
Abstract
Abstract
Efficacy of bioremediation for soil contaminated with polycyclic aromatic hydrocarbons (PAHs) may be limited by the fractions of soil-bound PAHs that are less accessible to PAH-degrading microorganisms. In previous test-tube-scale work, submicellar doses of nonionic surfactants were screened for their ability to enhance the desorption and biodegradation of residual PAHs in soil after conventional bioremediation in a laboratory-scale, slurry-phase bioreactor. Polyoxyethylene sorbitol hexaoleate (POESH) was the optimum surfactant for enhancing PAH removal, especially the high–molecular weight PAHs. This work extends that concept by treating the effluent from the slurry-phase bioreactor in a second-stage batch reactor, to which POESH was added, for an additional 7 or 12 days. Surfactant amendment removed substantial amounts of the PAHs and oxy-PAHs remaining after conventional slurry-phase bioremediation, including more than 80% of residual 4-ring PAHs. Surfactant-amended treatment decreased soil cytotoxicity, but often increased the genotoxicity of the soil as measured using the DT-40 chicken lymphocyte DNA damage response assay. Potential ecotoxicity, measured using a seed germination assay, was reduced by bioreactor treatment and was reduced further after second-stage treatment with POESH. Of bacteria previously implicated as potential PAH degraders under POESH-amended conditions in a prior study, members of the Terrimonas genus were associated with differences in high–molecular weight PAH removal in the current study. Research using submicellar doses of surfactant as a second-stage treatment step is limited and these findings can inform the design of bioremediation systems at field sites treating soil contaminated with PAHs and other hydrophobic contaminants that have low bioaccessibility.
Introduction
P
Due to their hydrophobicity, PAHs are often strongly associated with nonpolar soil domains, such as soil organic matter, combustion residue, and nonaqueous-phase liquids, and therefore may be unavailable to degrading microorganisms (Cornelissen et al., 1998; Huesemann et al., 2004; Lei et al., 2004; Richardson and Aitken, 2011). A 1993 USEPA case study concluded that slurry-phase bioreactor treatment could be used to effectively remediate PAH-contaminated soil but that “effective desorption of compounds from weathered soil can be an intractable problem” (USEPA, 1993). Application of surfactants has been proposed to enhance the bioavailability of PAHs to degrading organisms with the goal of increasing PAH removal in contaminated soils and is the subject of several review articles (Makkar and Rockne, 2003; Li and Chen, 2009; Cameotra and Makkar, 2010; Elliot et al., 2011; Bustamante et al., 2012). Surfactants can increase the rate of PAH desorption from soil at aqueous-phase concentrations both above (Tiehm et al., 1997; Grasso et al., 2001) and below (Yeom et al., 1996; Zhu and Aitken, 2010; Frutos et al., 2011) their critical micelle concentrations (CMC).
Previous research using field-contaminated soil suggests that surfactant addition is most beneficial for systems in which PAH biodegradation is limited by low bioaccessibility. This could be the case, for example, with soil treated in a conventional bioremediation system after which residual PAHs resist further desorption (Zhu and Aitken, 2010; Bueno-Montes et al., 2011) or for soil that does not exhibit substantial PAH removal during conventional treatment without surfactant (Tiehm et al., 1997; Di Gennaro et al., 2008). The addition of surfactant during the first-stage treatment of soils that already exhibit substantial PAH removal in surfactant-free controls often results in either no improvement or even inhibition of PAH removal (Deschenes et al., 1996; Kim and Weber, 2005; Lei et al., 2005; Zhu and Aitken, 2010; Bueno-Montes et al., 2011). For these reasons, surfactant-amended treatment can be applied efficiently as a second-stage treatment if conventional treatment without surfactant fails to meet soil cleanup goals.
A second aspect of surfactant-amended treatment that has received limited attention is the use of low doses of surfactant, which would reduce the cost of remediation. Few studies treating field-contaminated soil have investigated the use of doses corresponding to aqueous-phase surfactant concentrations below the CMC in the soil slurry system (sub-CMC doses) (Kim and Weber, 2005; Lei et al., 2005; Zhu and Aitken, 2010).
