
Abstract
Abstract
The aim of this study was to find an arsenite-resistant bacterial strain, having high oxidizing capability from As(III) to As(V) in soil and stabilize the bio-available arsenic. An As(III) oxidizing bacteria strain XS3, was isolated from arsenic-rich gold mine tailings in Xinjiang, China. XS3 exhibits high resistance to As(III) and was able to oxidize up to 6,400 mg/L in sucrose minimal salt low phosphate (SLP) medium, higher than any of the previous reported Pseudomonads. To determine the molecular basis of resistance and transformation aoxB/aioA gene encoding arsenite oxidases were amplified using a degenerate primer. The aioA gene was specific for the arsenite-oxidizing bacteria, and deduced amino acid sequence revealed 62% homology with Pseudomonas arsenicoxydans. Arsenite oxidase enzyme activity from this isolate displayed a high arsenite resistance level. Based on 16S rRNA gene sequence analysis XS3 was closely related to Pseudomonas sp. Strain XS3 showed potential for soil remediation by removing arsenite up to 62% in soluble exchangeable fraction (bio-available) from soil contaminated with 500 mg/kg. Due to high resistance, efficient transforming ability and successful soil remediation, XS3 can be used as a potential candidate for soil bioremediation of As(III) contaminated sites.
Introduction
A
Different methods have been developed for the treatment of arsenic-contaminated sites based on physiochemical or electrochemical techniques such as, chemical oxidation, precipitation, adsorption, and so on, but these methods are not preferred due to the generation of harmful byproducts and are not economically feasible, therefore it is necessary to develop an alternative method. Biological oxidation or bioremediation offers the possibility to detoxify harmful contaminants using natural biological activity. It uses relatively low-cost, low-technology method, which have a high public acceptance and can frequently be carried out on site. Previously, some arsenite-oxidizing bacterial species have been reported including Alcaligenes faecalis (Osborne and Enrlich, 1976), Pseudomonas strains (Ilyaletdonov and Abdrashitova, 1981) Thiobacillus ferrooxidans and Geobacillus kaustophilus (Leblanc et al., 1995; Cuebas et al., 2011), Thermus genus (Gihring et al., 2001), and Comamonas terrae (Chitpirom et al., 2012). However, there is still a need to find the bacterial species having high resistance level and high oxidizing ability because many species have high resistance levels, but low transforming ability. Some bacteria show resistance to arsenite through the activity of an arsenite oxidase (Muller et al., 2003) or resistance to both arsenite and arsenate through the genes of the ars operon. Earlier, Cai et al. (2009) described arsenite oxidase gene (aoxB/aioA gene) that was involved in transformation by arsenite-oxidizing bacteria in Achromobacter sp. Arsenite oxidation was catalyzed by a periplasmic arsenite oxidase. This enzyme consists of two subunits and was encoded by the gene cluster aoxB/aroA/asoA (large Mo-pterin subunit), 88 KDa (825 amino acid), and aoxA/aroB/asoB (small Fe-S Rieske subunit) 14 KDa (134 amino acid) respectively (Mukhopadhyay et al., 2002).
Arsenite oxidase plays a significant role in detoxification mechanism, which can also impact the mobility and speciation of arsenic in environment. This arsenite oxidase enzyme allows the bacteria to metabolize high concentration of arsenite. Bacteria with aioA-like sequences have been found in different arsenic-contaminated soil and water system in the environment (Inskeep et al., 2007). Efficiency of in situ remediation is the vital part of the selected strain, therefore this study deals with identification of efficient indigenous As(III) oxidizing bacterial strain from an arsenic-contaminated site that has high tolerance to arsenite, the ability to transform As(III) to As(V), and survives in the soil.
