
Abstract
Abstract
This article reports the isolation and characterization of two strains of bacteria from a hydrocarbon-contaminated industrial area (Cubatão-Brazil), which were able to degrade gasoline compounds, such as benzene, toluene, and ethylbenzene (BTE), the highest water-soluble fractions of commercial gasoline. Among the isolates, 16S rRNA gene sequence analysis revealed that these two strains belong to the gamma group of proteobacteria, with 95.0% and 99.0% similarity to the 16S rRNA gene sequences of Serratia sp. (HCS1) and Pseudomonas sp. (HCS2), respectively (BLAST, GenBank, NCBI). Degrading potential of each isolate was measured by inoculating pure culture in a mineral medium containing BTE or commercial gasoline (containing ethanol 24% [v/v]) as the only carbon source and monitoring hydrocarbon disappearance rate at regular time intervals. Growth of isolates in mineral medium containing hydrocarbons was monitored by measuring the turbidity (OD 600 nm) and substrate consumption by high-performance liquid chromatography (in the BTE assays) and gas chromatography–mass spectrometry (gasoline biodegradation assays). After 15 days, BTE (50 mg/L) were totally degraded by both strains. After 21 days, HCS1 and HCS2 had also degraded commercial gasoline by 60% (v/v) and 80% (v/v), respectively. Results suggest that these strains have a high level of tolerance to hydrocarbon toxicity and, consequently, have potential applications in the biotreatment of industrial wastewaters or in situ bioremediation of hydrocarbon-contaminated soils.
Introduction
O
The benzene, toluene, and ethylbenzene (BTE) group of contaminants consists of benzene, ethylbenzene, and toluene. These organic chemicals, including BTE, are major components of gasoline and are thought to be the most prevalent contaminants of soil and groundwater due to frequent leakages from underground storage tanks and accidental spills (Choi et al., 2013). The contamination of soil and groundwater systems by gasoline is considered a serious problem, since the compounds have long-term toxic effects. Besides its toxicity, benzene is also known to be a carcinogen (Garcia et al., 2015).
According to Schneider et al. (2006), solid and other residues from petroleum processing and other operations (building materials, organic waste, etc.) were disposed of on land in Cubatão until 1986. Moreover, a significant number of sites contaminated by crevice corrosion in gasoline storage tanks have been identified by CETESB, an environmental regulatory agency in Brazil.
Furthermore, Brazilian commercial gasoline contains ethanol (24% added), which increases the aqueous solubility of these hydrocarbons, making soil and groundwater contamination a more critical issue (Oliveira et al., 2011). An elevated load of petroleum hydrocarbons in soil causes a significant decline in soil quality, making these soils unusable. To control the environmental and health risks caused by petroleum products, many new regulations have been introduced and research on remediation and bioremediation of soil has substantially increased.
In particular, bioremediation is being increasingly used in the treatment of areas contaminated by oil spills. Despite a relatively long history of research on oil spill bioremediation, it is still an essentially empirical technology and many of the factors that control bioremediation have yet to be properly understood; contaminated sites, for example, demand diverse strategies and development of new technologies that are efficient and cheap enough to make remediation feasible even in remote areas and where financial support is limited (Jonsson and Haller, 2014).
Biodegradation by natural populations of microorganisms or in situ attenuation is a primary mechanism by which petroleum hydrocarbons could be eliminated from contaminated sites (Wongsa et al., 2004). Microorganisms isolated from contaminated areas have been used to improve biodegradation. Hendrickx et al. (2005) isolated fluorescent Pseudomonas strains, especially Pseudomonas putida strains, from BTEX- and gasoline-contaminated sites that degrade petroleum derivates metabolically.
Collins et al. (2002) observed the total mineralization of benzene and toluene, and incorporation was enhanced by methyl substituents of toluene-derived catechols (Srijata and Pranab, 2011). Two main pathways have been described for the aerobic degradation of BTE in bacteria leading to their mineralization. Benzene can be metabolized by toluene dioxygenase (tod); however, toluene may subject to oxidation by either the toluene dioxygenase or the toluene monooxygenase (tol) (Coombs and Franco, 2003).
Aerobic and anaerobic biodegradation of several individual gasoline components has been studied, often with pure microbial cultures and numerous isolates having been found, which can be used by individual gasoline hydrocarbons as a carbon source. However, little work has been done on the biodegradation of the whole commercial product. This could be due to the challenge of resolving the many components in gasoline and experimental difficulties in maintaining aerobic conditions while preventing the volatilization of the smaller hydrocarbons.
This article describes the effect of inoculation of Serratia marcescens and Pseudomonas aeruginosa bacterial strains on the biodegradation of a mixture of BTE and commercial gasoline in aqueous solution. These bacteria were previously isolated from an area contaminated with commercial gasoline by the selective enrichment culture technique, which was able to degrade the hydrocarbon components in liquid culture.
