
Abstract
Abstract
The present study investigated fluorene biodegradation by Sphingobacterium sp. KM-02 in the presence of heavy metals. A fluorene-degrading strain of Sphingobacterium was isolated from polycyclic aromatic hydrocarbon-contaminated soil near a mine-impacted area in Korea. When Sphingobacterium sp. KM-02 was grown in a medium with fluorene as the sole carbon source, it removed 78.4% of this compound within 120 h. Identification of the metabolic intermediates (9-fluorenone, 4-hydroxy-9-fluorenone, and 8-hydroxy-3,4-benzocoumarin) were also performed. Composting experiments under laboratory conditions indicated that this microbe also removed fluorene from contaminated soil. In particular, treatment of microcosm soil with strain KM-02 for 20 days resulted in a 65.6% reduction of fluorene concentration. Experiments of effects of heavy metals on fluorene removal by strain KM-02 showed that 10 mg/L cadmium, copper, zinc, and lead, reduced growth and fluorene degradation by this microbe. Cadmium and copper had strong effects at 10 mg/L, although zinc and lead had relatively slight inhibitory effect at concentrations of 10 and 100 mg/L. Arsenic had no effect on the growth or fluorene degradation, even at 100 mg/L.
Introduction
P
It is a major component of fossil fuels and has been detected in vehicle exhaust, air particulates, and marine sediments (Keith and Telliard, 1979; Wattiau et al., 2001). Generally, high-molecular weight PAHs (four or more rings), such as pyrene, benzo[b]fluoranthene, and benzo[g,h,i]perylene, are strongly sorbed to particles, and this greatly reduces their adverse effects. However, low-molecular weight PAHs (two or three rings), such as naphthalene, fluorene, and phenanthrene, are water soluble and can occur in groundwater or surface water, and pose environmental threats (Achten and Hofmann, 2009; Mostert et al., 2010). Thus, the degradation or sequestration of low-molecular weight PAHs in the environment is an important priority. Among the low-molecular weight PAHs, given priority by the EPA, fluorene (C13H10) and phenanthrene (C14H10) are most commonly found at contaminated sites (Baath, 1989; Amor et al., 2001).
The fluorene in this soil could have been from various chemicals and/or incineration used for ore dressing or smelting during mining activity several decades ago. PAHs and fluorene are also generated from incomplete incineration following wildfires (Storelli and Marcotrigigano, 2000; Antizar-Ladislao et al., 2006).
Numerous microorganisms such as Pseudomonas, Arthrobacter, Staphylococcus, Mycobacterium, Brevibacterium, Burkholderia, and Acinetobacter can metabolize fluorene through various intermediates (Monna et al., 1993; Grifoll et al., 1994; Casellas et al., 1997; Wattiau et al., 2001; Pagnout et al., 2007; Schuler et al., 2008; Thangaraj et al., 2008). Previous studies indicated that biodegradation of fluorene can proceed by three different pathways. First, the initial 1,2-dioxygenation of fluorene forms fluorene-1,2-diol, and this is transformed to 3-chromanone through 2-hydroxy-4-(2-oxo-indan-1-ylidene)-2-butenoic acid, 1-formyl-2-indanone, 2-indanone-1-carboxylic acid, and 2-indanone (Monna et al., 1993; Casellas et al., 1997). Second, the initial 3,4-dioxygenation of fluorene leads to salicylate formation through 2-hydroxy-4-(1-oxo-indan-2-ylidene)-2-butenoic acid, 2-formyl-1-indanone, 1-indanone-2-carboxylic acid, 1-indanone, 2-chromanone, and 3-(2-hydroxy-phenyl)-propionic acid (Grifoll et al., 1995; Casellas et al., 1997). Third, the initial C-9 monooxygenation in Brevibacterium sp. DPO1361 and Pseudomonas sp. F274 (which is only productive if there is a subsequent angular carbon dioxygenation) leads to the formation of phthalate, and then protocatechuate (Boldrin et al., 1993; Grifoll et al., 1994; Trenz et al., 1994; Grifoll et al., 1995; Wattiau et al., 2001). Previous research has identified several genes involved in the degradation of fluorene to phthalate in Terrabacter sp. DBF63 (Habe et al., 2004).
