
Abstract
Abstract
Atlantic Forestry is one of the most important biomes that has been reduced by deforestation. Plants as Senna multijuga and Erythrina crista-galli are important for this ecosystem. High concentrations of copper in the soil can be toxic to plants and other organisms. There are many situations of copper-contaminated areas that need intervention for recovering the natural and or a sustainable condition of the environment. This study is a preliminary study in a wide project, studying different species and conditions for reclamation of areas contaminated with copper. The study aimed to evaluate the initial growth, tolerance, and copper phytoremediation of S. multijuga and E. crista-galli, both native trees from Atlantic forestry. The seedling is the most important phase in the growth of species in the field. This study was conducted for 120 days in a greenhouse experiment, using a completely randomized design in a factorial arrangement (2 × 6). Plant height, stem diameter, dry weight of the roots and the shoots, the cumulative amount of copper in the roots and the shoots, Dickson quality index, tolerance index, translocation index, and bioaccumulation factor were measured. Results indicated that doses of copper reduced morphological parameters of the plants and it may reduce the growth and stabilization of the plants in the following steps. Copper uptake by plants was predominantly accumulated in the root system, with a low translocation toward shoots. E. crista-galli and S. multijuga did not demonstrate high tolerance to the highest copper concentrations in this study; however, they had tolerance to this metal and can be used for reclamation of copper-contaminated areas.
Introduction
I
An alternative for recovering some areas contaminated with copper is the phytoremediation, which is the use of plants for extracting and/or reducing the toxicity of pollutants to the organisms and to the environment (Chigbo et al., 2015; Zaheer et al., 2015). There are many ways for reducing the bioavailability of Cu(II) by plants such as phytoextraction, phytoaccumulation, phytoadsorption, and phytostabilization (Kavamura and Esposito, 2010). All these techniques reduce the availability in the environment by binding the metals in the roots and/or in the phytomass. According Kabata-Pendias (2011), the copper is found with more concentrations in the roots of the plants because Cu(II) is tightly bound to the cell walls of the roots and it is not readily mobile into the plant, resulting in a low metal translocation to the shoots. This behavior is a system for resistance of heavy metals used by many plants and organisms to reduce translocation of heavy metal to the upper plant, and thus prevent damage caused by toxicity of the heavy metal.
The phytoremediation has become viable as an alternative for chemical-contaminated environments because it is considered an eco-friendly low-cost technique and promotes the maintenance of fertility and stability of soil (Robinson et al., 2003). Some authors indicate the use of forestry tree species as an important strategy for recovering areas contaminated with heavy metals, as they are perennial and have large biomass production (Domínguez et al., 2009; Jensen et al., 2009). Tree species that accumulate heavy metals in the roots and stems are important to restore contaminated environments (Domínguez et al., 2009; Jensen et al., 2009). In addition, the absorption of water by the trees reduce the overall flow of water through the soil, thus, it helps to reduce the amounts of heavy metals that are lixiviate to the ground water and surface water courses (Pulford and Watson, 2003). So, the use of the trees may be an important role for phytoremediation, reinforcing the necessity of more studies on species with the capacity for Cu(II) binding and reducing the bioavailability of Cu(II) for the environment and consequently reducing the contamination of the water courses.
The native trees and the Fabaceae family plants can be an interesting alternative to recover copper-impacted areas. It is due to their capacity for nitrogen incorporation, organic matter deposition, and improvement of physical properties of the soil (Manhães et al., 2007). The Senna multijuga ([Rich.] H.S. Irwin and Barneby) and the Erythrina crista-galli are both native species from Atlantic Forestry, and both are pioneer species representatives from this family that have fast initial growth to moderate growth (Lorenzi, 2008), between 2 and 12 months, which promote the use of these species in plantations for recovery of contaminated areas. However, there is a lack of studies on the behavior of these species in soil contaminated with Cu(II). In this context, the design and implementation of studies that evaluate the growth and resistance of these species in the soil contaminated with copper may contribute to the recommendation of native tree for reforestation in soil contaminated with copper. So, the aim of this study was to evaluate the initial growth, tolerance, and Cu(II) phytoaccumulation in the seedlings of the S. multijuga and the E. crista-galli.
Experimental Protocols
The experiment was carried out in greenhouse condition (with 12 h of light and 12 h of dark) and 80% of relative humidity of air and temperature of 20°C at night and 28°C at day. The soil used for the experiment was sampled from a collection bank of soils from the Federal University of Santa Maria, at Frederico Westphalen, Brazil, and characterized as Oxisol, whose the physicochemical analysis were carried out according to the methodology described by Mann and Ritchie (1993) for the soluble Cu(II) (extractor KCl 0.005 mol/L), USEPA (1996) to Cu(II) pseudo-total (3050B), and Tedesco et al. (1995) for pHágua, Ca, Mg, H, Al, P, K, MO, and clay (Table 1).