We recently screened five nonionic surfactants at sub-CMC doses at the test-tube scale for their ability to enhance the biodegradation of the residual PAHs remaining in soil after bench-scale aerobic, slurry-phase bioremediation (Adrion et al., 2016). Polyoxyethylene sorbitol hexaoleate (POESH) surfactant was most effective at enhancing the removal of the residual high–molecular weight (HMW) PAHs compared to further incubation for the same amount of time without surfactant. Despite substantial removal of PAHs, including 5 of the 7 PAHs considered probable human carcinogens by USEPA, POESH-amended treatment was associated with an increase in soil genotoxicity.
Both PAH removal and the formation of potentially genotoxic transformation products will be affected by the bacterial community active during bioremediation. In a small-scale companion study, we demonstrated the substantial effect of surfactant addition on the bacterial community (Singleton et al., unpublished data*). That work implicated several groups of bacteria with increased representation in gene libraries correlated with increased degradation of HMW PAHs in surfactant-amended samples. Among the groups most influenced by amendment with POESH were uncharacterized members of the Alphaproteobacteria, and sequences associated with the Terrimonas, Phenylobacterium, Sediminibacterium, and Luteimonas genera (Singleton et al., unpublished data).
In the present study, we expand on our previous work by utilizing POESH at sub-CMC doses to treat soil slurry effluent from the aerobic bioreactor in a second-stage batch bioreactor. The objective of the study was to evaluate the reproducibility of the two-stage treatment concept at a larger than test-tube scale and to compare two different residence times in the second-stage bioreactor. We evaluated the effects of two-stage treatment on PAH and oxy-PAH removal; on soil cytotoxicity, genotoxicity, and ecotoxicity; and on the bacterial community.
Materials and Methods
Experimental design
In our previous work screening effects of different nonionic surfactants on PAH removal from soil after treatment in a semicontinuous, slurry-phase bioreactor (Adrion et al., 2016), a POESH dose of 24-mg POESH/g-dry soil corresponded to an aqueous-phase surfactant concentration well below the CMC (260-mg POESH/L aqueous-phase concentration) and a negligible fraction of total PAH mass present in the aqueous phase after surfactant addition. We selected the same dose of POESH to implement the two-stage treatment concept in the present study.
In the previous screening experiments, the slurry removed from the bioreactor was centrifuged, resuspended in a fresh buffer, and amended with surfactant at the test-tube scale; tubes were shaken vigorously on an orbital shaker. In the present study, the whole slurry removed as effluent from the bioreactor (i.e., not centrifuged and resuspended in buffer) served as influent to a second-stage, batch bioreactor to which POESH was added; mixing was accomplished with a metal stir bar on a magnetic mixer. To permit sorption of the surfactant to the soil before commencing aerobic biodegradation (Zhu and Aitken, 2010; Adrion et al., 2016), the surfactant-amended slurry in the second-stage reactor was mixed under a continuous stream of nitrogen for 48 h before aerobic conditions were re-established.
Three trials of second-stage batch treatment were conducted at a residence time of 7 days, and three trials were conducted at a residence time of 12 days. Residence times were chosen based on preliminary test-tube scale experiments indicating that most PAH removal (Supplementary Fig. S1) and changes in genotoxicity (Supplementary Fig. S2) occurred between day 3 and 21. A preliminary experiment was also conducted to compare the efficacy of a supra-CMC dose (60-mg/g-dry soil) identified in our previous work (Adrion et al., 2016) with that of the sub-CMC dose (Supplementary Fig. S3), as described in Supplementary Data.
For each trial, a different batch of effluent soil slurry from the first-stage bioreactor was used (6 different batches altogether). Because the first-stage bioreactor was operated in a manner that produced effluent once a week (described in the First-stage treatment section), the trials were conducted over 6 consecutive weeks; trials at 7-day residence time were alternated with the trials at 12-day residence time. Photographs of the first- and second-stage reactors are provided in Supplementary Figure S4.