Abandoned mines pose environmental hazards worldwide (Thornton, 1996). In many parts of the world where mining activities have shut down, there is a need to control and reclaim mining areas that were polluted with acidic mine drainage and heavy metals (El Adnani et al., 2007; Moreno-Jimenez et al., 2009). Research in bioremediation has sought to isolate new strains of microorganisms and develop more successful protocols for reducing metal toxicity with heavy metal-resistant specificity. The soils of former gold and silver mines are mainly contaminated with arsenic, because these precious metals often occur in arsenic-rich regions; cadmium contamination is common in mines associated with copper, lead, and zinc extraction. Heavy metal pollution is a very serious problem, so the effects of heavy metals on the biodegradation of organic pollutants, such as fluorene, must be examined.
Mining soils have considerable amounts of heavy metals such as cadmium, lead, zinc, arsenic, and copper (Thornton, 1996; El Adnani et al., 2007; Moreno-Jimenez et al., 2009). Although low concentrations of these metals are necessary for most organisms, high concentrations can cause significant toxicities (Nies, 1999). Numerous studies showed that a wide range of concentrations of various metals, such as Al, Co, Ni, Cu, Zn, and Pb, have adverse effects on the degradation of organic compounds (Baath, 1989; Nies, 1999; Mittal and Ratra, 2000; Amor et al., 2001; Sokhn et al., 2001). However, there is limited information on the effect of individual heavy metals on the biological degradation of PAHs, such as fluorene. Moreover, the effects of heavy metals during biodegradation in former mining areas have not been considered. Most heavy metals are adsorbed on soil particles, and some heavy metals (arsenic and chromium) can be oxidized to anionic forms that freely move into the soil environment. Currently, we are developing a remediation system for former mining soils that includes a slurry-based acid mine drainage detoxification reactor for the removal of heavy metals, and single or mixed inoculations of microbes. Thus, there is a need for examination of the effect of heavy metals on the biodegradation of organic pollutants to develop the (in)organic pollutants detoxification reactor successfully in the future study.
The present study examined Sphingobacterium sp. KM-02-mediated degradation of fluorene, in the liquid or solid phase, in the presence of five common heavy metals, and with the use of mass spectrometry to identify all metabolic intermediates. This bacterium was isolated from former mining soils that are contaminated with heavy metals and PAHs, and can metabolize fluorene to 9-fluorenone, 4-hydroxy-9-fluorenone, and 8-hydroxy-3,4-benzocoumarin. We also performed a microcosm study to document bioavailability for in situ remediation.
Experimental Protocol
Reagents
Inductively coupled plasma (ICP) standard solutions (SPEX plasma standard, 1,000 mg/L of H3AsO4, Cd, Cu, Pb(NO3)2, and Zn) in 2% nitric acid were purchased from SPEX) and employed as sources of heavy metals and standards for ICP mass spectrometry (ICP-MS; Thermo Elemental). Fluorene and its metabolites were purchased from Sigma. Methylene chloride, ethyl acetate, n-hexane, and acetone were all analytical grade (for organic trace analysis), and purchased from Merck. The nutrient broth, ortho-phosphoric acid (85%), and agar media were also from Merck. Minimal salts medium (MSM) was prepared as described previously (Nam et al., 2006, 2008), and contained 3.5 g Na2HPO4 2H2O, 1 g KH2PO4, 0.5 g (NH4)2SO4, 0.1 g MgCl2 6H2O, 50 mg Ca(NO3)2 4H2O, and trace elements per liter of distilled water. All other chemicals and compounds were of the highest grade that was commercially available.
Isolation and identification of bacteria
Fluorene-degrading aerobic bacteria were collected from soil in the Kangwon province that was contaminated with PAHs and heavy metals. This region is in the eastern part of the Republic of Korea, and was greatly impacted by a nearby copper mine. A mixed culture obtained from soil samples served as the starting material for isolation of pure cultures. Soil samples were placed in sterilized 50-mL plastic centrifuge tubes and were not refrigerated or frozen during transportation or storage. One-gram soil samples were added to 10 mL of phosphate buffer (pH 7.0). After severe shaking, particles were removed by centrifugation at 1,000 g for 10 min. One hundred microliters of the mixed culture was then inoculated into 100 mL of MSM with fluorene (10 mg/L) and incubated overnight.