Materials and Methods
Isolation, enrichment, and screening
Arsenic-rich gold mine tailing sample were collected from Karamay, Xinjiang, China. This sample had an arsenic concentration of about 200 mg/kg. Five grams mine tailing sample was added to a 250 mL conical flask containing 50 mL of sucrose minimal salt low phosphate (SLP) medium. The constituents of SLP medium are follows: sucrose (1%), (NH4)2SO4 (0.1%), K2HPO4 (0.5%); MgSO4 (0.05%), NaCl (0.01%), yeast extract (0.05%). Flasks were incubated on orbital shaker at 30°C for 5 days; following which 3 mL of this enrichment culture was inoculated into fresh SLP medium. This procedure was repeated twice and arsenic-resistant bacteria were isolated by serial dilution, by plating on SLP agar (1.5%) medium supplemented with 200 mg/L As(III). Plates were then incubated in biochemical oxygen demand chamber for 24 h at 30°C. Morphologically different colonies were isolated and purified by repeated subculturing on SLP agar plate with 200 mg/L As(III). Resistant bacterial strains were further screened for their As(III)-oxidizing ability by using silver nitrate (AgNO3) screening method described by Simeonova et al. (2004). The strain was grown in SLP medium containing 100 mg/L As(III) for 2 days at 30°C. The plates were flooded with 0.1 M AgNO3 solution. Brown precipitate around the colony indicated positive As(III) oxidation reaction. One efficient As(III)-oxidizing strain was selected for further study and designated as XS3.
Determination of As(III) resistance
Determination of arsenite resistance was done by growing XS3 in SLP liquid medium amended with different concentrations of As (III) (100, 200, 400, 800, 1,600, 3,200, and 6,400 mg/L). Three flasks for each concentration were inoculated with appropriate (2% inoculums previously grown) cell suspension growing in SLP medium. The growth was evaluated by measuring the optical density at 600 nm using DU*800 UV/vis spectrophotometer (Beckman Coulter) after 48 h of incubation at 30°C.
As(III) oxidation by XS3
To investigate the ability of XS3 to oxidize arsenite, XS3 were inoculated into 50 mL of SLP medium containing different concentrations (100, 200, 400, 800, 1,600, 3,200, and 6,400 mg/L) of arsenite and incubated at 30°C with 120 rpm shaking. Control experiments using noninoculated, sterile media with 100, 200, 400, 800, 1,600, 3,200, and 6,400 mg/L arsenite were also conducted under the same conditions. Two milliliter of sample from inoculated and noninoculated (control) experiments were taken over time to measure cell density and for determination of arsenic speciation. Optical density was measured at 600 nm using a DU*800 UV/vis spectrophotometer.
Samples were then centrifuged and decanted, acidified by adding concentrated trace metal-grade HCl to 1% (v/v), and stored at 4°C for <7 days before arsenic analysis. As(III) and total As concentration were measured by a hydride generation atomic flame spectrometer (AFS-800; Beijing Jitian Instrument Co., Ltd.) as described by Glaubig and Goldberg (1988) and Voth-Beach and Shrader (1985). The AFS parameters and optimized condition were as follows: negative high voltage of photomultiplier (280 V), carrier gas argon, with flow of 400 mL/min and atomizer height (10 mm). The calibration curve was prepared using the known concentrations of arsenic and linearity was maintained (r > 0.999) during analysis.
Induction of arsenite oxidase activity
To ensure the arsenite oxidase system was constitutive or inducible, the XS3 strain was grown in SLP medium without arsenic and subcultured thrice in the same medium. After the third subculture, cells were inoculated separately into SLP medium supplemented with 100 mg/L As(III) and without As(III). After overnight incubation at 30°C, cells were centrifuged at 10,000 rpm at 4°C for 15 min and washed twice with saline solution. The cells were suspended in 20 mL of MES (morpholinoethelenediol sulfonic acid) buffer (pH 6), and As(III) was added to a final concentration of 1.33 mM. The flasks were incubated at 30°C for 2 h. Samples were removed after every 30 min, and As(III) concentration was determined.
Spheroplast preparation and determination of arsenite oxidase activity
For spheroplast XS3 was grown in SLP medium supplemented with 100 mg/L As(III). Fully grown cells were centrifuged (10,000 rpm for 10 min) and pelleted, resuspended in 20 mM Tris-HCl, 0.1 mM phenylmethylsulfonyl fluoride, 10 mM EDTA (pH 8.4) containing 20% sucrose, and treated with 0.5 mg/mL of lysozyme for 40 min at 25°C as described by Anderson et al. (1992). Arsenite oxidase activity was determined based on the transfer of reducing equivalent from arsenite to 2,4-dichlorophenolindophenol (DCIP). Reduction of DCIP (60 μM) was monitored at 600 nm in the presence of 200 μM sodium arsenite in 50 mM MES buffer (pH 6.0) at 25°C. Specific activity was defined as micromoles of DCIP reduced per min per milligram of protein.
