
Abstract
Abstract
This review provides insights on (1) the chemistry of arsenic (As) in the soil environment and factors (e.g., pH, presence, nature, and concentration of competing inorganic and organic ligands), which regulate the sorption/desorption of arsenate and arsenite (the most abundant As species in soils and waters) on/from soil components; (2) the chemical fractionation and speciation of As to identify species that are more mobile and phytoavailable; and finally (3) the uptake of As by selected edible plants and production techniques able to reduce its translocation in plant tissues.
Introduction
A
In natural environments, As has oxidation states ranging from −3 to +5. The dominant forms are the inorganic species, arsenite [As(III); H3AsO30, H2AsO3−, HAsO32−, AsO33−] and arsenate [As(V); H3AsO40, H2AsO4−, HAsO42−, AsO43−]. Arsenite is more toxic and mobile than As(V). Redox potential (Eh) and pH are the most important factors controlling As speciation. Under oxidizing conditions, H2AsO4− is dominant at low pH (less than pH 6.9), while at higher pH, HAsO42− becomes dominant (H3AsO40 and AsO43− may be present in extremely acidic and alkaline conditions, respectively). Under reducing conditions, at pH less than pH 9.2, the uncharged arsenite species, H3AsO30, usually predominates (Frankenberger, 2002; Smedley and Kinniburgh, 2002; Violante et al., 2008a). Arsenic might be present as methylated forms in nature, but these organic compounds are usually scarce in soils and water.
The mobility and bioavailability of As in soils are affected by many processes, such as sorption/desorption, precipitation, surface complexation, and oxidation/reduction. Arsenite and As(V) are retained mainly on the sorption sites of variable charge minerals (Fe, Mn, and Al oxides, allophanic materials, and imogolite) and edges of phyllosilicates (Violante et al., 2008a).
Arsenic in natural waters is a worldwide problem. Arsenic pollution has been reported in many countries, such as Argentina, Bangladesh, Cambodia, Chile, China, Thailand, and Vietnam (Smedley and Kinniburgh, 2002; Ahmed et al., 2006). Over the past few years, the presence of As in groundwater has been considered a matter of concern in Italy also, where numerous cases of As contamination in natural waters have been reported (Di Giuseppe et al., 2014; Sappa et al., 2014). The issue of As contamination in aquatic environments, mainly in the Bengal region and Bangladesh, assumes a catastrophic proportion for human health (Sharma et al., 2014).
Uncontaminated waters usually contain less than 0.001 μg As/L. In contaminated areas, however, high levels of As have been reported in water bodies. The WHO imposes a maximum As concentration in drinking water of 10 μg/L (WHO, 2004; USEPA, 2006). However, in some countries, such as India, Bangladesh, Taiwan, China, and Vietnam, the maximum contamination level for As in drinking water is still 50 μg/L (Mondal et al., 2013).
Human beings may be exposed to As from air, food, and water, with dietary sources being largely predominant (Adriano, 2001; Violante et al., 2008a). Prolonged As ingestion by drinking water or food may lead to typical arsenical lesions, which are chronic symptoms of As toxicity (USDHHS, 2000). In addition, in plants, As hampers the metabolic processes, for example, photosynthesis, which determine stunted growth and can lead to death (Tu and Ma, 2002).
The present review aims to provide information on (1) the factors affecting the sorption/desorption processes of As on/from soil components and soils; (2) the chemical fractionation and speciation of As in soil to identify the species more mobile and phytoavailable; and finally (3) the As uptake by plants and possible techniques of soil As compound biostabilization.
Sorption of Arsenic onto Soil Components
In soil and related environments, the transformation and mobility of As is governed by abiotic and biotic processes (Huang and Gobran, 2005; Violante et al., 2008a). Abiotic processes include sorption, desorption, precipitation, dissolution, and redox reactions.
The soil components, which are able to sorb ions in soil, include phyllosilicates, allophanes, imogolite, carbonates, Fe, Al, and Mn oxides, organic matter (OM), and microorganisms (Violante, 2013). Soil components differ greatly in their sorption capacities, their cation and anion exchange capacities, and the binding energies of their sorption sites. Soil components usually are not present as separate entities, but interact each other forming stable complexes. The mutual interactions among phyllosilicates, metal oxides, humic substances, and/or microorganisms promote the formation of complexes with peculiar sorption properties and significantly affect the sorption of ions (Violante and Gianfreda, 2000; Violante, 2013).
Spectroscopic techniques, for example, electon spin resonance and extended x-ray adsorption fine structure (EXAFS), provide useful information on As complexes at the surfaces of soil components (Violante, 2013). When As species react with a surface functional group of a soil component, a surface complex is formed. There are two types of surfaces complexes, inner sphere and outer sphere. An inner-sphere surface complex has no water molecules interposed between the surface functional group and the ion it binds, whereas an outer-sphere surface complex has at least one interposed water molecule (Sparks, 2003). Ions forming inner-sphere complexes on the surface of a sorbent are said to be specifically sorbed, indicating that they are strongly held through bonds with a covalent character. Vice versa, ions retained by the outer-sphere mechanism are said to be nonspecifically sorbed.
Arsenate and As(III) may form different surface complexes on inorganic soil components: monodentate, bidentate–binuclear, and bidentate–mononuclear complexes in different proportions depending on pH and surface coverage (Violante and Pigna, 2002; Violante et al., 2008a; Violante, 2013). Arsenate easily forms inner-sphere complexes on minerals showing a behavior similar to PO4 in soils. Arsenite is not strongly retained by aluminous minerals, but has a strong affinity for the surfaces of minerals containing Fe [mainly (hydr)oxides] (Violante and Pigna, 2002; Violante et al., 2008a). By EXAFS spectroscopy, Ona-Nguema et al. (2005) found that As(III) forms bidentate mononuclear edge sharing and bidentate binuclear corner sharing on ferrihydrite.
As all other anions, As(III) and As(V) sorb primarily to variable charge minerals, carbonates, and at the edges of phyllosilicates (Cornell and Schwertmann, 1996; Kampf et al., 2000). Phyllosilicates have a permanent negative charge due to isomorphous substitution (e.g., Al3+ for Si4+ in the tetrahedral sheet and Mg2+ for Al3+ in the octahedral sheet), which is balanced by cations, but the sites at crystal edges (e.g., protonation of broken Al-OH bonds exposed at particle edges) may easily sorb elements in the anionic form as PO4, As(V), SeO3, and chelating organic ligands. Phyllosilicates exhibit less As(III) sorption than As(V). Figure 1 shows the sorption of As(V) (mmol/kg) on selected phyllosilicates as a function of pH. Greater quantities of As(V) were sorbed on smectite than on the other minerals for the high surface area of this phyllosilicate (up to 800 m2/g).