The pH of the soil was determined in water (1:1).
Extrator KCl 1 mol/L.
Extrator Mehlich-1.
H+Al determined by SMP index; OM, sulfochromic solution with exterior hit; clay determined with hydrometer after soil dispersion in sodium hydroxyl solution.
The seeds of the trees used were of the species of S. multijuga (Rich.) H.S. Irwin and Barneby and E. crista-galli L., both seeds provided by the Center of the Forest Research of the State Foundation for Agricultural Research (FEPAGRO), at Santa Maria, Rio Grande do Sul, Brazil. To overcome the dormancy of the seeds, the S. multijuga and E. crista-galli seeds were immersed for 15 and 30 min. in a solution of concentrated sulfuric acid (ACS standard, 95–98% v/v), respectively. Subsequently, they were washed in running water for 1 min. Seeds were sown in seedbeds and when presented a pair of true leaves they were transplanted into plastic bags of polyethylene with volumetric capacity of 600 cm3.
The experimental design was completely randomized in a factorial arrangement (2 × 6), with two tree species (S. multijuga and E. crista-galli) and five doses of Cu(II) added to the soil [60, 120, 180, 240, and 300 mg/kg of Cu(II)] and a control treatment (natural soil) with six replications of each treatment. Each experimental unit contained one seedling. Previously, Cu(II) doses were applied into the soil 30 days before transplanting in the form of Cu(II) sulfate (CuSO4·5H2O), and diluted with 50 mL of water to allow homogenization by sparkling into the soil in the plastic bags.
The experiment was conducted for 120 days after transplanting the seedlings. The plants were daily irrigated with distilled water based on the weighting of experimental units, maintaining the humidity of the soil to about 80% of the field capacity. Fertilization was performed in two phases: at the first phase of the experiment applied the equivalent of 150 g of N/m3 of soil (as urea form), 700 g of P2O5/m3 of soil (as triple superphosphate), and 100 g of K2O (as potassium chloride) per m3 of soil; and the second phase was applied three times as nutrient solution with nitrogen (2.0 g/L) and potassium (1.5 g/L) and applied three times at 30, 60,. and 90 days after transplanting, following the recommendations by Gonçalves and Benedetti (2005). To meet the requirements of experimental design, a weekly rotation of experimental units was conducted.
At the end of the experiment, the shoots height (H) with a graduated scale from the base of the seedlings to the top of the shoots was measured; stem diameter was measured with a digital caliper with a precision of 0.01 mm. To determine the dry mass of the roots (DMR) and dry mass of the shoots (DMS) both fractions were separated in the cervical region of the plant. The roots were washed with water until all the soil particles were removed and after that washed again with distilled water. Subsequently, the material was dried in an oven with forced-air at 60°C ± 1°C. The total dry matter (TDM) was determined by the sum of the DMR and DMS. As per the methodology described by Tennant (1975), the specific surface area (SSA) of the roots was estimated; and through the Equation (1), the Dickson quality index (DQI) (Dickson et al., 1960) was determined.
After weighting the DMR and DMS, the plant mass was ground in a mill type Wiley (10 mesh of sieve) for the determination of Cu(II) content in plant tissues, through digestion with nitric perchloric acid (3:1) (Van Schouwvenberg and Walinge, 1973) and the determination was made with the atomic absorption spectrophotometry, with apparatus of Shimadzu, model: AA6200 having 0.04 ppb detection limits for copper.
Based on the TDM, the Cu(II) content (mg/kg) of the roots (CuR) and shoots (CuS) were analyzed, the accumulated amounts of Cu(II) (μg/plant) in the roots system (CuAR), shoots (CuAS), and total copper in the seedlings (CuAT) in the control treatment (d0) and in the amended doses from 60 to 300 mg/kg (dn) were also analyzed. The tolerance index (Toi) was calculated according to Equation (2) (Wilkins, 1978) and the translocation index (Tri) using Equation (3), which it is the total percentage of Cu(II) absorbed and transported to the shoots (Abichequer and Bohnen, 1998).
The bioconcentration factor (BCF) of copper was calculated as follows (Yoon et al., 2006; Shi and Cai, 2009):
where [Cu]shoot and root is the concentration of copper in the shoots plus roots, and the [Cu]soil is the concentration of copper in the soil.
The metal extraction ratio (MER) is defined as the ratio of metal accumulation in the shoots to that in soil (Mertens et al., 2005), which was calculated as follows:
where Cplant is the concentration of metal in the harvested component of the plant biomass, Mplant is the mass of the harvestable aboveground biomass (shoots) produced in one harvest, Csoil is the concentration of metal in the soil volume, and Mroot zone is the mass of the soil volume rooted by the species under study.