Chemicals
PAH standards (EPA 610 PAH mixture and individual PAHs), 9,10-phenanthrenequinone (PQ), 9,10-anthraquinone (AQ), POESH, and dimethyl sulfoxide (DMSO) were obtained from Sigma-Aldrich (St. Louis, MO). 9-fluorenone (FLO) and benz[a]anthracene-7,12-quinone (BAQ) were purchased from Acros Organics (Morris Plains, NJ). All other solvents were high-performance liquid chromatography (HPLC) grade and were obtained from either Fisher Scientific (Pittsburgh, PA) or VWR International (Radnor, PA).
First-stage treatment
Source soil was collected from a former manufactured gas plant site in Salisbury, North Carolina, in the vicinity of the former tar well, 1.2 m below the surface. Soil was blended and sieved through a 10-mm sieve and then stored at 4°C. The soil contained 64% sand, 30% silt, and 6% clay, with total organic matter of 8.6%, as determined by a thermogravimetric method (Lukasewycz and Burkhard, 2005), and extractable organic matter of 14-mg/g dry soil. The sieved source soil was sieved again through a 3.35-mm sieve and is, from this point forward, referred to as “feed soil.” Feed soil underwent first-stage treatment in a semicontinuous, laboratory-scale, aerobic bioreactor. The bioreactor was made of stainless steel, had a working volume of approximately 2 L, a solids content of 15% (w/w), and average solids retention time of 35 days. Every 7 days, 400 mL of the treated slurry was replaced with a slurry of feed soil in a pH 7.5 buffer containing 5-mM phosphate and 2.5-mM ammonium nitrate (bioreactor buffer). Each week, 100 mL of the effluent slurry was analyzed for PAHs and oxy-PAHs as described in the Slurry extraction and analysis section. The remaining 300 mL of effluent slurry served as influent to the second-stage bioreactor.
Preliminary experiments
We conducted a preliminary experiment to test the selected dose of POESH using whole slurry from the first-stage bioreactor, rather than centrifuged slurry that had been resuspended in fresh buffer, to ensure that the sorptive properties of the soil had not changed since the previous screening experiments (Adrion et al., 2016). Slurry from the first-stage bioreactor was centrifuged at 2130 g for 30 min and the supernatant was collected. A 2.08 g (dry weight) aliquot of the centrifuged soil was added to each of triplicate 30-mL glass centrifuge tubes with PTFE-lined silicon septa screw caps. POESH (50 mg) was added to each tube. Supernatant was then added back to each of the tubes to bring the solids content to 15% (w/w). The tube headspace was evacuated and replaced with nitrogen, and then, the tube was put on an orbital shaker at 275 rpm for 48 h. After 48 h, the tubes were centrifuged at 2700 g for 30 min. The supernatants (aqueous phase) were syringe filtered through 0.8-μm polycarbonate membrane filters and analyzed for pyrene concentration by HPLC, and surfactant concentration by measuring surface tension as described in Adrion et al. (2016).
Second-stage treatment
Two second-stage batch bioreactors were set up in parallel to accommodate treatment of the weekly effluent removed from the first-stage bioreactor. Each second-stage bioreactor was a 1-L glass filter flask, fitted with a stainless-steel tube piercing a silicon stopper at the top of the flask to provide gas input to the headspace; the side arm was left open to allow gas to escape.
Every week, 300 mL of first-stage treated slurry was transferred to a second-stage reactor for batch treatment with surfactant. The slurry was stirred on a magnetic stir plate using a 3.2-cm metal stir bar at approximately 750 rpm. To add surfactant, 1.34 g of POESH was added to a 30-mL glass centrifuge tube and mixed with 10 mL of deionized water before being transferred to the second-stage reactor; this mass corresponded to a dose of 24 mg POESH per g dry soil. An additional 10 mL of deionized water was then used to rinse the remaining mass of POESH into the reactor. The surfactant-amended slurry was allowed to mix under a stream of nitrogen delivered through the gas tube at a flow rate of 60 mL/min for 48 h. After 48 h, a 10 mL aliquot of slurry was removed from the reactor while mixing using a glass pipette, transferred to a 30-mL glass centrifuge tube with PTFE-lined cap, and centrifuged at 2700 g for 30 min. The supernatant was syringe filtered through a 0.8-μm polycarbonate membrane filter to measure the aqueous-phase surfactant and pyrene concentrations as described in Adrion et al. (2016).