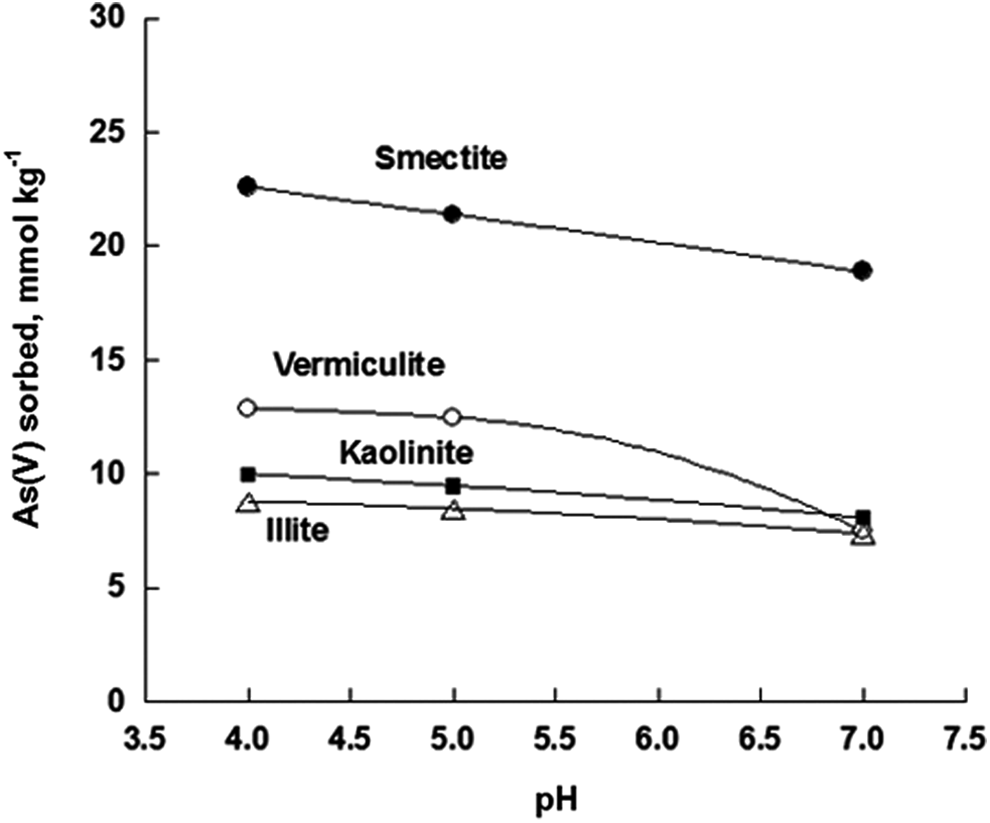
Sorption of arsenate [As(V)] (mmol/kg) on selected phyllosilicates as a function of pH.
Arsenic has been found to be bound to humic substances. Many binding mechanisms were identified for As sorption by OM to comprehend the formation of (1) outer-sphere complexes with protonated amino groups, (2) covalent bonds with carboxylate or phenolate groups, and (3) the formation of ternary complexes with polyvalent cations (Fe, Al) (Mikutta and Kretzschmar, 2011). These authors showed spectroscopic evidences for the formation of a ternary complex between As(V), ferric ions, and humic substances, with at least 70% of As(V) bound to Fe(III)-HS complexes through inner-sphere complexation.
Metal oxides, among which those of Al, Fe, and Mn are ubiquitous in soils, are of great interest in soil chemistry because of their specific adsorptive properties for anions and cations. Al, Fe, and Mn (hydr)oxides may exist as crystalline minerals (e.g., gibbsite, goethite, hematite, birnessite) and as short-range ordered (ferrihydrite) or noncrystalline and nanosize minerals. These minerals often occur as a coat over clay mineral surfaces (Yuan and Theng, 2012). Allophane and imogolite are nanosize and short-range aluminosilicates with sorption properties similar to those of the metal oxides.
The pH strongly affects As sorption on variable charge minerals and soils. Arsenate is sorbed more on Fe than Al oxides (Fig. 2). Arsenite may be sorbed more easily at high pH values because it forms a weak acid at low pH and, consequently, may be dissociated only in alkaline environments (Sparks, 2003). On Fe oxides, particularly ferrihydrite, the sorption capacity of As(III) compares or exceeds that of As(V), the former showing only slight pH dependence in the pH range 4.0–8.0 (Fig. 3). Anyway, As(V) is sorbed in greater amounts and more strongly than As(III) on most soil components. Arsenite is easily oxidized to As(V) when sorbed on the surfaces of Mn oxides. Arsenate is sorbed on calcite, showing a maximum sorption between pH 10 and 12, but few studies have been carried out on the sorption of As on calcite.

Sorption of arsenate [As(V)] (mmol/kg) on ferrihydrite and Al(OH)x as a function of pH. The numbers in parentheses indicate the surface areas (m2/g), which were determined by water adsorption.

Effect of pH on the sorption of arsenite [As(III)] and arsenate [As(V)] on ferrihydrite.
Arsenic mobility is strongly affected by microorganisms (biotic processes), extracellular enzymes, and other biopolymers (Huang and Germida, 2002). Arsenic, free and sorbed on soil colloids, may affect the activity of extracellular enzymes (Gobran et al., 2000) and, consequently, the biological activity of the soil.
Influence of competing anions
Inorganic [e.g., sulfate (SO4), phosphate (PO4)], and organic ligands (humic and fulvic acids and low-molecular-mass organic ligands [LMMOLs]) play a relevant role on As availability in soil environments. LMMOLs are produced by leaching from plants, decomposition of litter by microbes, and exudation by plants and microorganisms. The rhizosphere is enriched with biomolecules of plant and microbial origins that include organic acids, amino acids, lipids, proteins (enzymes), carbohydrates, and other substances. Among them, the organic acids are the most significant as substrates for microbial metabolism (Violante and Gianfreda, 2000). Oxalic, malic, succinic, citric, tartaric, and malonic acids are the most abundant aliphatic acids present in the rhizosphere (Violante and Gianfreda, 2000).