Identification aioA/aoxB gene
Polymerase chain reaction (PCR) amplification for aioA gene was performed using bacterial genomic DNA as a template. Sequence of the PCR primer for aioA gene were (Forward primer 5′GTCGGYTGYGGMTAYCAYGYYTA3′ Reverse primer 5′YTCDGARTTGTAGGCYGGBCG3′) as described by Inskeep et al. (2007). The PCR program included 35 cycles of denaturation (94°C, 1 min), annealing (48°C, 1 min), and extension (72°C, 2 min). Initial denaturation (94°C, 2 min) and final extension (72°C, 5 min) were performed. The amplified PCR product was purified using a Mini BEST DNA Fragment Purification kit (Takara). The presence of aioA gene in the PCR product was confirmed by sequencing at Beijing Genomics Institutes, Beijing with an ABI Prism 3730 DNA analyzer (Applied Biosystems). Sequence was analyzed by BLASTN (gene sequence) and BLASTX searching tool to deduce the amino acid sequences. Sequences were manually edited for the same length using Multalin program and phylogenetic tree was constructed using Neighbor-joining methods with MEGA 5.0 software.
Soil bioremediation
Farmland soil samples were collected from Urumqi, China. Samples were air-dried and sterilized before study. The As remediation experiment was carry out in beaker containing 200 g of soil supplemented with As(III) and up to 500 mg/kg was mixed thoroughly into the soil. The overnight grown bacterial cell culture (107 CFU/mL) in SLP medium was added to soil samples. Control remediation study was also conducted in a similar way without adding bacterial cells. The experiment was performed at room temperature 30°C and terminated in 7 days. The soil samples were analyzed for As concentration from different fractions of soil as described by Tessier et al. (1979), again arsenic was measured using hydride generation atomic flame spectrometer with parameters and conditions as described above.
Identification of bacterial isolate
16S rRNA gene sequence and phylogenetic analysis were done by extracting genomic DNA from cultures grown overnight using a simple lysis method as described in Karn and Reddy (2012). One microliter (100 ng) of purified genomic DNA served as template. 16S rRNA gene was amplified using MG25+LongGene PCR systems (Scientific Institute Co., Ltd) using degenerate primer. The amplification product was gel purified using QIA-Gel extraction kit (Qiagen), and ligated into the pGEM-T easy vector according to manufacturer's instruction (Promega, Inc.). Ligated plasmid was transformed into Escherichia coli DH5α cells by calcium chloride treatment and heat shock method. After screening of the positive clones, the sequence was generated using an Applied Biosystems (ABI Prism 3730 DNA analyzer) sequencer (DNA sequencing facility; BGI). The sequence was compared to the available DNA sequences from type strains in GeneBank (www.ncbi.nlm.nih.gov/) using BLASTN sequence match tool. The sequences were aligned using MultAlin (http://bioinfo.genotoul.fr/multalin/multalin.html) program and the alignment was manually corrected and phylogenetic tree was constructed using the MEGA 4.0 Tamura et al. (2007) software. For analysis, 1,000 bootstrap replicates were performed to access the statistical support for the tree.
Nucleotide sequence and bacterial culture accession numbers
DNA sequences described in this study have been submitted to GenBank under accession number JX448407 (16S rRNA gene) and JX657678 (aioA gene). The bacterial culture XS3 was also submitted at CGMCC China under accession number 1.12351.
Data analysis
Data were statistically analyzed by analysis of variance and the mean differences were compared by Tukey–Kramer Multiple Comparison Test at p < 0.05. Three replicates were maintained for each treatment. All the analyses were performed using GraphPad Prism (v 4.03) software.