The plant effective number for the shoots (PENs) and the plant effective number for the total plant (PENt) were applied to evaluate the ability of a hyperaccumulator plant to rehabilitate contaminated soil (Sun et al., 2008).
where [Cu]shoots is the concentration of the Cu in the shoots, Mshoots is the total dry mass produced after 57 days of plant growth, [Cu]entire plant is the Cu concentration in the shoots and roots, and the Mentire plant is the DMS and DMR.
The results analyzed the variance and when presented significant interaction between the quantitative factors, the regression analysis was plotted. For parameters without significant interaction, simple effects were plotted and the means of qualitative factor compared by Tukey test at 5% of error probability and the average quantitative factor were submitted to the polynomial regression analysis by SISVAR program (Rocha et al., 2012).
Results and Discussion
Doses of copper and plant's growth
The doses of Cu(II) affected the growth of both plants and also the pseudo-total copper in the soil (Figs. 1 and 2). Also, the highest quality of seedlings (amended dose of 150 mg of Cu/kg of soil) was showed from the estimated concentration of 342.35 mg of Cu/kg of soil (Fig. 1), higher concentration than the value standardized to agricultural soils of Brazil by CONAMA (2009) and that caused a reduction in other morphological parameters such as height and dry mass of the seedlings. The DQI showed the maximum pick with addition of 150 mg of Cu/kg of soil (Fig. 3C), the value showed for the E. crista-galli was significantly higher than that in the S. multijuga (Table 2). The value of the DQI will be better as much as high quality of the seedlings. Thus, Cu(II) concentrations amended higher than the 150 mg of Cu/kg of soil and caused a reduction in the quality of the both seedlings.
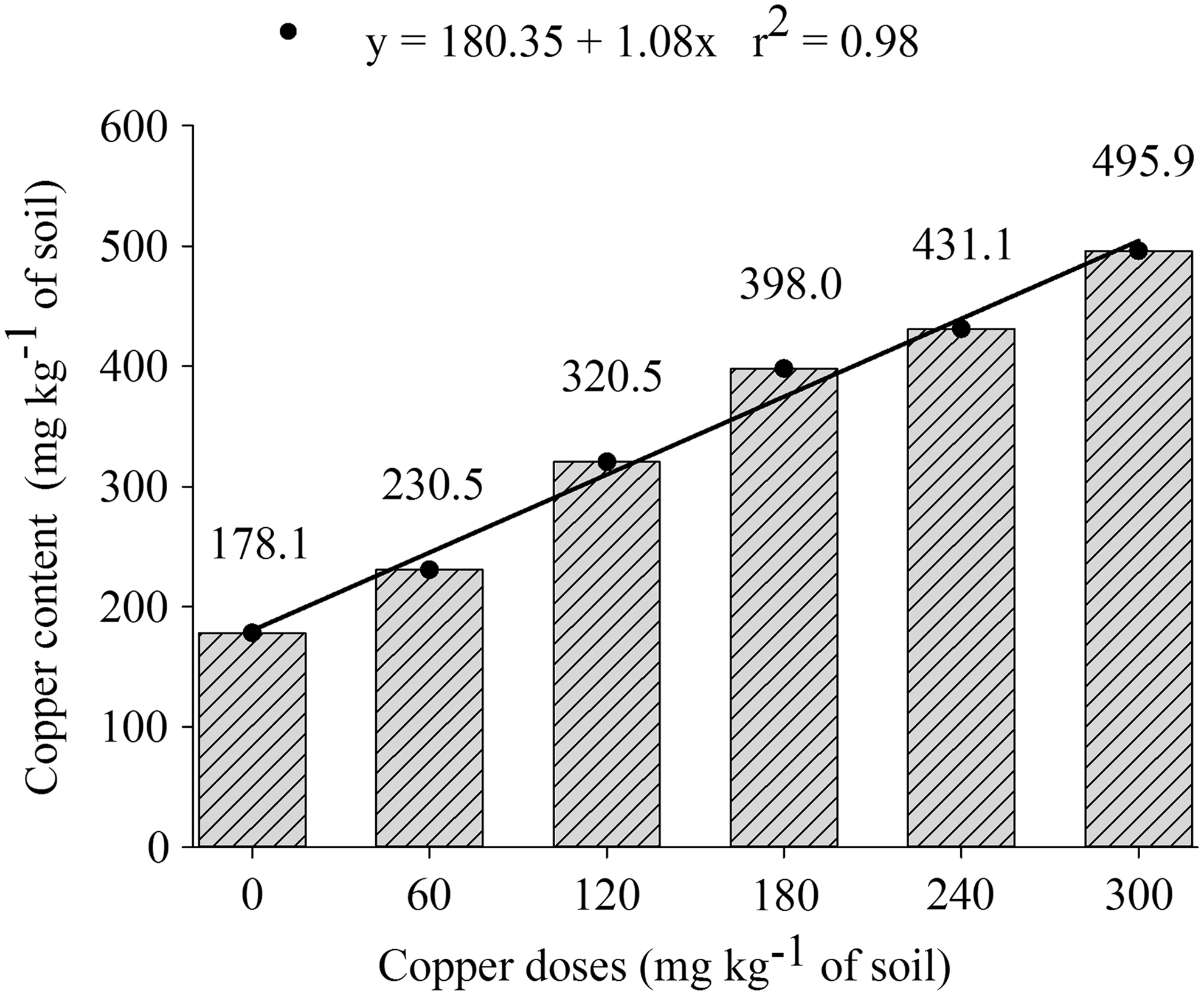
Initial contents of copper (pseudo-total) and copper doses added to soil.