Experimental Protocols
Isolation and identification of hydrocarbon-degrading microorganisms
Soil samples were collected from two different hydrocarbon-contaminated areas in Cubatão, Brazil, during winter time (June). Accurately weighed samples consisting of about 1 g were diluted and inoculated into 100 mL of minimal salt medium (MSM) consisting of FeSO4 (10 μM), MgSO4 (1 mM), CaCl2 (0.5 mM), NH4Cl (0.9 mM), Na2HPO4 (1.2 mM), and KH2PO4 (0.8 mM) containing 50 mg/L of benzene, 50 mg/L of toluene, and 50 mg/L of ethylbenzene (totaling 150 mg/L of contaminant) in 150-mL sealed flasks and incubated at 30°C under constant shaking at 200 rpm in an orbital shaker for 30 days. Aliquots of the grown bacterial culture were then plated on MSM agar plates containing BTE and incubated overnight at 30°C. Growing colonies were purified by repeated streaking onto agar plates. The isolated strains were maintained on nutrient agar at 4°C.
Identification of bacterial isolates with the ability to degrade BTE as the only carbon source was performed on the basis of microscopic examination and MALDI-TOF spectrometry (Matrix-Assisted Laser Desorption/Ionization–Time of Flight spectrometry). The bacterial strains were isolated according to colonial morphology, surface, shape, size, margin, and pigmentation on the nutrient agar medium. Molecular analysis of 16S rRNA was used to confirm the biochemical characterization of isolated bacterial strains, once polymerase chain reaction (PCR) was performed with the universal 16S primers designed to amplify the region between positions 27 and 1492R of the 16S rRNA gene.
Two genomic DNA of the isolated strains were extracted according to the Chen and Kuo (1993) procedure and then identified through analysis of 16 rRNA gene sequencing using the universal primers designated as 27F (5′ AGAGTTTGATCCTGGCTCAG 3′) and 1492R (5′ ACCTTGTTACGACTT 3′) (Yu et al., 2013). The PCR was carried out in 25 μL reaction volumes with the following reagents: 1 μL of 27F (100 ng/μL), 1 μL of 1401R (100 ng/μL), 3.7 μL of 10× Taq buffer (5% 40 mM deoxynucleoside triphosphates, 40% 25 mM MgCl2, 50% 10× PCR buffer, 5% water), 18.8 μL of water, 0.2 μL of Taq polymerase (10 U/μL), and 0.3 μL of template DNA. The reactions were subjected to the following temperature cycling: 94°C for 8 min, followed by 20 cycles of 94°C for 1 min, 61°C for 1 min, and 72°C for 2 min, followed by 61°C for 1 min, and finally 72°C for 10 min.
The PCR products were purified and sequenced directly using ABI PRISM Big Dye Terminator Cycle Sequencing Kit (Perkin-Elmer) on an ABI 3100DNA sequencer, according to the manufacturer's instructions. The resulting 16S rDNA sequences were compared to GenBank databases using BLASTN (Palleroni, 2005).
Biochemical oxygen demand—BTE as only carbon source
In order to evaluate the biodegradation rate inhibited by the unavailability of oxygen, an experiment based on the Biochemical Oxygen Demand (BOD) system was carried out. The bottles containing enough amount of oxygen and using only BTE as a carbon source were incubated in a completely closed system with a sensor (avoiding changes in external atmosphere pressure that could interfere with the experimental data).
A 43.5 mL mixture containing BTE (50 mg/L of each compound), pelleted culture of microorganism suspension (absorbance concentration 0.8 in λ600 nm), and 1 mL of each nutrient buffer solution (phosphate buffer solution of 1.5 M, ammonium chloride solution of 0.71 M, calcium chloride solution of 0.25 M, magnesium sulfate solution of 0.41 M, and ferric chloride solution of 0.018 M) was added into proper amber glass bottles (500 mL) equipped with a CO2 trap system and closed with BOD measuring heads (OxiTop®-C controlled by the OxiTop OC110 system, Germany). The bottles remained in a closed system, thereby avoiding changes in external atmospheric pressure that could interfere with the experimental data. The liquid was kept under constant shaking at 20°C for 15 days. This protocol was first described by Mazzeo et al. (2010) where the BOD assay is able to quantify the pressure drop within the sealed bottles. The pressure within the system decreases because the available oxygen is consumed by the microorganisms, being expressed as mg/L. The CO2 produced by the microorganisms through oxidation of organic matter presents no effects on the pressure measurement, since it is removed by the sodium hydroxide crystals present in the bottle lids. A bottle containing only the microorganism culture and the nutrient buffer was also prepared to serve as a biodegradation control (control sample). Duplicate testing was performed.
Biodegradation assays with BTE
Biodegradation assays with BTE were prepared using bacterial cells precultured in the LB medium (1,000 mL of deionized water, 10 g of peptone, 10 g of NaCl, and 5 g of yeast extract) containing 10 mg/L of BTE. The cell pellets were washed twice with 0.85% of saline solution and transferred to a 50-mL screw cap flask containing 40 mL of MSM, 50 mg/L of benzene, 50 mg/L of toluene, and 50 mg/L of ethylbenzene (totaling 150 mg/L of contaminant). A stock solution was aliquoted by the following procedure: 2 mL of each aliquot was separated at time zero in vials for high-performance liquid chromatography (HPLC) and growth analyses (Abs 600 nm). This procedure was important to avoid BTE losses to the atmosphere.