For selective enrichment, 100 μL of the liquid culture was streaked onto solid MSM agar medium with 10 mg/L fluorene. After 3 days of incubation, each colony was inoculated into 1 mL of MSM that contained 1 mM fluorene as the sole carbon source in a 15-mL centrifuge tube with a silicon stopper. Single colonies and defined consortia were tested for their ability to metabolize fluorene.
To identify the newly isolated strain, genomic DNA was extracted using a DNA extraction kit (NucleoSpin; Macherey-Nagel) from the strain, according to the manufacturer's instructions. The DNA extraction protocol included a 10 min incubation of the extraction mixture at 70°C before physical disruption of the cells, a phenol extraction step to remove protein, and a precipitation step. The DNA recovered from each aliquot was combined into a single sample, and the quality and quantity of the extracted DNA were assessed by gel electrophoresis. The 16S rRNA gene was polymerase chain reaction amplified using the universal primers 27F (5′-AGAGTTTGATCMTGGCTCAG-3′) and 1492R (5′-TACGGYTACCTTGTTACGACTT-3′) (Heuer et al., 1997) and the amplification protocol consisted of an initial denaturation at 98°C for 60 s, followed by 35 cycles of denaturation at 98°C for 10 s, annealing at 56°C for 30 s, and extension at 72°C for 30 s, followed by a final extension at 72°C for 10 min. The amount of recovered DNA was estimated by agarose gel electrophoresis. The amplified gene was sequenced and deposited in the GenBank at the National Center for Biotechnology Information (NCBI) under accession number FJ548749.
Growth of strain KM-02 on fluorene
Sphingobacterium sp. KM-02 was selected from among a number of bacterial isolates obtained from this fluorene-contaminated soil by selective enrichment culture. The growth of strain KM-02, which can use fluorene as the sole carbon source, was investigated by culturing in 100-mL Erlenmeyer flasks containing 10 mL of sterilized MSM and 10 mg/L fluorene in crystal form. For fluorene biodegradation experiments, 10 mL of KM-02 cell suspension (OD600 = 0.1) was added to 100-mL Erlenmeyer flasks (in triplicate) that contained 10 mg/L of fluorene, and incubated at 160 rpm and 28°C for 5 days. Cells subjected to heat inactivation (70°C for 40 min) and poisoning (10 mM NaN3) were used as controls (Nam et al., 2006). Control experiments with NaN3-poisoned KM-02 cells showed that abiotic processes did not reduce the fluorene concentration.
The colony-forming units per milliliter (CFU/mL) were measured by conventional plating on 36 solid MSM agar media, including fluorene (10 mg/L), at 0, 24, 48, 72, 96, and 120 h. At each time point, the six culture media, including controls, were collected and stored at −70°C until the final flasks were collected at 5 days, and 36 samples were subjected to CFU and chemical analyses for the measurement of growth of strain KM-02, fluorene degradation, and metabolite formation. All experiments were performed by triplicate independently, and the levels of fluorene and its metabolites were quantitatively determined by gas chromatography–mass spectrometry (GC-MS) analysis as described below in the Analytical Methods section.
Microcosm experiments
A second experimental series was conducted in a soil microcosm with isolated KM-02 to evaluate the ability of this strain to metabolize fluorene in actual mining soil. A plastic container with 1 kg of dry and autoclaved soil was supplemented with fluorene from acetone stock solutions to a final concentration of 100 mg/kg soil. The acetone was allowed to evaporate and then the soil was mixed thoroughly. Then, 200 mL of pregrown on MSM with 10 mg/L fluorene and harvested KM-02 cells (OD600 = 2.0) were added to each container and mixed. The KM-02 cells were washed three times with MSM before use. Two control experiments were performed with heat inactivation (70°C for 40 min) and poisoning (10 mM NaN3) of KM-02 cells, and with cell-free mine soil. Every 5 days, 2 g of soil was collected from the soil microcosms and stored at −20°C. After 15 days, all samples were extracted, and the concentrations of fluorene were determined as described below in the Analytical Methods section. Dibenzofuran was used as internal standard to determine recovery rate. All data are reported as the mean of three measurements.