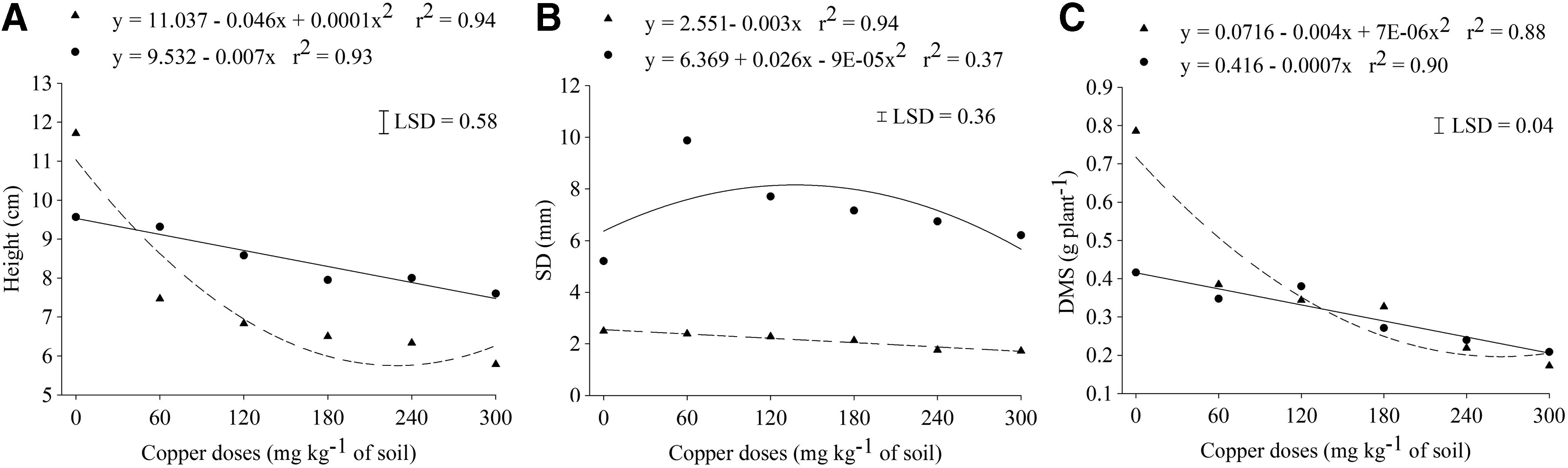
Regression analysis of plant's height
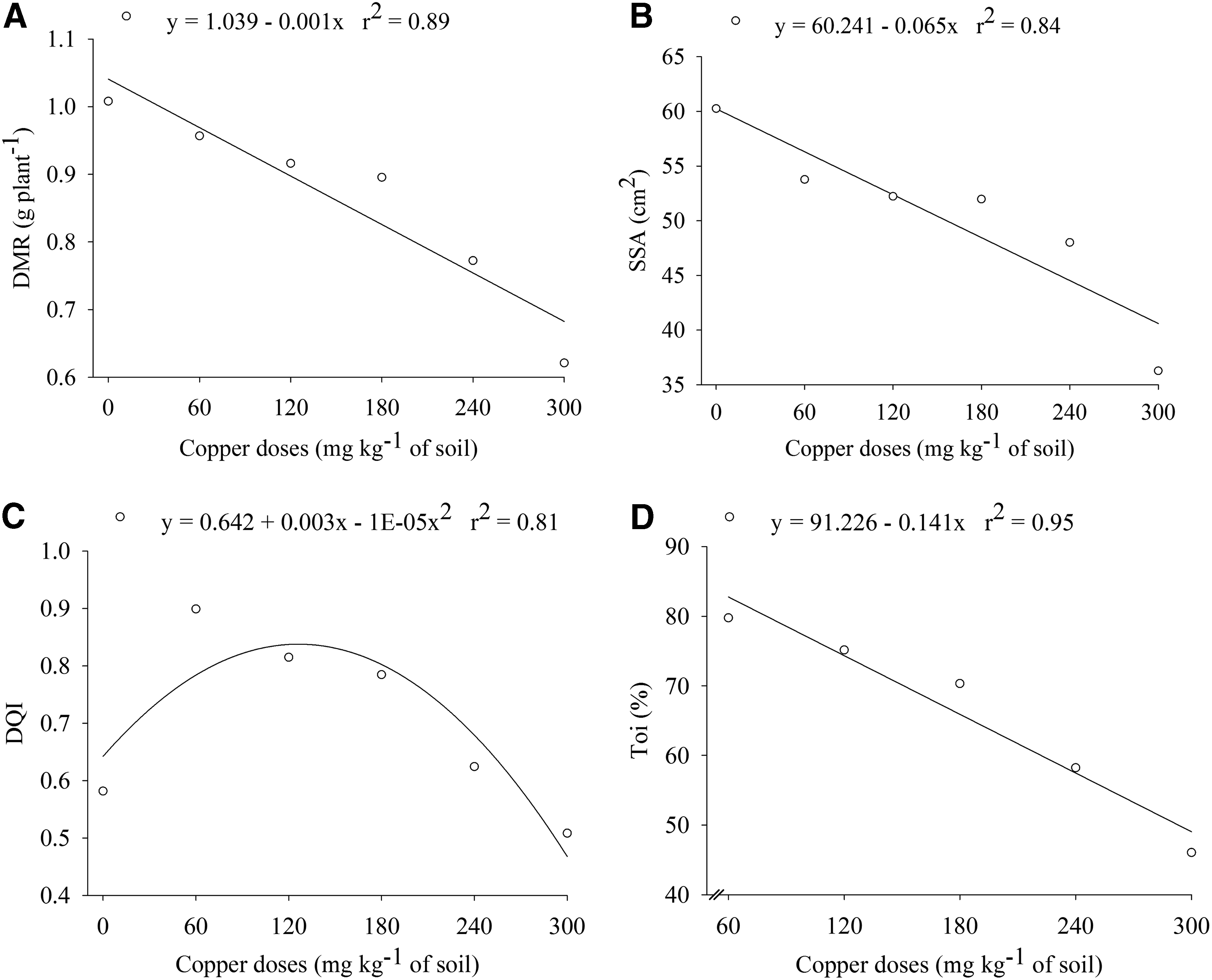
Regression analysis of the dry mass of roots
Averages with the same letters in the column are not significant by Tukey test (p > 0.05).
CV, coefficient of variation; DMR, dry mass of the roots; DQI, Dickson quality index; SSA, specific surface area; Toi, tolerance index.
Total Cu(II) concentration in the soil was above the toxic limit (200 mg/kg) reported by Resolution No. 420 for agricultural soils of Brazil (CONAMA, 2009). Therefore, serious problems for human health and environment can be noted when it crossed this limit (CONAMA, 2009). However, it should be noted that values of reference should be defined in each state, since the natural content of the Cu(II) in soils is widely variable and depends on the rock matrix and intensity of soil formation processes (physical, chemical, and biological properties) on the rock. Thus, more studies are necessary on soil quality in each state of Brazil and also in Rio Grande do Sul.
The analysis of variance showed a significant interaction (p ≤ 0.05) between both tree species and the doses of Cu(II) for height, stem diameter, and shoot dry mass (Fig. 2). Results showed that the doses of Cu(II) caused a linear decrease in the height of the seedlings of the E. crista-galli and a quadratic reduction in the S. multijuga with minimal growth when applied 230 mg of Cu/kg of soil (Fig. 2A). Cu(II) may cause disturbances in plant growth and development, affecting important physiological processes in plants (Kabata-Pendias, 2011). The excess of Cu(II) affected the growth of the roots and the shoots due to disorders in the structure of proteins and inhibition of cell elongation caused by permeability, decrease of the plasma membrane, and the cell wall lignification, presenting with symptoms of dwarfing plants (Yruela, 2009). It is possible that these physiological changes have occurred, resulting in the reduction of height of both the studied species with increment of the doses of Cu(II) added to the soil.