The presence of foreign ligands affects the sorption/desorption of As species on/from soil minerals and soils by competing for available binding sites and/or reducing the surface charge of the sorbents (Frankenberger, 2002; Violante and Pigna, 2002; Violante et al., 2005; Violante, 2013).
The competition in sorption between As(III) or As(V) and other anions is influenced by the affinity of such anions for the nature of sorbent surfaces, the surface coverage, and the reaction time (Zhu et al., 2011, 2013).
The influence of PO4 on the sorption/desorption of As in the soils has been widely studied since the application of PO4 as a fertilizer has an impact on the mobility and availability of As(V) in the soil (Violante and Pigna, 2002; Violante et al., 2008a). Violante and Pigna (2002) investigated the competitive sorption between PO4 and As(V) on metal oxides, phyllosilicates, and different soils. These authors observed that Mn and Fe oxides and Fe-based phyllosilicates (nontronite, ferruginous smectites) were more efficient in sorbing As(V) versus PO4; conversely, more PO4 than As(V) was sorbed on Al oxides (gibbsite, boehmite), allophane, and kaolinite (Table 1). Violante and Pigna (2002) also noted that pH deeply influenced the competition between As(V) and PO4; specifically, at pH <7, As(V) competed more than PO4, whereas the opposite was observed at higher pH values (Table 1).
Reaction time 24 h. Modified from Violante and Pigna (2002). rf stands for sorbed As(V)/PO4 molar ratio.
Violante et al. (2008a) also carried out experiments on the effect of the sequence of anion addition on the competitive sorption of PO4 and As(V) on short-range-ordered Al and Fe oxides (Fig. 4). The anions were added to the samples as follows: contemporary addition (As(V)+PO4 system), addition of As(V) 24 h before PO4 (As(V) before PO4 system), or addition of PO4 24 h before As(V) (PO4 before As(V) system). Figure 4A shows the amounts of As(V) sorbed on Al(OH)x, while Fig. 4B depicts the amounts of As(V) sorbed on ferrihydrite (Fig. 4B) after 24 h of reaction in the absence or presence of PO4. The numbers in brackets refer to the efficiency (%) of PO4 in inhibition of As(V) sorption, calculated according to the expression of Deb and Datta (1967) as follows:
It is clear that PO4 inhibited more efficiently As(V) sorption on Al(OH)x versus ferrihydrite. In fact, PO4 efficiency in inhibiting As(V) sorption on Al(OH)x (in the case of the contemporary addition) was 79% (Fig. 4A), whereas on ferrihydrite, it was much lower at 41% (Fig. 4B). The sequence of anion addition markedly affected As(V) sorption. Minor amounts of As(V) have been sorbed in the PO4 before As(V) system; vice versa, higher amounts of As(V) have been sorbed in the As(V) before PO4 system compared with the As(V)+PO4 system (Fig. 4).
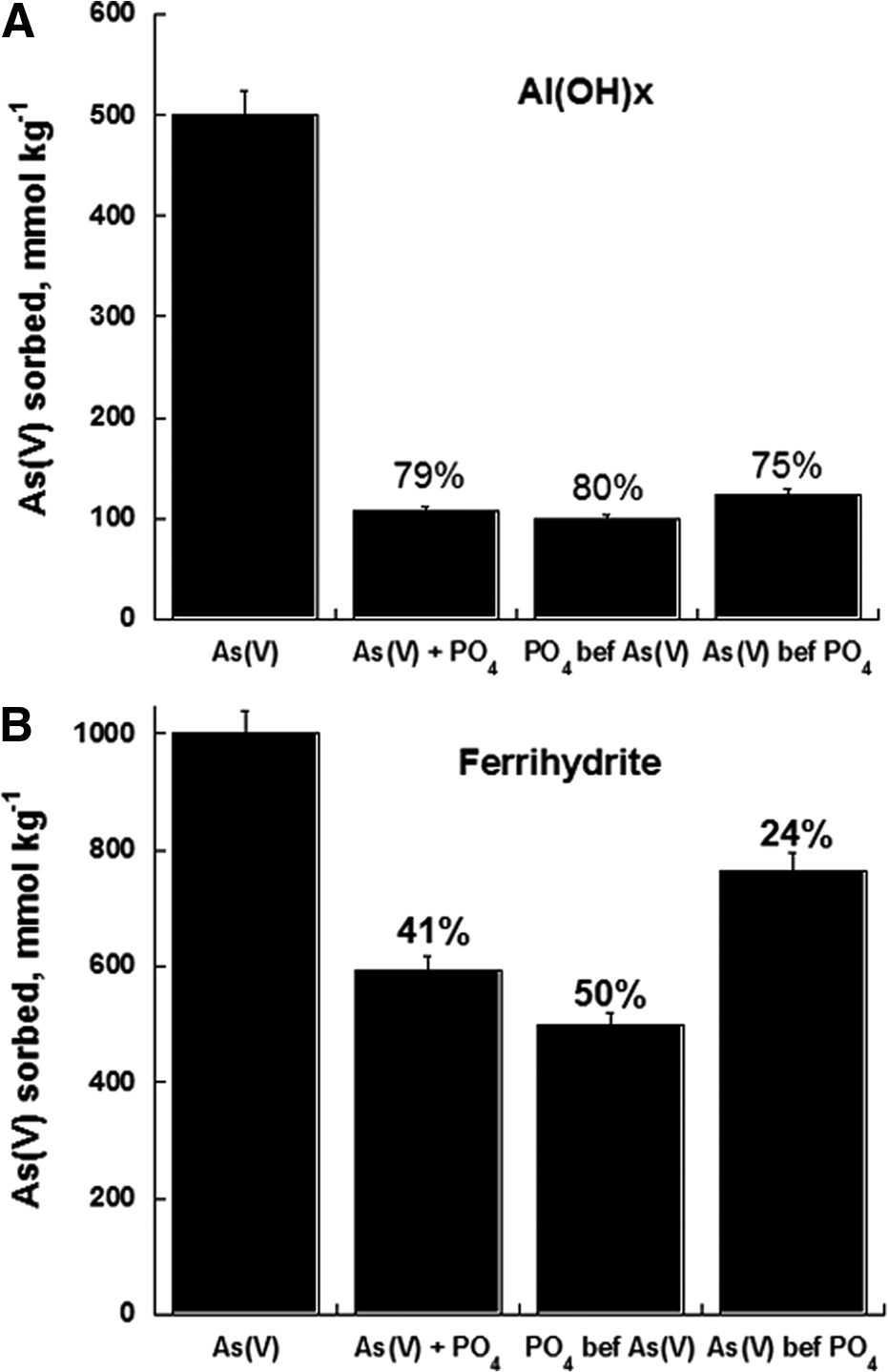
Arsenate [As(V)] sorbed onto Al(OH)x
The influence of other inorganic ligands (sulfate, molybdate, silicate), fulvic and humic acids, and LMMOLs on the sorption of As(III) and As(V) on the soils and minerals was studied (Liu et al., 2001; Zhu et al., 2011, 2013).