Cells were grown for 10 days at 30°C under shaking at 200 rpm. The remaining cells and BTE in the culture medium were determined by increasing OD by 600 nm and in the HPLC analysis as a function of time, respectively. The bacterial strains without BTE and uninoculated media enriched with BTE were used as control samples under similar conditions. All the experiments were carried out in duplicate. This biodegradation method was suggested by Gracioso et al. (2012).
Biodegradation assays with commercial gasoline (ethanol 24% [v/v])
To test the efficiency of the strains to biodegrade commercial gasoline (24% ethanol added [v/v]), bacterial cells from the BTE assay were harvested by centrifugation at 10,000 g for 20 min at 4°C in a Cientec high-speed centrifuge. Cell pellets were washed twice in 0.85% NaCl to discard the excess BTE attached to the cell pellets. They were then transferred to a 50-mL screw cap flask (to prevent loss of volatile compounds) containing 40 mL of MSM plus 10% commercial gasoline (v/v). Cells were grown for 21 days at 30°C under shaking at 200 rpm. The remaining cells and gasoline in the culture medium were determined by increasing OD by 600 nm and by gas chromatography–mass spectrometry (GC/MS) analysis as a function of time, respectively.
Hydrocarbon ring fission mechanism
The ring fission mechanism was revealed using the Rothera reaction. Cells grown on MSM agar plates supplemented with 1 mM phenol were suspended in a 0.02 M Tris buffer (pH 8). The suspension was toluenized, and 0.2-mL catechol (0.1 M) was added to each 2-mL aliquot of suspension. The samples were shaken for 1 h at 30 ± 0.1°C and then tested for the appearance of β-ketoadipate (indicating an ortho-cleavage) by the modified Rothera reaction (Michaud et al., 2004).
Analytical Methods
HPLC of BTE analysis
HPLC analysis was performed using a Shimadzu LC-MS 2010A analytical system equipped with an online degasser, autosampler module, a UV detector adjusted to 208 nm, and a Supelco C8 column. The analysis was carried out with 60:40 ratio of methanol/water (v/v) for 20 min at a flow rate of 1 mL/min, with the column oven set to 35°C (according to the column manufacturer's instructions). Samples of culture media from the BTE test were collected every day and subjected to chromatography. A linear calibration curve for BTE and quantification of the main metabolites (phenol and catechol) by HPLC was obtained.
GC/MS analysis of commercial gasoline
Gas chromatographic analysis of gasoline components was performed based on the previously reported methods (Wongsa et al., 2004). After the incubation period, 1 mL of the cultures was extracted with two 2 mL volumes of n-hexane as a solvent using separation funnels to remove cellular material. Uninoculated control was incubated in parallel to monitor abiotic losses of the substrate. Biodegradation of gasoline was monitored by quantitative gas chromatographic analysis by means of a GC Varian 450 equipped with a Mass Spectrometer Varian 240-MS in an SE-54 capillary column (25 m × 0.32 mm). Helium was used as the carrier gas (0.8 mL/min). The oven was programmed as follows: 50°C (4 min) increasing to 250°C (5 min) at a rate of 20°C/min and held at least for 20 min. Injector temperature was 250°C while detector temperature was 325°C. The degradation of gasoline as a whole was expressed as the percentage of gasoline degraded in relation to the amount of the remaining fractions in the appropriate abiotic control samples (external standard technique). Combined areas under resolved peaks and the unresolved complex mixture were integrated to represent total gasoline compounds. The biodegradation efficiency (BE), based on the decrease in the total concentration of hydrocarbons, was evaluated using the following equation:
where AS, total area of peaks in each sample; Aac, total area of peaks in the appropriate abiotic control; and BE (%), efficiency of biodegradation.
The percentage of total BTE remaining after one test was calculated by comparing the area of their peaks with that of the corresponding peaks shown by a control that had been subjected to the same experimental conditions as the samples, except for the absence of bacterial culture.
Results and Discussion
Isolation and identification of BTE-degrading microorganisms
Two different strains (HCS1 and HCS2—Genbank number KF677012 and KF677013, respectively) were isolated from a BTE-contaminated soil sample. Both strains were able to degrade to 50 mg/L of BTE. Phylogenetic analysis based on 16S rRNA gene sequences indicated that these strains were members of Serratia sp. and Pseudomonas sp. genera, respectively (Fig. 1). The same results were obtained using the biochemical and mass spectrometry (MALDI-TOF) test. HCS1 was sensitive to trimethoprim, gentamicin, nalidixic acid, and ciprofloxacin and resistant to ampicillin, nitrofurantoin, and cephalexin, and the HCS2 obtained positive results to ADH (dihydrolase arginine), URE (urease), GEL (protease), GLU (glucose uptake), MAN (mannitol assimilation), NAG (assimilation of N-acetyl-glucosamine), GNT, CAP, ADI, and OX (data not shown).

Phylogenetic analysis based on 16S rDNA sequences of BTE-degrading strains HCS1 and HCS2. Neighbor-joining model is used for tree construction. Bootstrap values are indicated as percentages at the nodes. BTE, benzene, toluene, and ethylbenzene.