Effects of heavy metal on the growth of strain KM-02 and fluorene biodegradation
For measurement of the rates of growth and fluorene degradation, cells pregrown with fluorene were inoculated into Erlenmeyer flasks that contained 10 mg/L of fluorene and heavy metals (10 or 100 mg/L). One hundred milligram per liter stock solutions were prepared by 10-fold dilution of a 1,000 mg/L solution with 2% nitric acid. An autoclaved heavy metal solution (2 mL) was added to 18 mL of culture solution in a 250-mL Erlenmeyer flask. The final concentration of fluorene in the cultures was 10 mg/L. Flasks were incubated on an orbital shaker for 5 days (28°C, 160 rpm). The initial optical density at 600 nm was adjusted to about 0.1 diluting with MSM. The control cultures contained 2% nitric acid and no heavy metals. The optical density at 600 nm was measured every 24 h for 120 h. Fluorene concentrations were measured as described below.
Analytical methods
Extraction of PAHs from the sample soil used for isolation of KM-02 was determined by an accelerated solvent extraction (ASE) System (Dionex). Extraction of 2 g of soil with the same amount of sodium sulfate (Na2SO4) was performed using a methylene chloride:acetone (1:1, v/v) extraction ratio and ASE conditions of 100°C, 1,500 psi, static time of 10 min, flush volume of 60%, purge time of 60 s, and three static cycles. The soil extract was then concentrated by N2 purging and prepared in n-hexane for silica gel column chromatography. The extracted and column-treated samples were concentrated by N2 purging and analyzed for PAHs using an Agilent GC 6890N/MSD 5973 equipped with a 60-m DB-5 capillary column. All qualitative and quantitative analyses of PAHs were compared with 16 authentic PAH standards (AccuStandard).
GC-MS analysis was also used for quantitative analyses of the depletion of fluorene during bacterial growth and of the effect of heavy metals on the growth of KM-02 cultures. In these experiments, 1 mL of 85% ortho-phosphoric acid was added to the culture flasks to stop the reaction after various times of growth, and these flasks were immediately stored at −70°C. After the last sample was collected, all flasks were thawed and liquid–liquid extractions were conducted. The flasks and controls were rinsed six times with methylene chloride:acetone (1:1, v/v), extracts were dried over anhydrous Na2SO4, and the organic solvents evaporated under reduced pressure. GC-MS was used to identify fluorene and its metabolites based on comparison with commercial standards. In the GC-MS analysis, the initial temperature was 80°C for 3 min, the temperature increased to 280°C at 10°C/min, and the temperature was maintained at 280°C for 5 min. All mass spectra were obtained at an ionization energy of 70 eV, and the temperature of the ion source was maintained at 250°C.
For microcosm samples, fluorene was extracted as previously described. For determination of the heavy metal content, 5 g of dried mine soil was collected, placed in Teflon vessels, and extracted in a microwave oven using concentrated nitric acid (HNO3), according to EPA Method 3051. After cooling, samples were filtered and diluted by using ultra-pure water. Metal determination was performed by ICP-MS (Thermo Elemental). All samples were analyzed by ICP-MS in triplicate to check precision, which was within 5%. Final results (mean of three values from ICP-MS), are expressed as mg/kg (dry weight). The concentrations of heavy metals in the flasks after incubation and prepared under the same conditions were measured using ICP spectrometry.
Results and Discussion
Isolation of strain KM-02 from mine-impacted soil and characteristics of the soil
Sphingobacterium sp. KM-02 was isolated from PAH-contaminated soil near a former copper mine in Kangwon province, Republic of Korea. This study analyzed the 16 PAHs originally specified as priority pollutants by the U.S. EPA. There were a total of 10 PAHs in the contaminated soil, the total PAH concentration was 129.0 ng/g dry weight of soil, and fluorene was the most abundant PAH (46.9 ng/g, 36.6%) (Table 1). Six of the 16 EPA-designated PAHs, Benzo(a)pyrene, Benzo(g,h,l)perylene, Benzo(k)fluoranthene, Chrysene, Dibenz(a,h)anthracene, and Indeno(1,2,3-cd)pyrene, were not detected. Previous studies reported fluorene soil contamination levels of 3–180 μg/g (Achten and Hofmann, 2009; Mostert et al., 2010), mostly higher than in the present study. However, the fluorene concentration in the present study was somewhat higher than in some other mine-impacted soils (Wang et al., 2010).
PAH, polycyclic aromatic hydrocarbon.