Results
Isolation and characterization of bacterial isolate
Twelve bacterial strains were isolated from arsenic-rich gold mine tailing by enrichment technique. Out of 12 different bacterial strain, only one strain, XS3 showed high resistance and was positive for As(III) oxidization reaction by silver nitrate screening method. Other strain having lower resistance and showing negative As(III) oxidization were discarded and strain XS3 was further studied. XS3 was short-rod shaped, Gram-negative, and motile in character. 16S rRNA gene sequence data of the isolate were analyzed by BLAST search. The BLAST analysis showed 98% (97% coverage) identity to Pseudomonas thivervalensis strain McBPA3 (Genbank accession no. JQ317814). A phylogenetic tree was constructed based on 16S rRNA gene sequence by Neighbor-joining using MEGA software (version 5.0). The phylogenetic analysis revealed that XS3 was grouped with Pseudomonas sp. and hence designated as Pseudomonas sp. XS3 (Fig. 1). The XS3 strain showed high resistance toward As(III), being able to grow at high concentration.
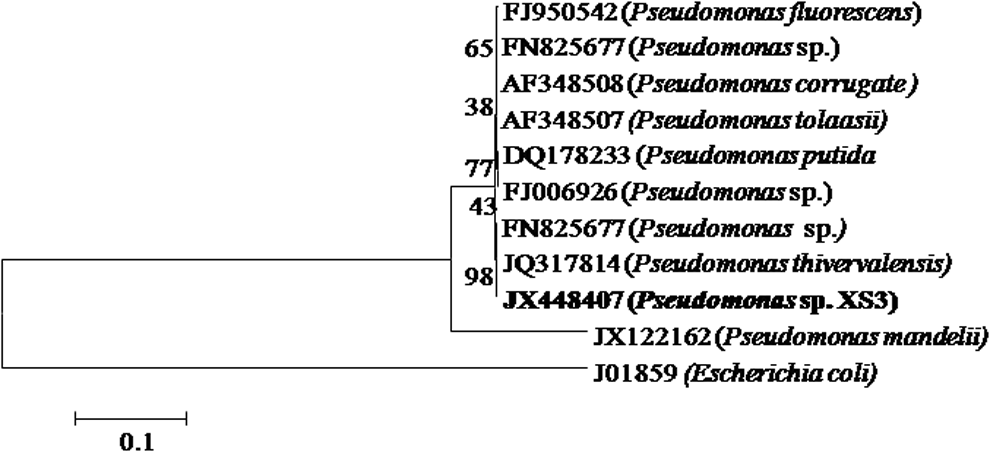
Neighbor-joining tree based on 16S rRNA gene sequence of XS3 from this study along with sequences are available in the GenBank database. Numerical values indicate bootstrap percentile from 1,000 replicates.
Growth and oxidation analysis
Growth and As(III) oxidizing ability of XS3 was observed in SLP medium containing different concentrations of As(III). The growth of XS3 strain was not significantly affected up to 800 mg/L As(III). It was observed that increased As(III) concentration has negative effect on the growth, but not affecting the oxidizing ability up to 1,600 mg/L. At 3,200 and 6,400 mg/L less growth was observed less presumably due to the higher concentration (Fig. 2a). Growth of XS3 significantly increased just after inoculation and showed maximum growth at 24 h of interval. XS3 strain efficiently oxidized As(III) in SLP medium. The oxidizing ability of XS3 was observed at different time intervals at a concentration of 200 mg/L as shown in Fig. 2b. XS3 was able to oxidize >90.15% at concentration of 800 mg/L and 87% at 1,600 mg/L. A decrease in As(III) oxidation was noticed at concentration 3,200 and 6,400 mg/L 68.44% and 56%, respectively (Fig. 3a). XS3 began As(III) oxidation at the start of exponential phase and finished by the end of exponential phase, it was also observed that cell density continued to increase even after oxidation of As(III).


Induction and determination of arsenite oxidase activity
Arsenite oxidase activity of XS3 was induced by As(III). The arsenite oxidase activity was not detected when cells were grown in medium without arsenic (Fig. 3b). However, As(III) oxidization was observed ∼40% for strain XS3 when cells were grown in presence of As(III), which suggest that genes were upregulated by the addition of As(III). The presence of arsenite oxidase was observed in both supernatant and biomass during the sphaeroplast preparation. Approximately 10% of total activity was found in released content from the periplasmic space, thus the protein appears to be located in the membrane fraction, showing 90% of the activity. Specific activity was calculated for arsenite oxidase from both supernatants and biomass. We found that the supernatant had 0.012 μM protein/min and the biomass had 0.1 μM protein/min.