BOD—BTE as sole carbon source
To evaluate biodegradation by BOD, O2 consumption was continuously measured during the entire experimental period. This process, carried out in a BOD Trak Apparatus, took 15 consecutive days up to the end of the experiments. Based on oxygen consumption, we could infer that the microorganisms consumed the organic compounds present (BTE). Through this analysis, it was possible to estimate the instant that O2 consumption began to decline, indicating the moment the experiment was interrupted. In this study, this period consisted of 15 days. The highest indexes of O2 consumption were observed after the first 5 days of the degradation experiment. After this period, there was an increase in oxygen consumption (Fig. 2). This increased consumption observed in the second phase may be a consequence of cellular growth in the sample. A prolonged lag phase of 96 h (Fig. 3) was observed in an experiment containing BTE as the sole carbon source, followed by an exponential biodegradation phase, in which the growth biomass and simultaneous transformation of BTE occurred, and after 10 days, the curves showed a decline as a result of the total substrate consumption. This characteristic behavior can be explained by the increase in biomass and by self-inhibition. Other studies also report a prolonged lag phase, for most of the enrichment cultures studied by Vogt et al. (2011); long lag phases (sometimes more than 100 days) were observed before anaerobic benzene degradation. Once the lag phase had been completed, significant increases in cell mass occurred.
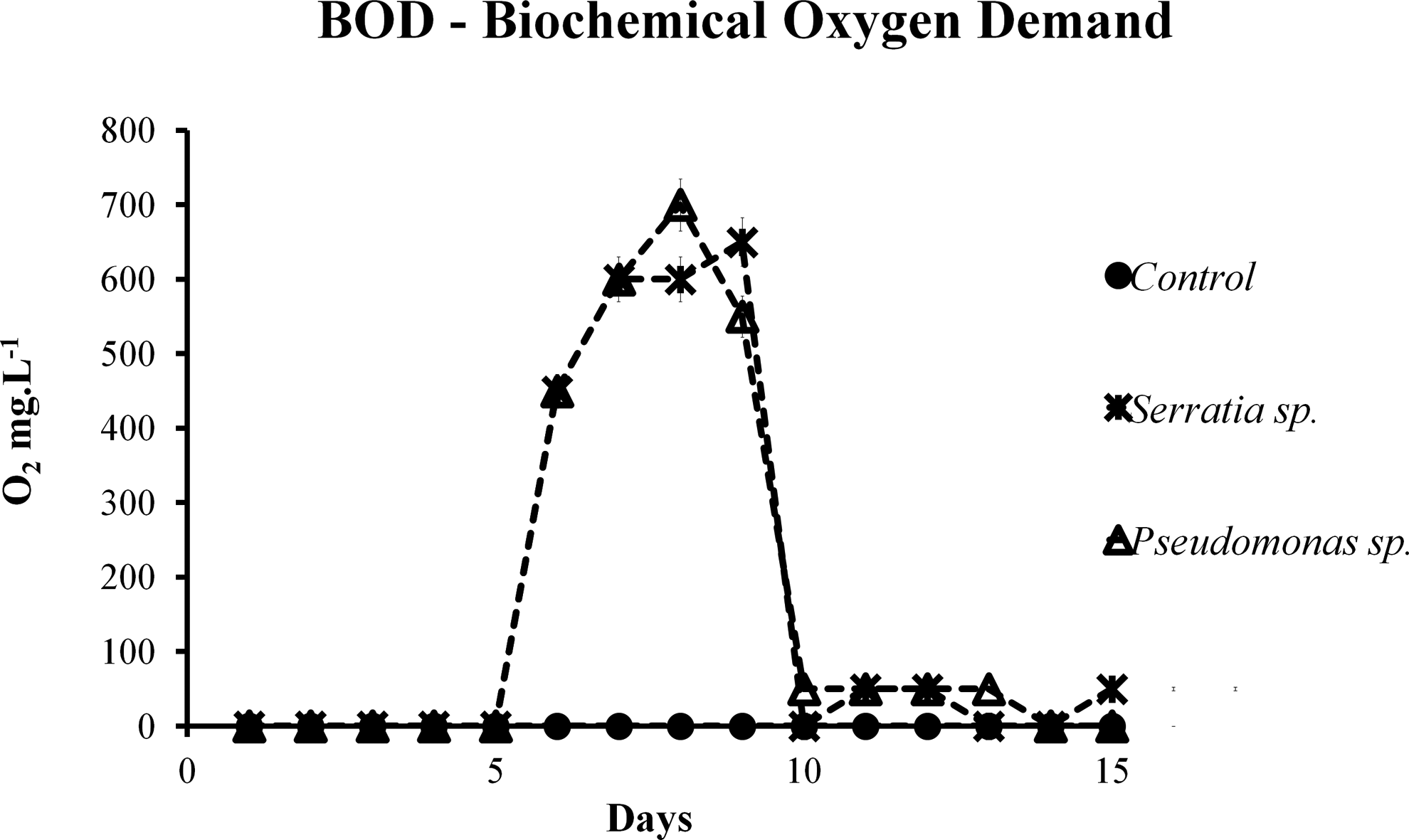
Mean values of O2 consumption in BTE concentration (50 mg/L) by bacteria in solution during 15 experimental days.