The mine soil was also contaminated with several heavy metals (Fig. 1), especially arsenic (121.65 mg/kg). Zinc (62.38 mg/kg), lead (47.02 mg/kg), copper (40.94 mg/kg), cadmium (13.52 mg/kg), manganese (9.65 mg/kg), chromium (2.35 mg/kg), and cobalt (1.95 mg/kg) were also present. We examined the effects of the five most abundant heavy metals on metabolism of fluorene by a strain of bacteria designated as KM-02. When grown on nutrient agar medium, KM-02 colonies exhibited a dark yellow pigmentation with high viscosity. 16S rRNA gene analysis using BLAST indicated that KM-02 had high sequence similarity to Sphingobacterium sp. (99.1% homology). This sequence was deposited in the NCBI gene bank (accession No. FJ548749).
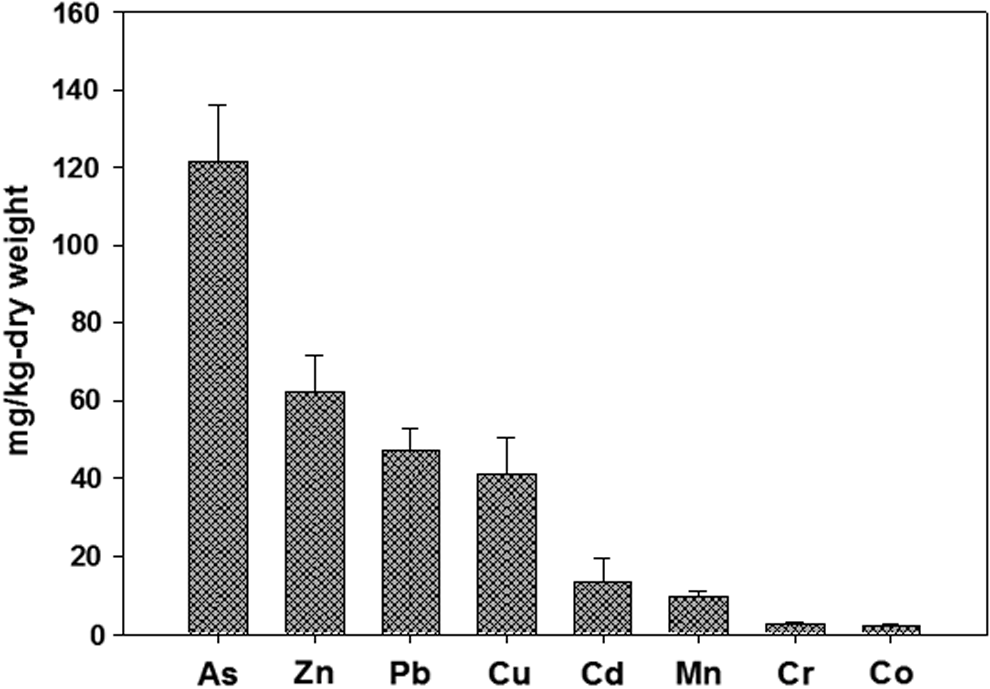
Heavy metal concentrations in mine-impacted soil containing Sphingobacterium sp. KM-02.
Growth of the strain KM-02 on fluorene
The bacterial growth and fluorene removal of Sphingobacterium sp. KM-02 under aerobic conditions are shown in Fig. 2. These results show that the biomass (CFU/mL) increased as fluorene was removed. Within 5 days, the initial quantity of fluorene (10 mg/L) was 78.4% lower, and the cell density stabilized at about 3.0 × 107 CFU/mL. Thus, KM-02 cells appear capable of using fluorene as a carbon source, and this microbe may play an important role in the natural degradation of fluorene in soils contaminated by mine activities. Catabolism of fluorene by this microbe gave rise to three polar intermediates: 9-fluorenone (Fig. 3a), 4-hydroxy-9-fluorenone (Fig. 3b), and 8-hydroxy-3,4-benzocoumarin (Fig. 3c). Previous studies suggested that 8-hydroxy-3,4-benzocoumarin could be completely mineralized into CO2 and H2O through phthalic acid (Grifoll et al., 1995; Wattiau et al., 2001). Based on the results, strain KM-02 and the fluorene-degrading species of other genera (Boldrin et al., 1993; Grifoll et al., 1994, 1995; Trenz et al., 1994; Wattiau et al., 2001) may share a fluorene biodegradation pathway.