Arsenite oxidase gene (aioA or aoxB gene)
Isolated bacterial strain, XS3 was found to be positive for the (Mo-pterin subunit) arsenite oxidizing gene or genetic basis of arsenite resistance. The gene sequence showed maximum identity 98% (coverage 91%) to aioA gene of uncultured organism clone VD-1d arsenite oxidase Mo-pterin subunit (aioA) gene (GeneBank accession no. DQ380576 and DQ380577) isolated from soil or environmental sample, further the gene was also showed similarity 94% (coverage 91%) to aioA gene of Pseudomonas arsenooxidans (GenBank accession no. FN824370). The deduced amino acid sequence from aioA showed 62% identity with the large subunit of the P. arsenooxidans (CBL95251) (Fig. 4). An out grooves was created using Vibrio splendidus 12B01aioA gene (ZP00992047). On the basis of deduced amino acid sequences a phylogenetic tree was constructed, which showed similarity with previously reported arsenic-resistant As(III)-oxidizing bacteria and closely related bacterial isolates.
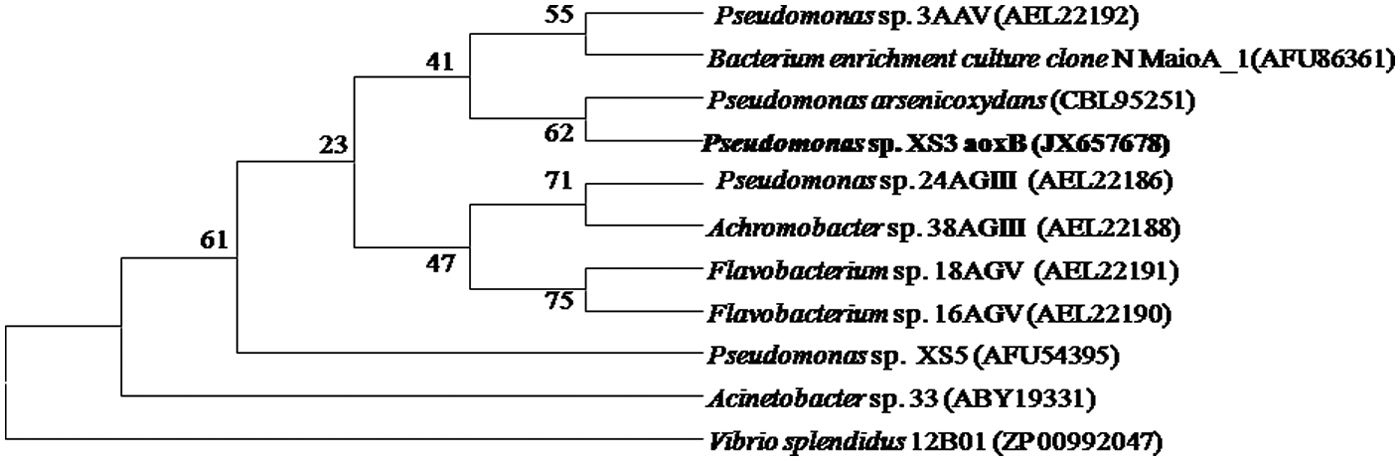
Bootstrap consensus tree based on deduced amino acid sequences among the arsenite oxidase. Scale bar 0.01 means sequence substitutions.
Soil bioremediation
Xinjiang is one of the most polluted regions in China and this region is facing multi-metal problem due to presence of gold, copper, uranium mines, and coal mines. The soil from this region is contaminated by many metal species, and therefore the present aim was to find efficient transforming microbial species. Pseudomonas sp. XS3 was highly effective in removing As from the soluble exchangeable fraction of soil. The soluble exchangeable As concentration in control soil samples was very high (138.5 mg/kg) and in bioremediated samples significantly lowered (48 mg/kg). The amount of Fe-Mn bound and residual fraction of As in bioremediated soil samples was remarkably higher than in controls (Fig. 5). No significant changes were observed in organic and carbonate bound fractions. According to World Health Organization (WHO) guidelines As concentration below 10 μg/L is considered within the safe limit, however, it will be difficult to reduce the As concentration into the safe limit.