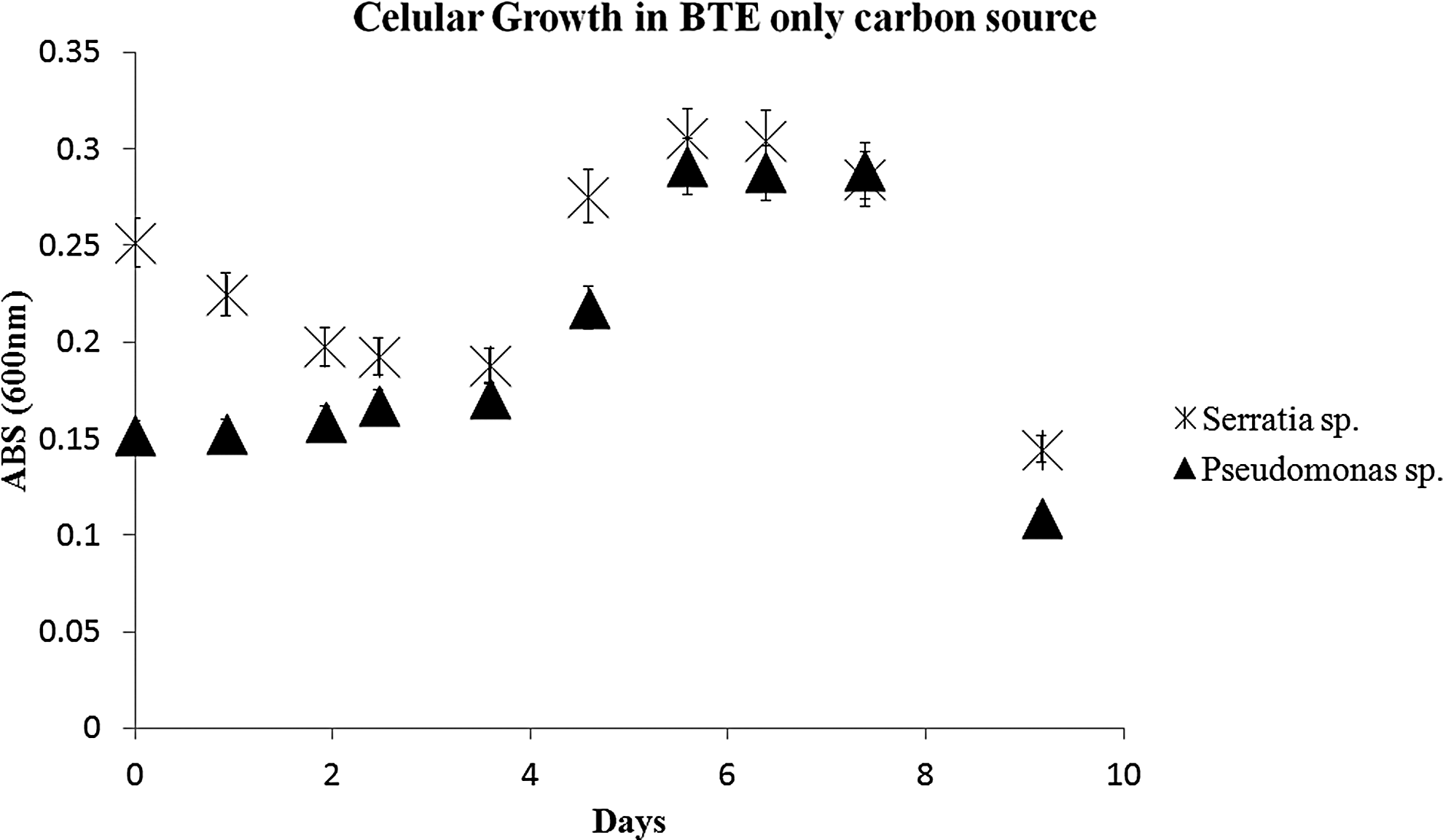
HCS1 and HCS2 growth in BTE (50 mg/L) as only carbon source.
The control samples had no available carbon source, so the O2 consumption was not that high, thereby probably indicating that the BTE present in other samples were important to bacteria growth. After 15 days, the samples were analyzed by GC/MS to quantify the amount of residual BTE. Figure 4 shows that all the BTE were consumed by Serratia sp. HCS1 and Pseudomonas sp. HCS2.

GC/MS profiles of BTE extracted from the aqueous phase of the medium after 15 days of incubation in a BOD system containing 50 mg/L of BTE with and without inoculation with strains Serratia sp. (HCS1) or Pseudomonas sp. (HCS2)
Biodegradation assays
BTE biodegradation assays performed on the tested samples are shown in Fig. 5. Chemical analysis after biodegradation showed that BTE amounts decreased in the test concentration (50 mg/L). The biodegradation assay was performed in a fully closed system and a control containing only BTE, and the minimum medium was used for confirming that the reduction in BTE concentrations was exclusively caused by consumption of the sole carbon source for the selected bacterial pool.