Growth of Sphingobacterium sp. KM-02 with fluorene as sole carbon source. Here and below, all values represent the mean of three independent experiments. Symbols: CFU/mL of untreated cells (●), CFU/mL of poisoned cells (◯; 10 mM sodium azide and heating), fluorene concentration with untreated cells (■), and fluorene concentration with poisoned cells (□).
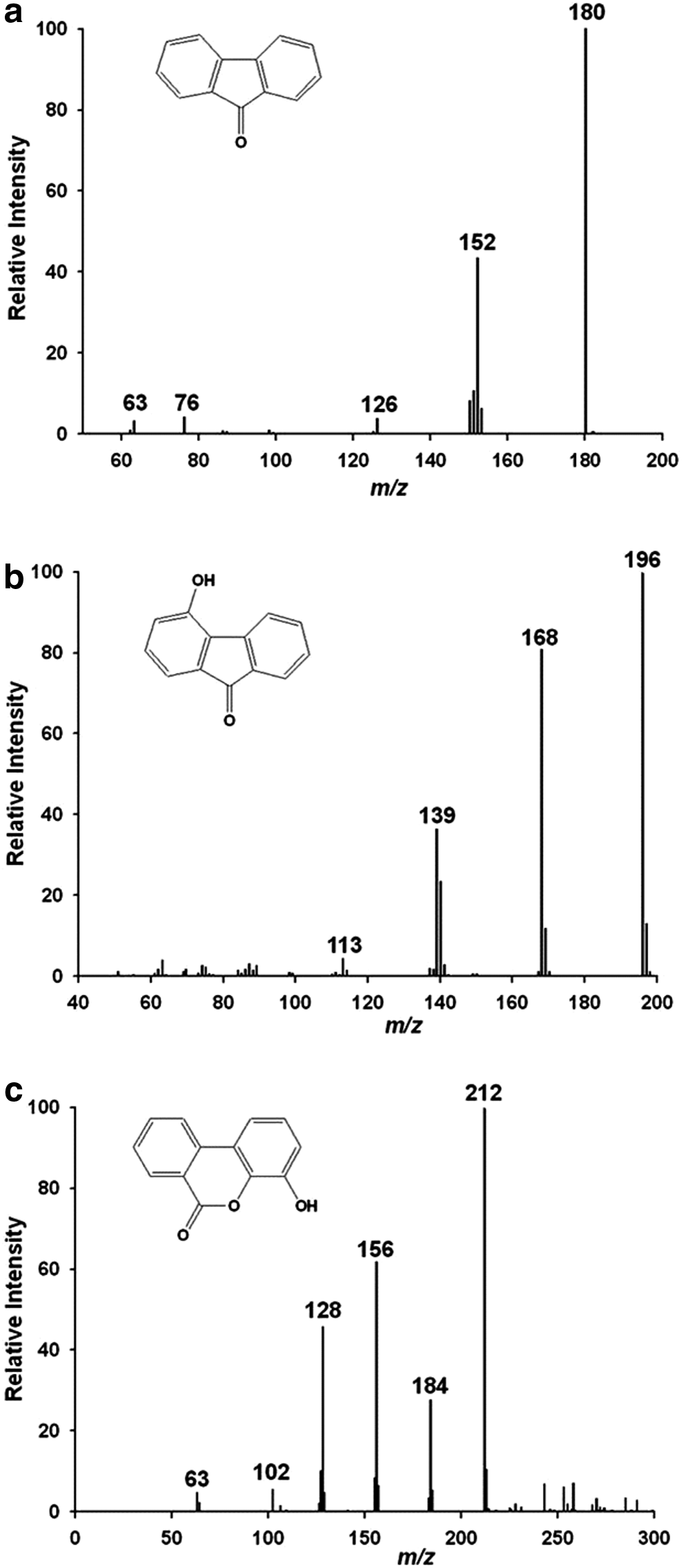
Fluorene metabolites produced by Sphingobacterium sp. KM-02.