Recently, Zhu et al. (2011) studied the effect of different organic [oxalate (OX), malate (MAL)] and inorganic [phosphate (PO4), sulfate (SO4), selenate (SeO4), and selenite (SeO3)] ligands on the adsorption of As(III) and As(V) on ferrihydrite (Fig. 5). These authors demonstrated that the organic and inorganic ligands diverged significantly in preventing As(III) and As(V) fixation onto ferrihydrite. Phosphate and SeO3 were the most efficient, whereas SO4 and SeO4 were the weakest. In fact, at pH 6.0 and an initial As(V)/ligand molar ratio (R) of 1 (Fig. 5A), PO4 prevented As(III) sorption of 48.3% and SeO3 of 22.3%, whereas SO4 and SeO4 had a negligible influence in inhibiting As(III) fixation (<2.5%). The organic ligands had a similar behavior, with malate slightly stronger than oxalate. The trend was similar for As(V) even though the inhibition of its sorption by organic and inorganic anions resulted in lower levels (Fig. 5B).
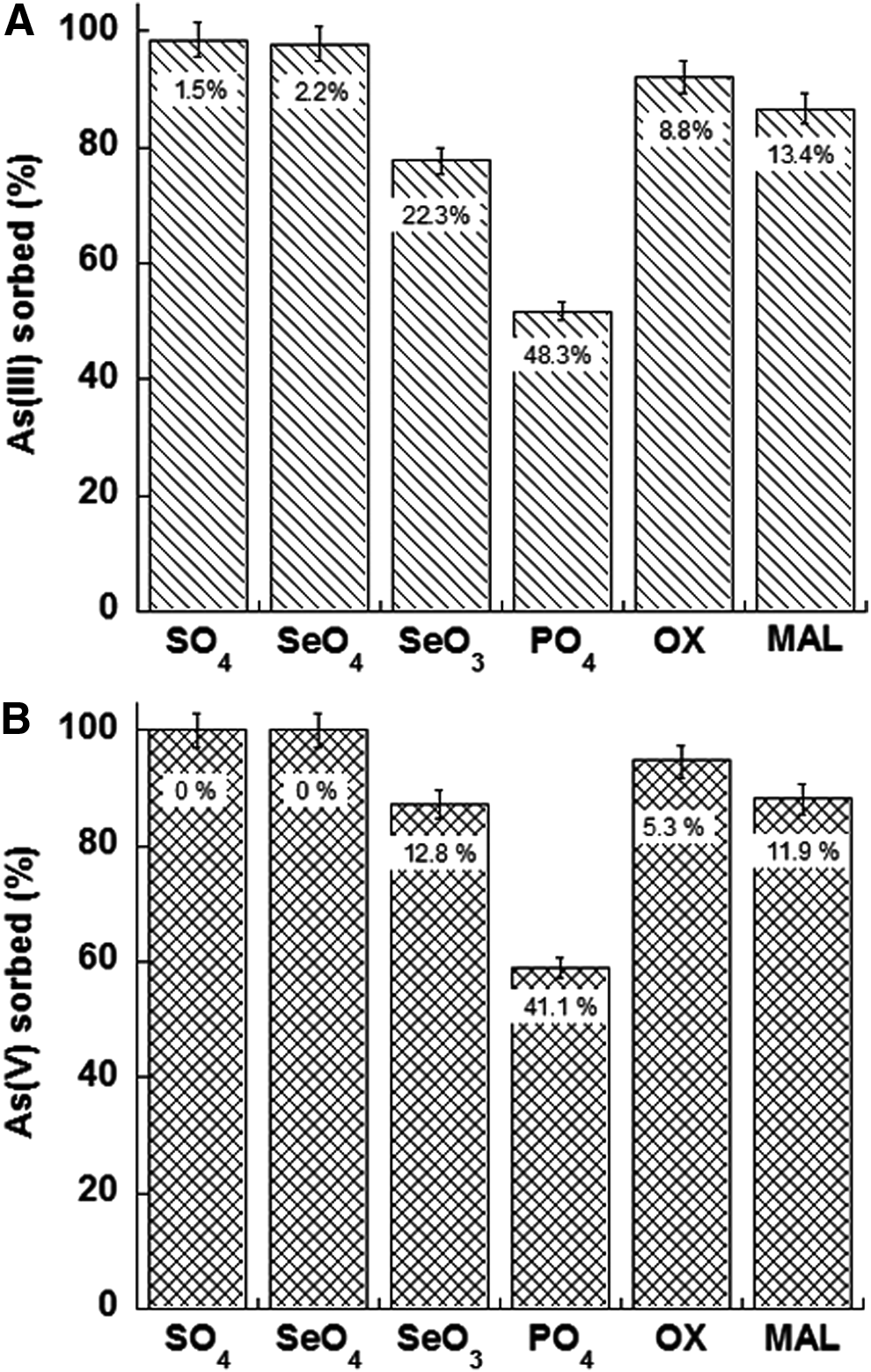
Sorption % of As(III)
Arsenic desorption from minerals and soils
Few scientific works have been done on desorption of As from soils and minerals. Kinetic studies on desorption of As by competing anions may define the As(V) desorption mechanisms toward the soil solution (Frankenberger, 2002; Violante and Pigna, 2002; Pigna et al., 2006; Violante et al., 2009). A number of factors, such as the type, mineralogy, and crystallinity of the sorbents, the pH, the surface coverage, the residence time of As on the surfaces of the sorbents, and the oxidation state of As, could influence the desorption of this metalloid.
Soils enriched in Al, Fe, or Mn oxides, allophane, and imogolite do not desorb As easily. Phosphate was found to be the strongest scavenger in displacing sorbed As from minerals and soils (Arai and Sparks, 2002; Violante and Pigna, 2002; Pigna et al., 2006).