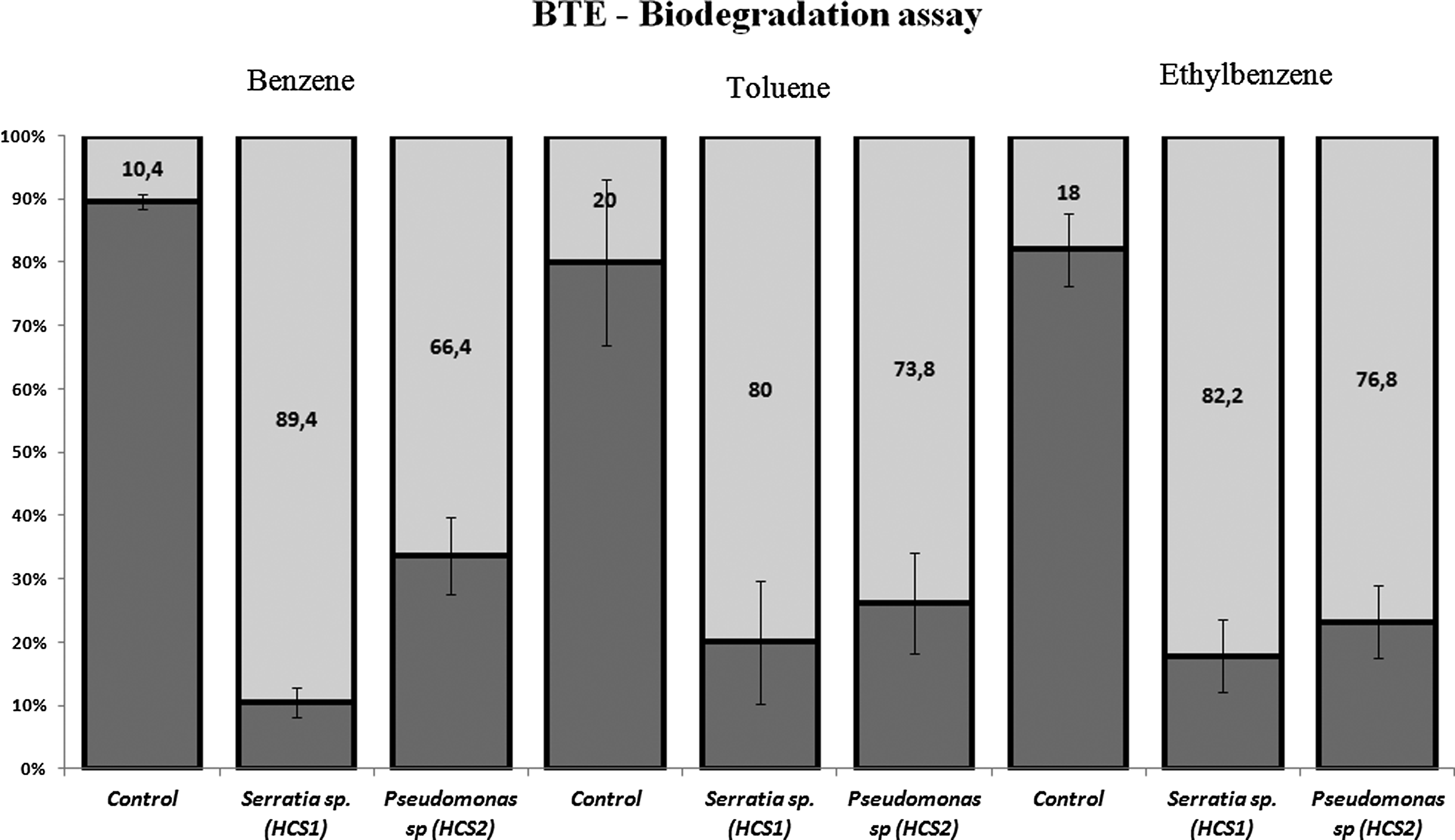
BTE consumption by bacteria after 10 days of incubation.
With regard to biodegradation kinetics for strains grown on BTE as the sole carbon source, in general, cells continued to grow until all substrates were consumed, but in this case, 10 days into the experiment we noticed cell death by the decrease in absorbance at 600 nm (Fig. 3). The incomplete BTE biodegradation could be attributed to oxygen depletion in the culture medium. Bacteria used in the experiment were aerobic and oxygen was required for complete BTE mineralization. Possibly, in this assay, the total amount of oxygen in the flask (2 mL of sample in a 2-mL flask, no air included) was not enough for the complete biodegradation of BTE, since biodegradation was complete in the BOD assay (bottle) where the bottle had a large volume of air (456.5 mL of air) (Shim et al., 2005).
Assays were done with BTE as soil carbon sources because biodegradation is affected by xylenes, which are toxic to unacclimatized microorganisms, as reported in some studies. Dou et al. (2008) and Lee and Kim (2011) reported that BTE were degraded when these compounds were provided together; however, xylene isomers persisted during degradation by P. putida E41. This fact could be particularly important since xylenes (ortho, meta, and para) comprise almost 10% of commercial gasoline composition, and in the event of an accidental gasoline leak, only bacteria extremely adapted to these compounds would be able to survive and promote the bioremediation process.