Removal of fluorene from a mine soil microcosm
We performed a microcosm study to examine the ability of this microbe to remove fluorene from actual mine soil (Table 2). After 20 days of incubation with 100 mg/kg fluorene, KM-02 cells removed 65.6% of the fluorene, indicating the presence of catabolic activity in this microcosm. The controls (inactivated KM-02 cells) decreased the fluorene level by 25.4%, possibly due to adsorption to dead biomass or soil particles. This result is consistent with previous reports that microbial adsorption plays an important role in the removal of recalcitrant pollutants and that the adsorption by dead biomass is an important factor in toxin removal (Tyler, 1974; Hattori, 1992; Sokhn et al., 2001; Lu and Zhu, 2012). This effect may be due to the affinity of fluorene for certain biomolecules that remain in inactivated cells (e.g., lipids) and/or because fluorene is adsorbed to soil particles due to its low solubility (Nam et al., 2005; Gan et al., 2009). This finding should be considered in future studies of the potential use of strain KM-02 in the bioremediation of soils contaminated with PAHs. Strain KM-02 is the first microbe to be isolated from an area contaminated by a metal mine that is capable of fluorene degradation. Thus, this organism appears to have great potential for use in the remediation of mining soils contaminated with PAHs. In particular, strain KM-02 is a potential candidate for development of a biological treatment technology for the remediation of soils contaminated with fluorene and PAHs. Further biochemical and genetic studies of strain KM-02 may improve our understanding of fluorene degradation in nature and help to optimize procedures for the remediation of PAH-contaminated mining soils.
Fluorene was determined by gas chromatography–mass spectrometry analysis.
Mean and standard deviation of three replicate determinations.
%b = [(Control − with dead cells)/Control] × 100.
%c = [(Control − with live cells)/Control] × 100.
Effects of heavy metal on growth of strain KM-02
We determined the effect of two concentrations of five heavy metals on the growth of Sphingobacterium sp. KM-02 in liquid cultures with fluorene as the sole carbon source. The addition of 10 mg/L copper, cadmium, zinc, and lead significantly inhibited microbial growth, although arsenic had no effect, and lead only had a slight inhibitory effect (30%). Notably, zinc inhibited growth by 66% compared to controls (no heavy metals or only 2% nitric acid solution) and copper and cadmium completely inhibited growth (Fig. 4a). Experiments with 100 mg/L heavy metals yielded similar results (Fig. 4b), with 61% inhibition by lead and 63% inhibition by zinc. Previous studies also reported that metal ions can inhibit the metabolic activity of microorganisms involved in the degradation of organic compounds (Hattori, 1992; Amor et al., 2001). The extent of inhibition depends on the concentration and availability of heavy metals and other factors, including the type of metal, growth medium, and microbial species or strain specificity (Tyler, 1974; Chen et al., 2013). Further, several microorganisms have high resistance to heavy metals (Sultan and Hasnain, 2003). In the case of arsenic, a previous report showed that arsenic-resistant bacteria survived in an area with high arsenic contamination due to enzyme specificity (Pepi et al., 2007).

Effects of 10 mg/L
Effects of heavy metals on biodegradation of fluorene
Next, we determined the effects of heavy metals on fluorene degradation rate by strain KM-02 by measuring the residual fluorene concentration in growth medium that had 10 mg/L of fluorene as the sole carbon source (Fig. 5). The fluorene removal rates in the presence of the different heavy metals (10 mg/L) had the same general trends as growth of strain KM-02 in the presence of the different heavy metals. In particular, the fluorene concentration decreased to 3.21 mg/L after 120 h in the presence of 10 mg/L arsenic, similar to the level when cells were grown in a medium without heavy metals (2.32 mg/L) or with 2% nitric acid (2.29 mg/L). After 120 h of growth in 10 mg/L of the other heavy metals, the residual fluorene concentrations were lower for Pb (5.56 mg/L), Zn (7.26 mg/L), Cu (9.84 mg/L), and Cd (10.02 mg/L). Notably, fluorene degradation (Fig. 5a) and cell growth (Fig. 4a) were completely inhibited by 10 mg/L copper or cadmium. For an arsenic concentration of 100 mg/L, fluorene degradation was similar to that at 10 mg/L (Fig. 5b). However, degradation was completely inhibited by 100 mg/L copper and 100 mg/L cadmium, and slightly inhibited by zinc (72.5%) and lead (70.2%). Thus, the results of Figs. 4 and 5 indicate that the heavy metal-induced inhibition of growth and fluorene degradation by strain KM-02 can be described as: cadmium = copper > zinc > lead > arsenic. Future studies should investigate the mechanism of arsenic resistance in strain KM-02. Arsenic is a metalloid widely distributed in soils and natural waters that can occur naturally from the weathering of rocks, or from anthropogenic sources such as mining and agriculture (Pepi et al., 2007). Although arsenic is toxic to most organisms, some microorganisms use arsenic compounds as electron donors, as electron acceptors, or have arsenic detoxification mechanisms (Ji and Silver, 1992; Pepi et al., 2007).