Some literature on the influence of residence time on the sorption/desorption of As has been done. Arai and Sparks (2002) studied the influence of residence time on the desorption of As(V) on bayerite. They noted that a higher residence time corresponded to a decrease in As(V) desorption in the presence of PO4.
Pigna et al. (unpublished data) evaluated the influence of residence time (24 and 168 h) on As(V) desorption by PO4 [PO4/As(V) molar ratio of 3] from a 42% allophane-containing volcanic soil (Fig. 6). The kinetics of desorption of As(V) previously sorbed on Andisol within the first 15 days was studied by the Elovich kinetic model (Fig. 6). The Elovich equation might indicate inconsistency in results commonly unnoticed by other kinetic models. This inconsistency may indicate peculiarity of sorption sites of the sorbents. These breaks in the Elovich plot may suggest a changeover from one type of bonding site to another (Low, 1960). The amounts of As(V) desorbed by PO4, after 2 weeks (ln time=5.88) of reaction, decreased by 53% when PO4 was added 24 h after As(V) and by 22% when PO4 was added 168 h after As(V) (Fig. 6).
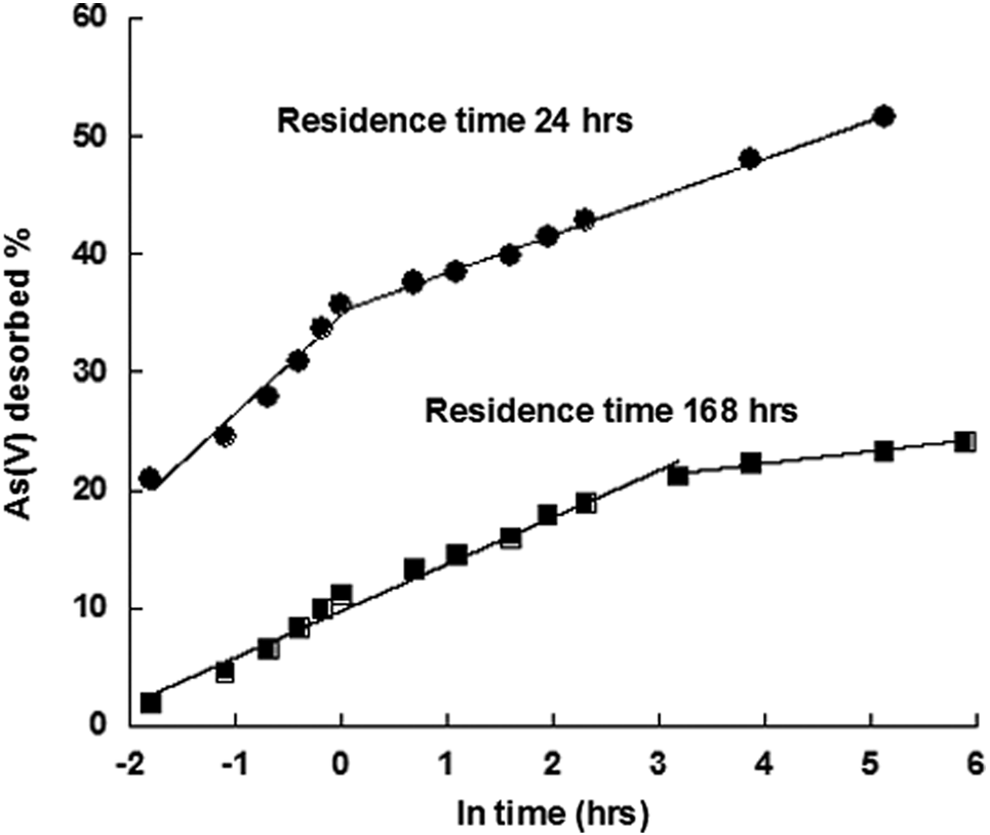
Influence of residence time on the desorption of arsenate [As(V)] by phosphate (PO4) from an Italian Andisol. PO4/As(V) molar ratio=3.
Violante et al. (2006, 2007) noted that PO4 was able to replace a higher amount of As(V) from Al or Fe oxides (surfaces on which the toxicant is simply sorbed) in comparison with oxides obtained by coprecipitating As(V) with Fe and/or Al because, on these latter oxides, As is partially blended into the network of the precipitates.
Chemical Fractionation of Arsenic
The determination of the total content of toxic elements, for example, As, in soil does not provide sufficient information on their bioavailability (Violante et al., 2008b).
In the last few years, some states began to introduce the concept of bioavailability in their laws regarding environmental protection. The bioavailability of toxic elements, and their uptake by plants and soil biota, must be better studied in terms of their chemical speciation. The bioavailability of a heavy metals and metalloids in the soil is related to its concentration in soil solution, the interaction with other soluble species, soil pH, and redox potential (Krishnamurti and Naidu, 2002; Huang and Gobran, 2005; Krishnamurti et al., 2007).
Little research has been done till now on this very important topic. Furthermore, while there are a large number of sequential extraction procedures available for metal cations, only limited work has been done on oxyanions, such as As(III) and As(V) (Wenzel et al., 2001; Krishnamurti and Naidu, 2002). Wenzel et al. (2001) proposed a novel method for As sequential extraction, providing five extraction steps using reagents that are progressively stronger (Table 2). This procedure is reported in Table 2.
Modified from Wenzel et al. (2001).
1, Nonspecifically sorbed; 2, specifically sorbed; 3, sorbed on amorphous and poorly crystalline hydrous Fe and Al oxides; 4, sorbed on well-crystallized hydrous Fe and Al oxides; 5, residual phases.
Cozzolino et al. (2010), by the abovementioned procedure, studied the sequential As extraction in Italian contaminated soil (Vetricella) from Scarlino (Tuscany, Italy) having 250 mg As per kg of soil (Fig. 7). Mostly, As was found in the crystalline oxides (55%, step 4) and short-range Al and Fe oxides (22%, step 3). The most available fractions (steps 1 and 2) resulted in 8.2% of the total; in particular, the first fraction was lower than 0.2%, whereas the second fraction was about 8%. The residual As fraction (14–15%) indicated a significant presence of As-enriched primary minerals.

Arsenic fractionation from an Italian polluted soil (Vetricella, 250 mmol/kgof As) following the Wenzel et al. (2001) method.