To check the ability of the isolated bacteria in biodegraded commercial gasoline, we performed an assay with 10% commercial gasoline as the substrate in MSM. In this assay, we demonstrated that the isolated bacteria were able to degrade about 60% (Serratia sp. HCS1) and 83% (Pseudomonas sp. HCS2) of the total of compounds within 21 days (Fig. 6 and Table 1). Serratia sp. HCS1 and Pseudomonas sp. HCS2 fully degraded benzene, toluene, p- and o-xylenes and partially degraded phenol (56% HCS1 and 73% HCS2), 2-methylphenols (49.5% HCS1 and 69% HCS2), and 4-methylphenols (54.7% HCS1 and 71% HCS2) besides the other unidentified components of commercial gasoline.
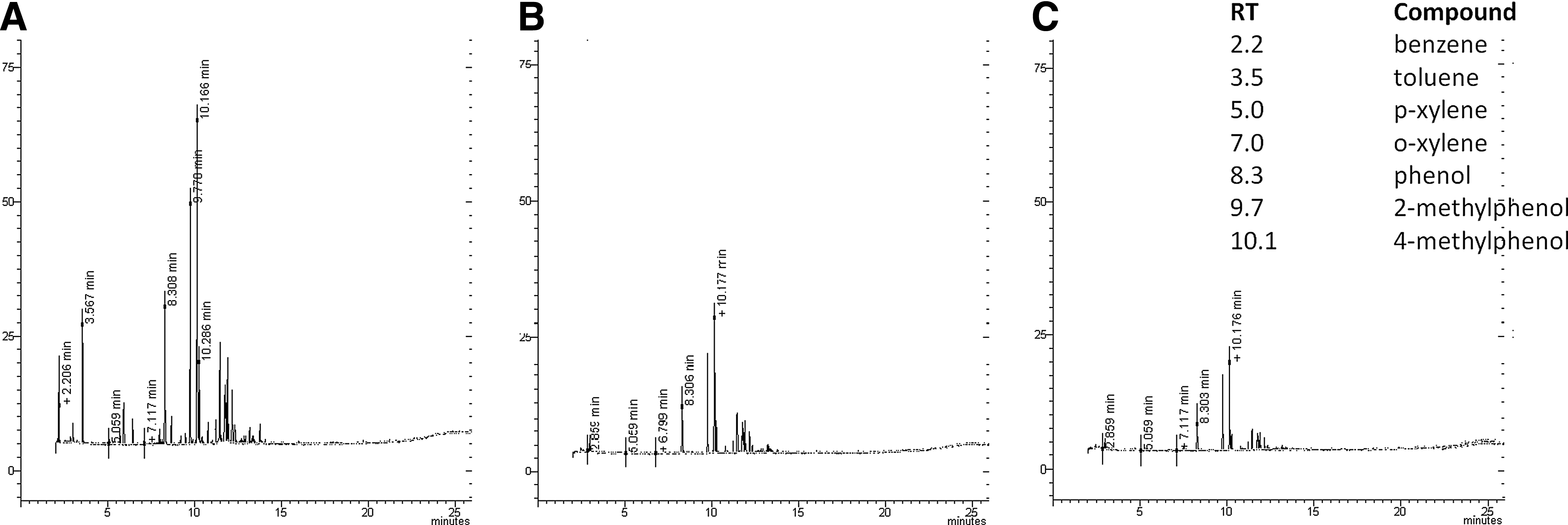
GC/MS profiles of gasoline extracted from the aqueous phase of the MSM after 21 days of incubation with 10% of gasoline (v/v) with and without inoculation with strains Serratia sp. (HCS1) and Pseudomonas sp. (HCS2).
GC/MS, gas chromatography–mass spectrometry; MSM, minimal salt medium; SD, standard deviation.
These results suggest that all the identifiable compounds were biodegraded, probably resulting in CO2 liberation, once no intermediates were detected in GC/MS analysis. These results also corroborate with the previous experiment of these bacteria with BTE compounds (Fig. 4), where no other intermediate aromatics were found.
HPLC and GC/MS analysis of BTE and commercial gasoline
The chemical analysis outcomes of BTE biodegradation (BOD assay) showed that the organic compounds were consumed within 15 days, and no metabolites were formed (Fig. 4). Both strains, HCS1 and HCS2, were able to mineralize all the compounds.
In Brazil, the ANP (National Petroleum Agency) establishes a limit of 1% for benzene and a combined limit of 45% for toluene, ethylbenzene, and xylenes in commercial gasoline. In this study, we investigated lower concentrations of the individual hydrocarbons (50 mg/L of each BTE in the biodegradation experiment), but the concentration of available aromatics in the aqueous phase of gasoline for microorganisms remained the same, because the aromatics were typically at the ppm level in the aqueous gasoline phase, as well as in ethanol. However, in these flask experiments, we did not observe total mineralization of the compounds (as we observed in the BOD results). The only difference between these experiments was flask size, which in the DBO assays contained more oxygen available for microorganisms. Accordingly, we concluded that available oxygen was the limiting factor for the biodegradation of these compounds.