Effects of 10 mg/L
Field studies are needed to assess the effects of different heavy metals upon different bacterial strains. For example, one study showed that 0.4 mM copper had no effect on the degradation of phenanthrene, but inhibitory effects were observed at a copper concentration of 4.4 mM (Sokhn et al., 2001). Other studies showed that low concentrations of copper (below 0.1 mM) inhibited the degradation of dibenzofuran or tert-butyl ether (Lin et al., 2006; Hong et al., 2007). In another study, diesel removal was significantly reduced at a copper concentration above 0.5 mM due to the reduced rate of oxygen consumption (Riis et al., 2002). Many researchers showed that copper accumulation decreases the activity of microbial dioxygenase (Said and Lewis, 1991). These results are consistent with the results of the present study, which showed that 10 mg/L copper inhibited the growth and fluorene degradation by strain KM-02.
Cadmium has well-known toxic effects comparable to other heavy metals, such as copper, zinc, and lead (Baath, 1989). Cadmium can attach to microbial proteins in the sulfhydryl functional group or glutathione, thereby interfering with cysteine synthesis, or by directly damaging DNA (Helbig et al., 2008). Also, cadmium can easily penetrate the cell membrane of the bacteria, in the metabolism of essential substances, extracellular or intracellular emissions, by reducing the permeability of substances (Bruins et al., 2000). The results of the present study, which show that cadmium inhibited growth and fluorene degradation by strain KM-02, is consistent with these previous studies. Another possible explanation for the detrimental effects of heavy metals is that they affect the biochemical oxygen demand due to oxidation of the metal. In particular, dissolved oxygen may be consumed during the oxidation of organic matter or by reaction with metal ions, resulting in oxide formation (Mittal and Ratra, 2000; Hong et al., 2007). Therefore, the oxidation of metal ions may affect the rate of PAH degradation and growth. However, we believe that metal oxidation cannot explain the results of the present study because copper and cadmium have very small oxidation potentials, but had the strongest effects on cell growth and fluorene biodegradation. Although it is possible that copper and cadmium are oxidized to precipitates under aqueous conditions, this mechanism can only be confirmed by analysis of reaction solutions after dissolution in strong acids.
Conclusions
We isolated a strain of bacteria from an abandoned mine in Korea that was contaminated with PAHs and heavy metals that could degrade fluorene. Based on 16S rRNA gene sequence analysis, the bacterium was Sphingobacterium sp., and designated as strain KM-02. KM-02 cells grown in culture with fluorene as the sole carbon source led to removal of 78.4% of this compound within 120 h. Identification of the metabolic intermediates (9-fluorenone, 4-hydroxy-9-fluorenone, and 8-hydroxy-3,4-benzocoumarin) were performed by GC-MS analysis. Incubation of the KM-02 strain in a mine soil microcosm for 20 days reduced the concentration of fluorene from 100 to 31.4 mg/g (65.6%). We also assessed the effect of five heavy metals on cell growth and the biodegradation of fluorene by incubation of strain KM-02 in liquid medium with arsenic, zinc, lead, copper, and cadmium. Cadmium and copper strongly inhibited growth and fluorene degradation, zinc and lead had modest inhibitory effects on growth and fluorene degradation, and arsenic had no effect on growth and fluorene degradation.
This study demonstrates the removal of a pollutant in the presence of abundant PAHs and heavy metals of a mine-impacted area, and the data reported here show that strain KM-02 consistently reduced fluorene concentration. Taken together, the results presented here suggest that strain KM-02, which is the first fluorene-degrading isolate identified from a copper mine-impacted soil, may be a potential candidate for the industrial elimination of PAHs from contaminated soils. However, in situ studies that examine the adsorption and degradation mechanisms are necessary before this strain can be used in practice.
Footnotes
Acknowledgment
This work was supported by the Korea Institute of Geoscience and Mineral Resources (KIGAM) research fund under Grant No. 3451.
Author Disclosure Statement
No competing financial interests exist.