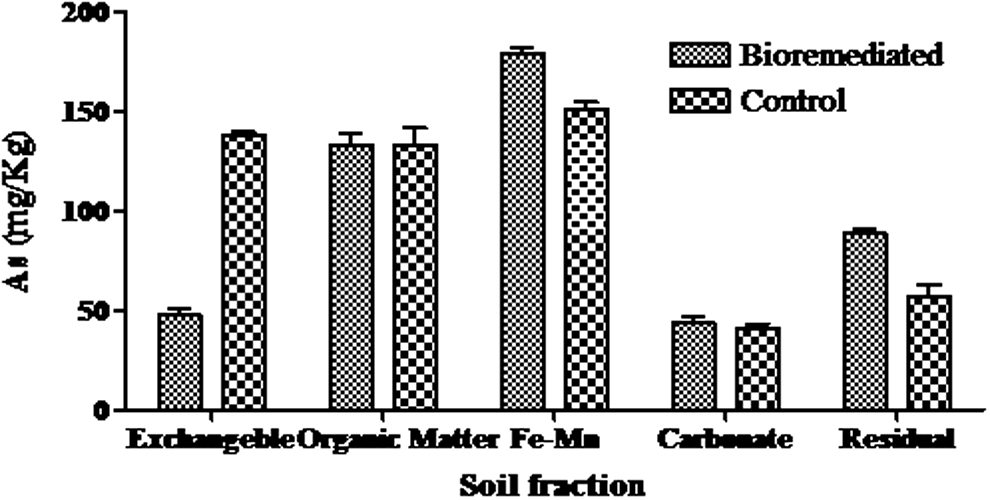
Arsenic concentration in different soil fractions for control and bioremediated contaminated soil samples.
Discussion
As(III)-oxidizing bacteria can act as a natural remover of arsenite pollution by decreasing its bioavailability by transforming As(III) to As(V) and can assist As removal from soil and waters through bio-processing because As(V) can be adsorbed much more strongly than As(III) (Ahmann et al., 1994; Afkar et al., 2003). Some As(III)-oxidizing bacteria have been isolated from various environments, especially mesophilic ecosystems (Afkar et al., 2003; Chowdhury, 2004). These bacteria belong to >25 genera, mainly from the Proteobacteria phylum (Messens et al., 2002; Arai et al., 2003). In this study, we investigated an efficient As(III)-oxidizing bacteria belonging to phylum Proteobacteria that exists in gold mine tailings with high levels of arsenic concentration. By 16S rRNA gene sequence analysis the present isolate belongs to Pseudomonas sp. Nearly full-length 16S rRNA gene sequences were used for bacterial identifications, which shows maximum identity to P. thivervalensis. Previously, phylogenetically diverse arsenite-oxidizing bacteria have been isolated from soil and water environments. XS3 was able to oxidize and transform at high concentration up to 6,400 mg/L, much higher than any of the previously reported Pseudomonads. The biological oxidation of arsenite using bacteria is of great interest for decontamination of arsenic-contaminated soil or ground water. Microbial oxidization of arsenite is a critical link in the global As cycle by transforming the toxic arsenite into less toxic arsenate, which is less mobile (Ehrlich, 2002).
The genus Pseudomonas is broadly represented among the arsenic-resistant strain isolated from environmental samples (Turner, 1949; Ilyaletdonov and Abdrashitova, 1981). De Vicente et al. (1990) isolated a Pseudomonas aeruginosa strain from marine environment, which also showed arsenic resistance. The genus Pseudomonas, is composed of ubiquitous bacteria endowed with a remarkable adaptability to diverse environments (De Vicente et al., 1990). Previous studies have reported As(III)-oxidizing ability of 100 mM (Green, 1918) and 60 mM (Turner, 1949). Studies conducted by Escalante et al. (2009) reported As-resistant isolates capable of tolerating As(III) concentration to 40 mM. Bachate et al. (2012) reported Achromobacter sp. SPB31 exhibited resistance up to 40 mM. Bahar et al. (2012) Stenotrophomonas panacihumi was found to tolerate as high as 60 mM arsenite. Srivastava et al. (2012) found Staphylococcus sp. NBRIEAG-8, which was capable of growing at 1,500 mg/L arsenite As(III). From the literature we found that many bacteria showed resistance to arsenite with limited transforming ability whereas XS3 displayed high resistance ability and successfully oxidized As(III) at high concentrations, which makes it a potential microbe for the bioremediation purposes. XS3 exhibited arsenite oxidase enzyme activity; a membrane bound protein was observed in released periplasmic and membrane fractions. This arsenite oxidase enzyme allows the bacteria to metabolize high concentrations of arsenite. Arsenite oxidase is probably transported over the cytoplasmic membrane via the TAT system and remains inner membrane attached by an N-terminal trans-membrane helix of the Rieske subunit as suggested by Lebrum et al. (2003). Arsenite oxidase has been found in other bacteria having resistance Hermimonas arsenooxidans (Muller et al., 2003), Thiomonas sp. (Duquesne et al., 2008), and Rhizobium sp. NT26 (Santini et al., 2000) under aerobic condition. Arsenite oxidase plays an imperative key role in the detoxification process.