The doses of Cu(II) added to the soil separately changed the stem diameter of the both species (Fig. 2B). The stem diameter of the plants of the S. multijuga was linearly reduced with the increase of the doses of Cu(II); while the E. crista-galli was stimulated in the lower dose of Cu(II), with subsequent reduction when the dose was increased. According to Taiz and Zeiger (2009), the E. crista-galli response to Cu(II) may be due to its essentiality as a chemical element for plant growth, participating as a catalytic element for biochemical reactions in the metabolism of carbohydrates, nitrogen, in chlorophyll synthesis, and the production of protein in plants. However, at higher concentrations than those required for optimal growth of each plant, it can inhibit the growth and interfere with important cellular processes. Guo et al. (2010) showed that the Cu(II) toxicity may differ between plant species; and for Lequeux et al. (2010), this effect has been attributed to plants that develop adaptive mechanisms in response to metal toxicity. Regarding the control dose in the soil, the results of this study indicated that the other doses of Cu(II) used reduced the stem diameter only for the S. multijuga species.
The DMS of E. crista-galli seedlings linearly decreased with the increasing doses of Cu(II), whereas for S. multijuga there was a quadratic reduction with minimal pick at 286 mg Cu/kg of soil (Fig. 2C). Besides the reduction of dry mass, symptoms of chlorosis is usually observed in the shoots (Yruela, 2013) due to the substitution of the ions Mg2+ and Fe2+ in the molecule of chlorophyll by Cu2+, inducing breakdown in the photosynthesis process (Küpper et al., 1996; Kabata-Pendias, 2011). However, symptoms of chlorosis was not seen on seedlings of the both species used in this study.
Results did not exhibit interaction (p > 0.05) between tree species and Cu(II) levels applied in the soil to the roots biomass (DMR), SSA, DQI, and index tolerance (Table 2 and Fig. 3). The dry weight of the roots and SSA were reduced linearly with increasing of doses of Cu(II) in the soil for both native plants (Fig. 3A, B), and the E. crista-galli was statistically higher than the S. multijuga in this issue (Table 2). According to the estimate, there was 29% reduction in root biomass production at the dose of 300 mg of Cu/kg of soil, compared with the seedlings produced without Cu(II) amendment (Fig. 3A), whereas for the SSA, the reduction was 32% after this same dose of copper (Fig. 3B). According to Kukkola et al. (2000), inhibition of root growth is recognized as one of the most evident symptoms of Cu(II) toxicity. These symptoms expressed as the lack of roots formation and darkening of the roots, reducing the capacity of the absorption of water and essential nutrients in roots, consequently reducing the plant's growth (Panou-Filotheu et al., 2001; Taiz and Zeiger, 2009). This toxic effect can be observed in this study with a reduction in DMR and SSA at the dose of 60 mg of Cu/kg of soil.
The amended soil with the doses of Cu(II) reduced the tolerance of the plants; however, the E. crista-galli had shown significantly higher tolerance than the S. multijuga species (Fig. 3D and Table 2). The tolerance to Cu(II) toxicity has been proposed in the literature and include mechanisms such as enzyme activity in the detoxification of free radicals, Cu(II) chelating by organic compounds, and/or compartmentalization into the plant vacuole (Qian et al., 2005) allowing tolerant species to grow in the soil with toxic levels with many plants (Macnair et al., 2000). Thus, the results of this study indicate the possibility of E. crista-galli to have tolerance mechanisms for Cu(II), allowing greater growth of S. multijuga in Cu(II)-contaminated soil; however, both species reduced the total dry mass (Toi <100%) on the first Cu(II) dose in the soil (60 mg Cu/kg of soil).
Potential of native plants for Cu phytoremediation
Analysis of variance showed a significant interaction (p ≤ 0.05) for both the tree species and the doses of Cu(II) to the copper content of the roots—CuR (Fig. 4A) and the shoots—CuS (Fig. 4B), copper accumulation in the roots—CuAR (Fig. 4C) and in the shoots—CuAS (Fig. 4D), and in the translocation index—Tri (Fig. 4E). The doses of Cu(II) added to the soil substantial influenced the metal contents in the roots and the shoots (Fig. 4A, B). The Cu(II) content in the roots of S. multijuga increased with the increase of doses added to the soil, and the Cu(II) content in the shoots was reduced at Cu(II) estimated dose at 182 mg of Cu/kg of soil. In the E. crista-galli plants, the highest Cu(II) content in the roots and the shoots was shown at estimated doses of Cu(II) of 154 and 163 mg of Cu/kg of soil, respectively; with subsequent reduction when the Cu(II) doses are increased. The Cu(II) content in the roots and shoots of the S. multijuga seedlings was significantly higher than the E. crista-galli seedlings, showing values about 3 times higher in the roots and 1.4 times higher in the shoots for S. multijuga than the E. crista-galli. Studies with Cedrela fissilis Vell demonstrated that Cu(II) content in the roots were higher than the shoots (Caires et al., 2011). According to Hall (2002), the plasma membrane plays an important role in the homeostasis of heavy metals by preventing or reducing the input of metal in the cell, or to promote the output of the toxic ions. Taiz and Zeiger (2009) reported that the differences between plant species and the absorption of metals are associated with the particular tolerance of each species to the contaminant. Thus, it is possible that the selectivity of the plasma membrane of E. crista-galli represents a major mechanism for tolerance to Cu(II), resulting in low metal concentration compared to the S. multijuga. However, this hypothesis should be confirmed by other studies.