Arsenic Uptake by Edible Plants and Production Techniques Able to Minimize the Translocation of As in Plant Tissues
To assess the possible risks arising from the consumption of agricultural products coming from crops grown on As-contaminated soils and/or irrigated with As-contaminated water, further study is necessary to understand the mobility of As in the soil–plant system and, even more, on the possibility to minimize its accumulation in plants of agricultural interest. Among the possible strategies able to reduce the As translocation from the soil to plant, the biological stabilization of soils could play an important role (Pigna et al., 2009; Cozzolino et al., 2010; Pigna et al., 2010, 2012; Violante et al., 2012). In plants, As is accumulated mainly in the roots, to a lesser extent in the aboveground organs, and causes physiological changes and damages (Marin et al., 1992; Cozzolino et al., 2010; Pigna et al., 2010, 2012).
Arsenic causes a reduction of the photosynthesis rate as well as damages the chloroplast membrane and disorganizes the membrane structure (Miteva and Merakchiyska, 2002). The capacity of the plants to survive in polluted soils is affected by many mechanisms of metal tolerance and detoxification, including chelation, compartmentalization, biotransformation, and cellular repair (Gonzaga et al., 2006).
There is evidence that reduction of As(V) to As(III) is a strategy that hyperaccumulator plants use to accumulate large amounts of As without developing toxicity symptoms (Ma et al., 2001). Although As(III) is more phytotoxic than As(V), once reduced, As(III) becomes less toxic within the plant due to the formation of As(III)-thiol complexes (-SH) and As(III) phytochelatins (Schat et al., 2002). The additions of PO4, compost, and arbuscular mycorrhizal fungi (AMF) are procedures that can reduce the translocation of As from the soil to the plant and therefore may encourage the cultivation of As-polluted soils.
Influence of phosphate
Phosphorus (P) and As are chemical analogs, which are taken up by plants though the same transporters (Ullrich-Erebius et al., 1989; Meharg and Macnair, 1992). In the cytoplasm, As(V) competes with PO4, for example, substituting PO4 in ATP to form unstable ADP-As, leading to the breaking of energy flows in cells (Meharg, 1994).
Sensitive plants may be made more tolerant to As(V) by enhancing their P nutrition. It is also due to the stimulation of the activity of As(V) reductase involved in the reduction of As(V) to As(III) and thus vacuolar sequestration as an As(III)-phytochelatin complex (Meharg and Macnair, 1992; Lee et al., 2003). In addition, in As(V)-resistant plants with good P nutrition, a higher As tolerance was found probably due to the higher cytoplasmatic P status, which decreases As(V) toxicity in the cells (Meharg, 1994). The influence of plant P status on As(V) metabolism may be due to the following: (1) high levels of P in the plant cause a downregulation of the As(V)/PO4 plasmalemma transporters and (2) high levels of P in the cell compete more efficiently with As(V) for biochemical processes (Meharg, 2005).
Pigna et al. (2009) studied the As phytotoxicity and accumulation in the grain, shoots, and roots of wheat (Triticum durum) cultivated in natural soil and irrigated with As-contaminated water, with or without PO4 supply. In the plants nonfertilized with P (P−), these scientists noted a decrease in total biomass of 15%, 52%, and 67% in comparison with control treatment as long as As concentration in the irrigation water increased from 0.5 to 1 and 2 mg/L, respectively. This stunted growth was less intense in plants fertilized with PO4 (12% [0.5 mg/L], 16% [1 mg/L], and 26% [2 mg/L]) (Table 3). Similar trends have been observed with rice (Abedin et al., 2002) and wheat (Liu et al., 2005). The higher the As level in the irrigation water, the higher the As concentration in all plant tissues (see table 3 in Pigna et al., 2009).
Modified from Pigna et al. (2009). Data are expressed as mean values±SD (n=4) and have been analyzed by two-way analysis of variance. Means, followed by the same letter within columns, are not significantly different by Tukey's test at the 5% level.
P+ treatments=0.20 g/pot P (as KH2PO4).
In P− plants, As concentration in the grain of control treatment resulted in 0.05 mg/kg, which rose to 0.20 mg/kg by irrigating plants with the 2 mg As/L solution, whereas in P+ plants, the grain As concentration slightly rose from 0.05 (control) to 0.14 mg As/kg (with the highest As level) (see table 3 in Pigna et al., 2009). These findings may suggest that at high concentrations in irrigation water, As causes a severe phytotoxic effect, resulting in reduced growth rate and lower translocation of nutrients (e.g., P) to the grain. Similar trends have been described by Rahman et al. (2007) in rice plants and by Carbonell-Barrachina et al. (1997) in tomato and bean plants.
Influence of AMF
Soil microorganisms are affected by the presence of high concentrations of toxicants in soil (Gadd, 2005); in turn, such organisms are able to influence the availability of toxicants in soil, either directly, through alterations of pH, Eh, biosorption, and uptake, or indirectly in the rhizosphere through their effect on plant growth, root exudation, and resulting rhizosphere chemistry (Leyval and Joner, 2001).
AMF are among the most widespread soil microorganisms and constitute an important functional component of the soil–plant system, occurring in almost all habitats and climates (Cozzolino et al., 2010). Inoculation with AMF has been widely reported to modify toxicant uptake by host plants even though the results are often contradictory (Göhre and Paszkowski, 2006). In general, AMF-inoculated plants are more efficient than those that are noninoculated in the acquisition of micronutrients, such as copper, iron, manganese, and zinc, when available at low concentrations (Vosátka et al., 2006).
However, when grown in excess of micronutrients or in soils contaminated by metalloids or heavy metals with unknown biological function, such as Cd, Cr, or Pb, AMF-inoculated plants can show enhanced metal and metalloid uptake and root-to-shoot transfer (phytoextraction, reviewed by Göhre and Paszkowski, 2006; crop biofortification, reviewed by Cavagnaro, 2008), while in other cases, AMF contribute to heavy metal immobilization in the soil through fungal metal-binding processes, thereby improving phytostabilization (Joner et al., 2000). The capacity of AMF in enhancing plant wellness affected by abiotic stresses is well recognized (Leyval and Joner, 2001). Survival studies demonstrated that AMF may protect plants against nonessential elements, besides the enhancing of P and essential elements nutrition, by stimulating plant growth (Smith and Read, 2008).