There are other different mechanisms adopted by bacteria and fungi for the removal of heavy metals (Holan and Volesky, 1995). The cellular response in the presence of metals includes various processes such as biosorption by cell biomass, active cell transport, binding by cytosolic molecules, entrapment into cellular capsules, precipitation and oxidation–reduction reactions (Gadd, 1990; Lovely and Coates, 1997), and protein-DNA adduct formation (Zhitkovitch and Costa, 1992) and induction of stress proteins (Ballatori, 1994). In addition to chromosomal genes that function for uptake of inorganic arsenic as alternative substrates to useful nutrients, many microbes possess genes that specifically confer resistance to inorganic arsenic, both arsenate As(V) and arsenite As(III), as their natural primary substrates (Silver and Phung, 2005). Different primer sets have been successfully developed to specifically target the functional aioA gene (Inskeep et al., 2002; Lievremont et al., 2003) encoding the large molybdenum-bearing catalytic subunit of As(III)-oxidase, an enzyme of the dimethyl sulfoxide (DMSO) reductase family. The expression level of aioA gene could probably be applied to monitor environmental arsenic-contaminated levels. Campos et al. (2009) reported an aioA gene was isolated from two arsenite-oxidizing bacteria isolated from volcanic rocks. This study also found the arsenite-oxidizing aioA gene from bacterial isolate XS3, which indicates microbe has the genetic basis for arsenite resistance and oxidizing ability. This also indicates that aioA gene may be specific for aerobic bacteria and useful for resistance of arsenite-oxidizing microorganism in environment. A phylogenetic analysis of the arsenite oxidizers based on the translated sequence of arsenite-oxidase gene using BLASTX tool showed a 62% similarity with Pseudomonas arsenicoxdans (CBL95251), indicating genomic stability of the aioA gene. Previously Prasad et al. (2009) also reported arsenite oxidase activity played an important role in the detoxification of high arsenic concentrations. This result suggests that bacteria capable of oxidizing arsenite using arsenite oxidase have an elevated arsenite resistance level.
Microorganisms are known to play an important role in the biochemical cycle of arsenic, through its conversion to species with different solubility, mobility, bioavailability, and toxicity (Silver and Phung, 2005). The result obtained after in situ remediation clearly showed a significant reduction of active As(III) was taking place in the bioremediated sample compared to the control. Pseudomonas sp. XS3 successfully removed up to 63% As(III) from the soluble exchangeable fraction of soil. Fe-Mn bound and residual bound As(III) are tightly associated with soil so this fraction is not easily released. According to Manning et al. (1998) As tends to bind more strongly with ferric hydroxide in the soil and is very stable. Heavy metals form a complex with organic matter in soil due to humic acid and fulvic acid (Stevenson, 1982) and retain metals in exchangeable form. The present research studied an indigenous efficient bacterial isolate having high arsenic oxidizing capability. This bacterium can be applied for the bioremediation efficiency of As(III) contaminated soil decreasing the bioavailability of As(III) and reducing its toxicity in the contaminated ecosystem.
Footnotes
Acknowledgments
This work was supported by the Program of 100 Distinguished Young Scientists of the Chinese Academy of Sciences, National Natural Science Foundation of China (U1120302 and 21177127), and Chinese Academy of Sciences Fellowships for Young International Scientists (No. 2011Y2ZB06).
Author Disclosure Statement
No competing financial interests exist.