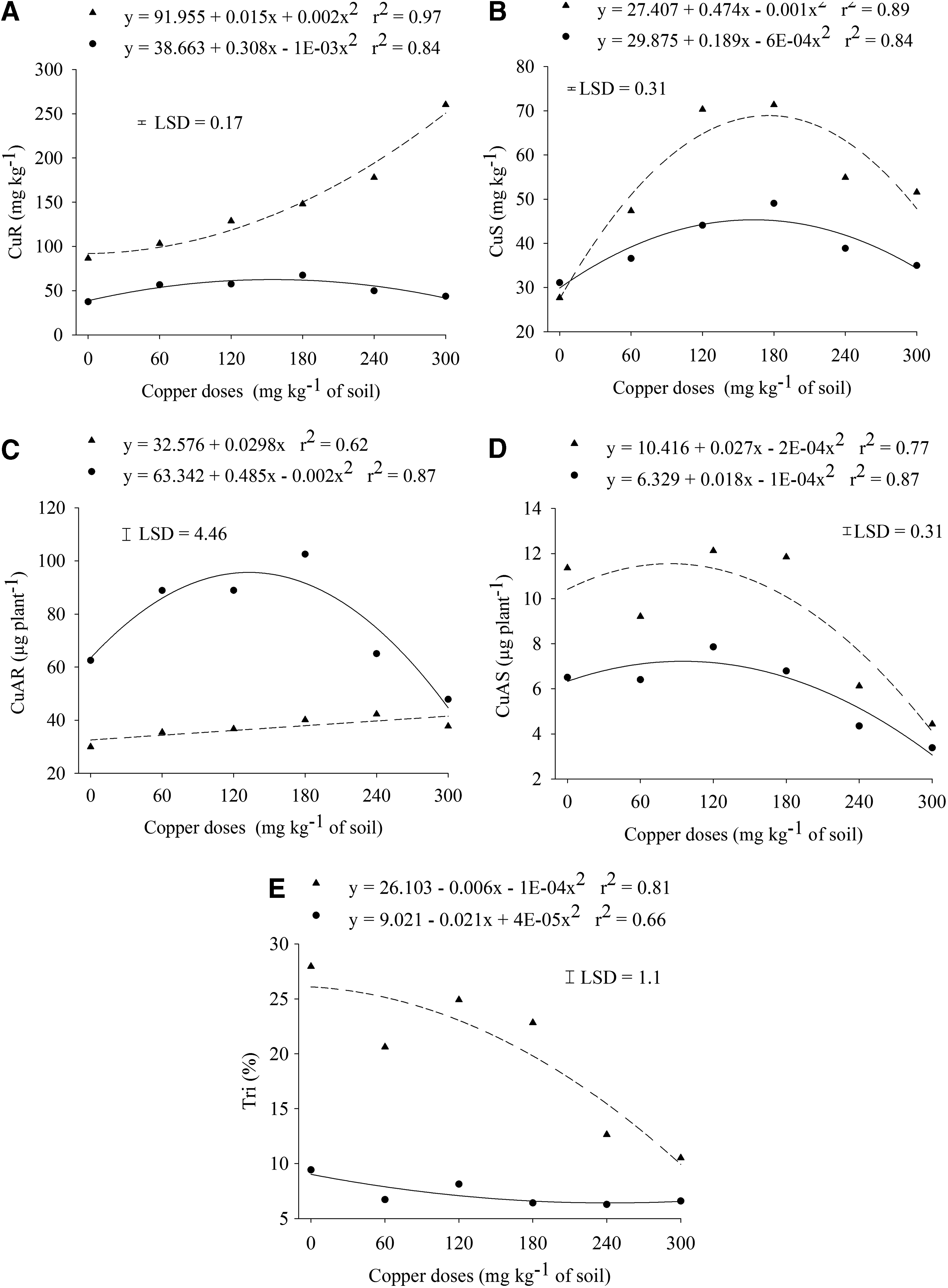
Regression analysis of copper content of roots
The results showed an increase in the accumulation of Cu(II) in the roots of the E. crista-gallii until the estimated dose of 121 mg of Cu/kg of soil, while for the S. multijuga the Cu(II) accumulated in the roots was linearly increased according to the high doses of Cu(II) (Fig. 4C). Therefore, the total amount of Cu(II) in the shoots was lower than the roots, with the maximum accumulation of Cu(II) on the estimated dose of 90 and 68 mg Cu/kg of soil for E. crista-gallii and S. multijuga, respectively (Fig. 4D). Plants with larger amounts of metals accumulated in the roots than the shoots are classified as species tolerant to metal absorption (Weis and Weis, 2004), it is an indication that the mechanism of tolerance of the species studied is to accumulate Cu(II) in the roots.