Because As(V) is a chemical analog of PO4, it is known that AMF might also affect the As mobility in the soil. In AMF-inoculated plants, mycorrhizal PO4 transporters are active (Rausch et al., 2001) and the direct uptake pathway in roots colonized by AMF is often inhibited (Smith et al., 2003). The inhibition of PO4 transporters in roots might promote As(V) tolerance (Sharples et al., 2000; Gonzales-Chavez et al., 2004). Moreover, by enhancing the plant P status, AMF stimulate plant growth, which leads to a dilution of As in the plant (Dong et al., 2008).
Nevertheless, AMF that come from different environments seem to behave differently to As. Generally, AMF collected by contaminated soil are better able to resist the metal toxicity (Gonzales-Chavez et al., 2004). Even though many scientific works indicate that AMF may decrease As concentration in plant tissues and thus limit the loss of yield, still less information is available on the role of the AMF in the As/P interactions at soil–plant interfaces.
Cozzolino et al. (2010) evaluated the growth of lettuce plants in As-contaminated soil from Scarlino (Tuscany, Italy) to examine the influence of P supply and AMF inoculation, by a commercial inoculum, on plant growth and As uptake. The assumption of the study was that AMF inoculation, plus P supply, may have helped lettuce plants to tolerate As of the soil, even improving plant P nutrition.
The highest production of biomass, indeed, was obtained by combining AMF inoculation and PO4 fertilization (Table 4); the sole AMF inoculation increased the biomass of the plants, but the best stimulatory effect on these plants occurred when fertilized with PO4. Therefore, this study revealed that the concomitant supply of AMF and PO4 promoted plant growth and reduced As concentration in the plant, particularly in the aboveground tissues (Table 4).
Modified from Cozzolino et al. (2010). Data are presented as mean values±SD (n=5) and have been analyzed by two-way analysis of variance. Means, followed by the same letter within columns, are not significantly different by Tukey's test at the 5% level. M+ stands for commercial fungal inoculum (100 AMF infective propagules per gram of product, information provided by the supplier) and consisted of a mixture of calcined clay containing spores, hyphae, and root fragments colonized by Glomus intraradices (AEGIS® produced by Italpollina, Rivoli Veronese, Italy.
P+ treatments=0.25 g/pot P (as KH2PO4).
AMF, arbuscular mycorrhizal fungi.
Influence of Trichoderma
Trichoderma spp. includes strains that are known worldwide as highly rhizospere-competent fungi, thus able to colonize the entire root system and actively persist on it for the entire life span of different annual crops (Tucci et al., 2011).
Therefore, several species are extensively exploited in agriculture for plant growth promotion, biological control, and, in general, as positive modifiers of crop metabolism (Harman, 2004; Adams et al., 2007; Hermosa et al., 2012). It has been well established that Trichoderma may enhance the development of the root system (Harman, 2004; Battaglia et al., 2013) by promoting the release of nutrients from the soil (Harman, 2004; Vinale et al., 2013) while generally it seems to improve soil health (Shukla et al., 2008). Recent studies revealed that Trichoderma spp. are able to tolerate moderate concentrations of As in vitro as well as alleviate As-mediated stress on chickpea (Cicer arietinum L.) under greenhouse conditions (Tripathi et al., 2013).
Lately, Caporale et al. (2014) published a research aiming to understand the role of Trichoderma spp. in ameliorating the tolerance of edible plants (i.e., lettuce) to certain abiotic stresses as it might be caused by irrigation of water contaminated with As. These authors evaluated the effect of two strains of Trichoderma (Trichoderma harzianum strain T22 and Trichoderma atroviride strain P1) on the growth of lettuce plants (Lactuca sativa L.) irrigated with As-contaminated water (5 or 10 mg/L of As) and their effect on the uptake and accumulation of the contaminant in the plant roots and leaves. Lettuce is an important leafy vegetable that is consumed primarily fresh or in salads and is considered to be a healthy food. Therefore, As accumulation in its edible portion (i.e., leaves) could pose a risk for the consumer's health.
The growth of lettuce plants was severely influenced by the irrigation with the As-contaminated water (see Table 3 in Caporale et al., 2014). The phytotoxic effect of As determined a strong inhibition of root growth and, to a lesser extent, leaf development. For instance, lettuce plants irrigated with water containing 10 mg/L of As produced less than a third of the root biomass of the control.
On the contrary, a significant plant growth promotion effect was obtained by inoculating lettuce with either strains of Trichoderma (T22 or P1) regardless of the level of As in the irrigation water. Hence, it is evident that the two fungi used, and T22 in particular, were able to mitigate, at least in part, the negative effect of As on plant growth. It is noteworthy that the higher the As level in the irrigation water, the greater the beneficial effect of the inoculation with T22 on plant growth.
In comparison with untreated controls, significantly higher concentrations of As were found in root and leaf tissues when the plants were irrigated with water containing increasing levels of As (Table 5). Most of the As taken up by plants was found in the root system: As concentration in the lettuce roots, indeed, was up to fourfold higher than that observed in the leaves (Table 5). The lower concentration of the metalloid in the leaves may indicate a limited translocation ability of the plant at a systemic level (Smith et al., 2009).
Data are expressed as mean values±SD (n=4) and have been analyzed by two-way analysis of variance. Mean values, followed by the same letter within columns, are not significantly different by Tukey's test at the 5% level. Control, As 5, and As 10 refer to As concentrations of 0, 5, and 10 mg/L, respectively; -T refers to plants noninoculated with Trichoderma; T22 refers to plants inoculated with Trichoderma harzianum; P1 refers to plants inoculated with Trichoderma atroviride.
Percentage of total arsenic content found in the leaves (%) was calculated by dividing the amount of As accumulated in the leaf biomass by the total amount of As stored in the entire plant (leaves+roots)×100.
Although leaf biomass was much more compared with roots (Caporale et al., 2014; Table 3), the amount of As accumulated in the lettuce leaves was always less than 50% of that found in the entire plant (Table 5), with the only exception of As 10-T treatment. The same phenomenon has been reported for several crops, including rice (Xie and Huang, 1998; Liu et al., 2004), bean (Cobb et al., 2000; Caporale et al., 2013), and tomato (Burlo et al., 1999; Pigna et al., 2012), where As content was roots>stems>leaves.