After the 48-h anoxic mixing period, the nitrogen flow was discontinued and air was delivered continuously through the gas tube for the duration of the aerobic treatment period (either an additional 7 or 12 days, as explained in the Experimental design section).
Slurry extraction and analysis
Slurry from the first- and second-stage bioreactors was centrifuged at 2130 g for 30 min. For each batch of slurry, a single 10 mL aliquot of the supernatant was syringe filtered through a 0.8-μm polycarbonate membrane filter to measure the aqueous-phase surfactant and PAH concentrations as described in Adrion et al. (2016). Four 3 g aliquots of the wet centrifuged soil were each mixed with 10 g of sodium sulfate and solvent extracted overnight twice, each time with 10 mL of acetone and 10 mL of dichloromethane, and analyzed for 14 PAHs by HPLC with fluorescence detection as described elsewhere (Richardson et al., 2011). Four oxy-PAHs were analyzed by liquid chromatography–mass spectrometry as described in Supplementary Data. Soil moisture content was determined in triplicate by heating 1 g wet weight aliquots of centrifuged soil in aluminum weighing dishes at 105°C for 24 h. Untreated soil used to feed the first-stage bioreactor was prepared for analysis by slurrying in bioreactor buffer, centrifuging, and extracting the centrifuged soil as described above. Concentrations of PAHs and oxy-PAHs in the feed soil and bioreactor-treated soils are provided in Supplementary Tables S1–S3.
Cytotoxicity and genotoxicity analysis
Effects of the two-stage treatment process on soil cytotoxicity and genotoxicity were evaluated using the solvent extracts from the first-stage and second-stage treated slurries and from the untreated feed slurry. For each soil slurry sample, 10 mL aliquots from each of the four replicate solvent extracts were combined in a preweighed vial and evaporated to dryness under a gentle stream of nitrogen. Residue mass was then determined gravimetrically. Cytotoxicity of the residues reconstituted in DMSO was evaluated in triplicate using a 96-well plate-based DT40 chicken lymphocyte DNA damage response assay adapted from Ridpath et al. (2011) and Hu et al. (2012). The untreated feed soil was evaluated in quadruplicate. Cytotoxicity was measured using the DT40 parental cell line, which is not deficient in any DNA damage repair or response pathway. The Rad54−/− and Rev1−/− DNA repair-deficient mutants were tested alongside the isogenic DT40 parental cell line to measure genotoxicity specifically and were selected because of their reported sensitivity to soil residue in previous experiments (Hu et al., 2012; Adrion et al., 2016). The Rad54−/− knockout is deficient in the homologous recombination DNA repair pathway, while the Rev1−/− knockout is deficient in the translesion synthesis pathway (Yamazoe et al., 2004). LC50 values (mg residue/mL media) were calculated by fitting the log residue concentration vs. % survival in GraphPad Prism version 6.05 for Windows. The LC50 values, as measured in residue mass, were converted to equivalent soil LC50 values (mg soil/mL media). The relative LC50 for each mutant cell line (a unitless ratio equal to LC50 of the mutant divided by the LC50 of the parental cell line) was calculated as a measure of genotoxicity as described by Hu et al. (2012). Values of relative LC50 less than 1.0 are indicative of genotoxicity, demonstrating that the residue is more cytotoxic to the DNA repair-deficient mutant than to the parental cell line.