In the presence of commercial gasoline as a substrate (10% in MSM), 60% and 80% of the HCS1 (Serratia sp.) and HCS2 (Pseudomonas sp.) strains, respectively, biodegraded out of all the compounds, including ethanol, within 21 days (as described previously, Brazilian commercial gasoline contains 24% ethanol). Previous studies, such as those by Alvarez and Hunt (2002), reported that the preferential degradation of fuel alcohols by microorganisms and the accompanying depletion of oxygen and other electron acceptors suggest that ethanol could hinder aerobic bioremediation of BTE. Alternatively, ethanol represents a carbon and energy source that is likely to stimulate the growth of a variety of aerobic and anaerobic microbial populations, including those that can degrade BTE compounds (Alvarez and Hunt, 2002).
Prince et al. (2007) described the primary aerobic biodegradation of regular gasoline by unacclimatized inocula within 7 days, but at a lower concentration (0.01% of gasoline in the Bushnell Haas medium). Biodegradation of individual hydrocarbons in gasoline follows a relatively consistent pattern: the larger n-alkanes and isoalkanes and simple and alkylated aromatic compounds were the fastest degrading compounds, followed by the smaller n-alkanes, isoalkanes, and naphthenes.
The last compounds to be degraded were butane, isobutane, and 2,2-dimethylbutane, but even these disappeared with an apparent half-life of <30 days. The fact that the aqueous concentration of many of the individual components was at the sub-ppb level could explain the ability of the aerobic microorganisms to respond to, and effectively biodegrade, gasoline-range hydrocarbons. In the gasoline biodegradation assay in this study, we identified the main aromatic compounds: benzene, toluene, p- and o-xylene, phenol, 2- and 4-methylphenol. All of them were degraded and there was a preference for biodegradation according to Prince's pattern: toluene/p-xylene/o-xylene>benzene>phenol>4-methylphenol>2-methylphenol (Table 1). The biodegradation of individual hydrocarbons in gasoline followed a relatively consistent pattern, which is the same in Prince et al. (2007).
Hydrocarbon ring fission mechanism
In tests for detecting the cleavage of catechols, we observed that the bacteria studied used only ortho-fission of catechol. The results were positive for Rothera's test, characterized by a purple color after the Rothera reaction, which indicates the presence of β-ketoadipate.
Most aromatic compounds are first converted into one dihydroxylated substrate (catechol), whose aromatic ring can be enzymatically cleaved (ortho- or meta-cleavage). In the β-ketoadipate pathway, a primarily chromosomally encoded catabolic route that is widely distributed in soil bacteria and fungi, catechol is cleaved between their two hydroxyl groups by catechol 1,2-dioxygenase or catechol 2,3-dioxygenase. This pathway converges on the ring cleavage products from catechol to β-ketoadipate, the metabolite for which the pathway was named. Two additional steps complete the conversion of β-ketoadipate to tricarboxylic cycle intermediates.
Paul et al. (2004) described only the ortho-fission by two strains of bacteria, Arthrobacter protophormiae RKJ100 and Burkholderia cepacia RKJ200, in pesticide degradation. Arenghi et al. (2001) reported the mechanism regulation of meta-cleavage by Pseudomonas stutzeri OX1. In contrast, Heinaru et al. (2000) reported that meta- and ortho-pathways are carried in the same bacterial strain in many cases. Many phenol-degrading bacteria harbored two types of catabolism pathways. Although the best known phenol degrading pathway is meta-cleavage, bacterial strains are capable of performing the two-way metabolic pathway, depending on the available substrate for growth.
Conclusions
From the results obtained, we observed that HSC1 and HSC2 are effective in biodegrading hydrocarbons, as well as gasoline. At the end of BTE experiments, small concentrations of hydrocarbons remained and the main reason could be the amount of oxygen in the experiment. In this case, it was insufficient for the total elimination of the high-concentration BTE present, implying the need for an oxygen source to achieve effectiveness in this process. This information is consistent with the results of the BOD assay, when all the BTE was consumed after 15 days, and where the important factor to be considered in the biodegradation process is the amount of available oxygen in the sample, which should correspond to the physiological needs of the degrading microorganism. The results in different conditions of oxygen availability (flasks × bottle) corroborated this theory.
In the Brazilian Commercial gasoline experiment, the entire contaminant was degraded 60% (v/v) and 80% (v/v) after 21 days by HCS1 and HCS2, respectively. These results suggest that these strains have a high level of tolerance to hydrocarbon toxicity and, consequently, have potential application in the biotreatment of industrial wastewater or in situ bioremediation of hydrocarbon-contaminated soils.
In the second phase of biodegradation, the results suggest that these two strains use the ortho-fission of catechol metabolism during hydrocarbon biodegradation and, subsequently, the contaminant is easily used as an energy source for bacteria.
Results also indicate that, under the appropriate conditions, the bacteria Serratia sp. HCS1 and Pseudomonas sp. HCS2 are efficient microorganisms for BTE and gasoline biodegradation and suitable for the biotreatment of industrial wastewater or remediation of environments contaminated with these compounds.
Footnotes
Acknowledgments
The authors thank Marinalva Martins-Pinheiro for helpful discussions. This study was supported by the National Institute of Science and Technology—Environmental Studies (INCT-EMA/Brazil), Fapesp and CNPq.
Author Disclosure Statement
No competing financial interests exist.