In all cases, plants inoculated with Trichoderma exhibited a significantly lower concentration of As both in the roots and the leaves (Table 5). These results may be related to the abiotic stress tolerance induced in the plant by Trichoderma as supported by enhanced production of biomass and nutrient use efficiency and availability (Harman et al., 2004). The percentage of total As content found in the leaves of the Trichoderma-inoculated plants resulted in significantly lower levels in comparison with the untreated control (Table 5). This may be due to the increased foliar biomass and/or a reduced translocation of the pollutant as a consequence of the beneficial activity of the fungus.
In any case, the treatment with Trichoderma spp. may help to reduce the concentration of inorganic toxicants in the edible portion of crops in addition to alleviating the phytotoxic effect, with a positive outcome on yield and food quality.
Influence of compost
Compost obtained from the organic fraction of municipal solid waste and from other selected waste biomasses can be used effectively as an organic fertilizer on agricultural soil. The high content of stabilized OM and the presence of nutrients of a good quality compost are a guarantee of agronomic advantages; at the same time, the use of compost makes the reuse of organic wastes possible and has numerous advantages (Caporale et al., 2013). The phytonutritive capacity of compost has often been demonstrated to be analogous to that of manure; the same level of productivity, both quantitatively and qualitatively, can be maintained by replacing manure with compost (USDA, 2000; Caporale et al., 2013).
Application of organic by-products as compost to soil increases soil fertility by improving the physical and chemical properties and augmenting microbial activity (and is a common way to dispose of waste). Organic fertility inputs improve soil physical properties by lowering bulk density, increasing water-holding capacity, and improving infiltration rates (Petersen et al., 1999). The addition of compost results in increased mineralization of nitrogen and also micronutrients (Dick and McCoy, 1993). There is a lack of agreement over the influence of OM on As mobility. Cao and Ma (2004) used compost to remediate copper-chromium-arsenate-contaminated soils and recorded an effect of As adsorption, as found by others after compost application (Perez-de-Mora et al., 2007).
Both Mench et al. (2003) and Hartley et al. (2009) observed large increases in leachable As from soils amended with compost probably due to dissolved organic carbon (DOC) competing with As for sorption sites such as Fe oxide surfaces, causing increased As mobility (Redman et al., 2002). DOC can facilitate the transport of contaminants in the cationic form through the formation of soluble metal-DOC complexes (Antoniadis and Alloway, 2002). Although metal-DOC complexes are more mobile in soils, leading to groundwater contamination, these complexes have been shown to be less readily available for plant uptake, thereby alleviating phytotoxicity resulting from excessive metal accumulation in soils (Han et al., 2001).
To evaluate the influence of compost (stabilized for 120 days, derived by decomposition and humification processes of a mix of organic materials, such as pruning waste, organic food sources, waste gardening, and by microorganisms) on the uptake of As by crop plants, Caporale et al. (2013) have studied the response of bean plants to compost application under As pollution.
Bean plants (Phaseolus vulgaris L.) cultivar Borlotto Nano were cultivated in an unheated greenhouse, illuminated with natural light, during the months of late April, May, and June 2011. The experiment provided As application at four rates in irrigation water [0 (As 0), 1 (As 1), 2 (As 2), and 3 (As 3) mg As/L] and three levels of compost application [0 (C 0), 30 (C 30), and 60 (C 60) t/ha]. Bean plant growth (roots, shoots, and beans dry matter production) was significantly affected by As and compost treatments (Table 6). Increasing As concentration in the irrigation water decreased markedly the dry biomass of bean plants, indicating the phytotoxic effect of As.
Data are presented as mean values±SD (n=5) and have been analyzed by two-way analysis of variance. Means, followed by the same letter within columns, are not significantly different by Tukey's test at the 5% level. C stands for the levels of compost supplied to As-uncontaminated soil at the bean plant transplanting (equivalent to 0, 30, and 60 t/ha). As stands for the As concentration in irrigation waters (0, 1, 2, and 3 mg/L).
For plants grown without compost application (C 0), there was a decrease in total biomass of 12.2% (As 1), 37% (As 2), and 50.8% (As 3) compared with the As control (As 0). This loss of biomass was less intense in the C 30 and C 60 treatments, where plants grew 11% and 13.2% (As 1), 27.3% and 24.1% (As 2), and 43% and 35.8% (As 3), respectively, less than their own control treatments (As 0) (Table 6).
Increasing compost application significantly increased bean biomass (Table 6). It is interesting to note that the higher the concentration of As in irrigation water, the greater the increase of bean biomass realized by the application of compost to the soil, indicating the ability of the compost to alleviate the As phytotoxicity. In fact, at the lowest As treatment (As 1), plants amended with compost, (C 30) and (C 60), grew 8.7% and 17.6%, respectively, more than those grown without compost supply (C 0), whereas at the highest As treatment (As 3), they grew 26.0% (C 30) and 53.5% (C 60) more than those of control treatment (C 0).
These authors have also observed that As concentration in roots, shoots, and bean yield rose markedly at increasing levels of As in the irrigation water, especially in the roots (see Table 3 in Caporale et al., 2013). A possible explanation of these findings may be that bean plants showed a typical behavior of the plants sensitive to As toxicity, which usually tend to limit the As translocation from roots to shoots and yield.
They also found that the compost application reduced the As concentration in all tissues of the amended plants compared with the nonamended ones. For instance, at the highest As concentration in irrigation water (As 3), the As concentrations in roots, shoots, and bean yield were 4.79, 2.68, and 1.08 mg/kg, respectively, in C 0 treatment, 4.36, 2.13, and 0.86 mg/kg, respectively, in C 300 treatment, and 3.87, 1.71, and 0.63 mg/kg, respectively, in C 60 treatment. Plants amended with compost showed a lower As uptake likely because As mobility was reduced through the adsorption on the OM of the compost.
Conclusions
This review has attempted to study the fate of As in the soil environment and to describe the main sorption/desorption processes on or from soil components, controlling its speciation, mobilization, and phytoavailability. This work has also indicated that the use of biofertilizers (i.e., AMF, Trichoderma, and compost), besides improving soil fertility, may alleviate the As phytotoxic effect in important food crops stressed by As.
Despite the information already available in literature, in future there is need of interdisciplinary research to better understand the biogeochemical mechanisms governing As dynamics in the soil–microorganism–plant system by using advanced analytical techniques.
Footnotes
Acknowledgment
This work was supported by the Italian Research Program of National Interest (PRIN), year 2010–2011 (Grant No. 2010JBNL17_005).
Author Disclosure Statement
No competing financial interests exist.