The doses of Cu(II) amended to the soil reduced the translocation index of the species under study (Fig. 4E). It was observed that less than 30% of the total absorbed Cu(II) was transported to the shoots of the S. multijuga and less than 10% in the E. crista-galli. Other studies have been shown same results in the reduction of translocation of Cu(II) from the roots to the shoots in three species of Brazilian native forestry plants such as Enterolobium contortisiliquum, Bauhinia forficata, and Pterogyne nitens (Silva et al., 2015). According to Pulford and Watson (2003), regulation of heavy metal uptake into the roots and the low translocation to the shoots are considered a tolerance mechanism with high levels of heavy metals in the environment.
The bioconcentration factor (BCF) was significantly higher in the species S. multijuga in all Cu(II) doses amended to the soil than E. crista-galli (Table 3). BCF is calculated with the potential of the plants bioaccumulate the metal according to that in the soil. E. crista-galli showed a major restriction to uptake Cu(II) even with the increment of the concentrations added in the soil. It contributes to E. crista-galli being more tolerant than the S. multijuga, and consequently, it showed a higher DMR (Table 2). Other studies demonstrated that both Plantago major and Bidens alba plants cultivated in copper-contaminated sites showed BCF values of 1.2 and 0.48, respectively (Yoon et al., 2006). In other indigenous species such as Acacia raddiana and Aerva javanica, the BCFs were almost 0.2 and 0.23 respectively in plants grown in mine tailings (Rashed, 2010). Compared with our results, our tree species can bioaccumulate more than those in the literature.
BCF, bioconcentration factor; MER, metal extraction ratio; PENs, plant effective number for the shoots; PENt, plant effective number for the total plant.
MER is an index that evaluates the ratio of metal accumulation in the shoots to the metal accumulation in the soil (Mertens et al., 2005). Our results indicated that both native plants cultivated in different Cu(II) amended soils had not that high MER index (Table 3) compared to other plants such as castor bean (Andreazza et al., 2013a) and Brachiaria decumbens (Andreazza et al., 2013b) cultivated in copper-contaminated areas. It demonstrates a nonphytoextraction characteristic for both native plants under study.
The plant effective number (PEN) is defined as the number of plants needed to take up 1.0 g of metal when the biomass of shoots and/or entire plant is considered (García et al., 2004). The native plants S. multijuga and E. crista-galli demonstrated a high number of PENs with values of 9,751 and 24,906, respectively, at treatment with amendment of 180 mg/kg of copper (Table 3). However, both native plants promoted PENt after 120 days of growth with values of 8,742 and 21,303 plants for S. multijuga and E. crista-galli, respectively, at treatment with amendment of 180 mg/kg of copper (Table 3). Other study with castor bean grown in vineyard soils contaminated with copper, presented results for of PENs and PENt of 31,136, and 187 plants after 57 days of growth (Andreazza et al., 2013a). These results demonstrate the capacity of the native plants to remove substantial copper from copper-contaminated soils, comparing with other studies, because the plants used are seedlings and will grow for many years substantially reducing the PEN number by high biomass production and consequently remove copper.
Conclusions
The native species of the Brazilian Atlantic forestry are grateful for growth in their native habitats, with high adaptability and resistance into the environment; however, if they are used for recovering Cu(II)-contaminated areas, they may have some problems with high Cu(II) concentrations in the soils such as up to 320 mg/kg of the pseudo-total copper in this Oxisol, different from the Brazilian legislation. The Cu(II) doses reduced the morphological parameters of the seedlings for both E. crista-galli and S. multijuga plants. The increment of Cu(II) doses in the soil affected the initial growth of the seedlings of E. crista-galli.
Phytoaccumulation of Cu(II) by seedlings of E. crista-galli and S. multijuga was predominantly promoted in the roots, its translocation to the shoots being small. So, the E. crista-galli exhibited tolerance to the doses of Cu(II) and accumulated high concentration of Cu(II) in their roots than the S. multijuga, being one interesting alternative, to reduce Cu leaching and toxicity in the environment and also recovering Cu(II)-contaminated areas.
Footnotes
Author Disclosure Statement
No competing financial interests exist.