid and methanol were intermediate products; (4) formaldehyde dehydrogenase activity could be determined during formaldehyde degradation. Unexpectedly, these four characteristics of formaldehyde degradation were observed for both pathways. These findings indicate that BZ-001H might degrade formaldehyde using both pathways simultaneously. The specifics of the BZ-001H formaldehyde degradation pathway and the relationship between the concentration of substrate and intermediate products require further investigation.
Conclusions
A formaldehyde-degrading bacterium Bacillus sp. BZ-001H was isolated from activated sludge in this study, which was able to tolerate extremely high concentrations of formaldehyde (up to 20,000 mg/L). Subsequently, optimization conditions revealed that this strain was able to degrade formaldehyde over a wide pH and temperature range, and for various initial concentrations of formaldehyde. Then, formic acid and methanol were identified as the intermediate products of formaldehyde degradation by BZ-001H. These intermediates accumulated during the degradation of formaldehyde under aerobic conditions, transforming by BZ-001H into biomass and CO2. The rate of removal for formaldehyde was much slower under these conditions, with formaldehyde dehydrogenase playing an important role in this reaction. In addition, formaldehyde dehydrogenase played an important role in this reaction. Taken together, this study could provide efficient strain and data reference to the bioaugmentation in the treatment of high-concentration formaldehyde wastewater.
Footnotes
Acknowledgments
This work was supported by a Special Project of the Ministry of Science and Technology of China for International Science and Technology Cooperation and Exchanges (No. 2013DFG92600), the National Natural Science Foundation of China (No. 51208489), and the Sichuan Scientific and Technological Support Program (No. 2012SZ0161).
Author Disclosure Statement
No competing financial interests exist.