Ecotoxicity analysis
We evaluated the effects of soil treatment on potential ecotoxicity by using the phytotoxicity test with the lettuce (Lepidium sativum) seed following standard procedures (USEPA, 1996) and reported elsewhere for application to PAH-contaminated soil (Eom et al., 2007; Manzo et al., 2008). Untreated feed soil was prepared for ecotoxicity analysis by slurrying in bioreactor buffer, centrifuging the slurried soil, and freezing the soil until analysis. To collect an adequate mass of first-stage treated soil, first-stage treated slurries produced the week before commencing second-stage treatment and those produced in the 2 weeks after ending second-stage treatment were centrifuged, combined, and frozen until analysis of the combined sample. Similarly, wet centrifuged soil (totaling approximately 80 g) was collected from each of the three 7- or 12-day second-stage treatment trials, combined, and kept frozen until ecotoxicity analysis of the combined samples. Before ecotoxicity analysis, the combined samples of first- and second-stage treated soil were analyzed for PAH concentrations by HPLC as described above (Supplementary Table S4). Ten L. sativum seeds were placed in each of five replicate Petri dishes containing 15–18 g of fresh soil (equivalent to 10 g dry soil and subsequently saturated with water) depending on the percent moisture of each sample. A clean soil (with no detectable PAHs, pesticides, or PCBs and only trace levels of transition metals) exhibiting no toxicity was used as a negative control, while K2Cr2O7 (100-μg/g dry soil) was used as a positive control. Seeds were allowed to incubate in darkness for 72 h at 25°C and the number of germinated seeds and total root elongation were measured. A germination index (GI) was calculated as the product of mean seed germination and mean root elongation, and expressed as a percentage relative to the negative control soil (USEPA, 1996). The positive control resulted in no seed germination and %GI value of zero.
Bacterial community analysis
We compared the bacterial communities of the second-stage treatment reactors among themselves and with those of prior POESH-amended small-scale incubations described in Singleton et al. (unpublished data). A brief description of incubation and analysis methods used in the small-scale incubations is presented in Supplementary Data. For each treatment trial, duplicate samples of slurry were collected from the first-stage reactor immediately before transferring effluent to the second-stage reactor and from the second-stage reactor at the end of each trial. DNA was immediately extracted from each sample and the bacterial community later examined by denaturing-gradient gel electrophoresis (DGGE). To recover soil from reactors, 1450 μL aliquots of slurry (approximately 0.25-g soil dry weight) were centrifuged for 3 min at maximum speed in a benchtop microcentrifuge and the supernatant discarded. The pellet was used for DNA extraction using the FastDNA Spin Kit for Soil (MP Biomedicals, Santa Ana, CA) following the manufacturer's directions, except that cell lysis was achieved by vortexing horizontally secured tubes at maximum speed for 6 min and DNA was eluted from the binding matrix with TE buffer. DNA from POESH-amended soil samples was additionally purified using the QIAEX II Gel Extraction Kit (Qiagen, Germantown, MD) before molecular analyses. PCR for the examination of bacterial communities by DGGE was performed using general 16S rRNA gene primers 341F-GC and 517R (Muyzer et al., 1993). DGGE gels consisted of 10% acrylamide with either a 30–60% or 35–55% denaturant gradient and a nondenaturing stacking gel (the narrower denaturant range was used to help separate bands during excision; see below). DNA from previous small-scale POESH incubations was recovered as described in Singleton et al., unpublished data and briefly in Supplementary Data.
Bacteria corresponding to one specific DGGE band were identified. This band was of interest because it was the only prominent band notably absent from the sample of a second-stage treatment trial with reduced PAH removal (see Results section). A sterile razor blade was used to cut out bands from adjacent samples where the band was present. The DNA was then diffused from the gel matrix by overnight incubation in 50 μL sterile water at 4°C. Two μL of eluted DNA was reamplified using primers 341F (without a GC clamp) and 517R and cloned using a TOPO TA Cloning Kit with vector pCR™4-TOPO® and a Top10 Escherichia coli host strain (Invitrogen/Life Technologies, Grand Island, NY). Random transformants were selected on LB plates containing kanamycin and inserts were PCR amplified from picked colonies with vector-targeted primers M13R and M13F. After checking the PCR products for the expected size and quality, five amplicons were sequenced using primer M13R at Eton Biosciences (Research Triangle Park, NC). Vector sequences were removed from the acquired sequences and compared using Sequencher 5.0 (Gene Codes, Ann Arbor, MI). Four of the 189 bp-long sequences were identical, while the fifth possessed a single mismatch to the other four. The consensus sequence was used in subsequent analyses.
The excised band sequence from this study and sequences grouped into operational taxonomic units (OTUs) during community analyses of the prior small-scale experiments (Singleton et al., unpublished data) covered different variable regions of the bacterial 16S rRNA gene. To associate the excised band sequence with a specific OTU from that prior work, the band sequence was first phylogenetically classified using the classifier program at the Ribosomal Database Project (Cole et al., 2009). The top 25 GenBank nr database BLAST hits to the excised band with sufficient length to cover both gene regions analyzed were used to create a separate local Blast+ database (Camacho et al., 2009), which was then queried using the representative sequences of abundant OTUs matching the phylogenetic affiliation of the excised band. The representative sequence with the highest nucleotide similarities to the GenBank sequences determined which OTU was most likely to represent the excised band. OTUs were previously defined using a sequence identity threshold of 97% (Singleton et al., unpublished data).
Data analysis
Averages and standard deviations of percent removals (summary statistics) for each trial were calculated through propagation of error, using the concentrations of PAHs and oxy-PAHs in the second-stage treated soil and in the corresponding batch of first-stage treated soil or untreated feed soil. All statistical analyses were done in SAS Enterprise Guide 6.1 (SAS Institute, Cary, NC). Comparisons of concentrations of PAHs and oxy-PAHs in soil before and after second-stage treatment were conducted using two-sample t-tests (two-tail homoscedastic, α = 0.05). To compare removals among all the trials for each PAH, a one-way ANOVA followed by Tukey's HSD test (α = 0.05) was conducted using summary statistics of percent removal. One-sample t-tests were conducted on each value of relative LC50 for the feed soil and first-stage treated soil to determine whether the values were significantly different than 1.0 (two-tail homoscedastic, α = 0.05). Comparisons were made between the LC50s and relative LC50s of second-stage treated soil with those of the corresponding first-stage treated soil for each trial using two-sample t-tests (two-tail homoscedastic, α = 0.05). Comparisons between seed germination indices were made using two-sample t-tests (two-tail homoscedastic, α = 0.05).
Results
Preliminary experiments
A preliminary test-tube-scale experiment was conducted to confirm that adding POESH to whole slurry removed from the first-stage bioreactor would be similar to our observations in prior work (Adrion et al., 2016), in which the slurry from the first-stage bioreactor was centrifuged and the soil resuspended in fresh buffer. In the test-tube-scale experiment, the aqueous-phase POESH concentration after 48 h of anoxic mixing was 3.4 ± 0.6 mg/L, well below the CMC of 260 mg/L, and corresponding to less than 1% of total surfactant mass added to the system (>99% was sorbed to soil). The pyrene concentration was less than the lower limit of quantification (LLOQ) of 29 μg/L. However, both aqueous-phase POESH and pyrene concentrations were substantially higher after 48 h of anoxic mixing in the second-stage bioreactors than in the preliminary test-tube-scale incubations (Table 1), although in all trials the aqueous-phase POESH concentrations were below the nominal CMC. After 48 h of anoxic mixing in the second-stage bioreactors, pyrene was consistently present at aqueous-phase concentrations above its reported pure compound solubility in water of 132 μg/L (Mackay, 1992), indicating solubilization of pyrene. By the end of all trials, however, all aqueous-phase surfactant concentrations were less than the LLOQ of 3.3 mg/L. Likewise, at the end of all trials, individual aqueous-phase PAH and oxy-PAH concentrations were less than their respective LLOQs. This corresponded to individual PAH and oxy-PAH masses present in the aqueous-phase representing less than 3% of their residual masses in the soil after first-stage treatment except for acenaphthene, which was less than 12%.
An asterisk indicates liquid-phase pyrene concentration in excess of its pure compound aqueous solubility of 132 μg/L (Mackay 1992). All POESH concentrations were below the CMC of 260 mg/L.
CMCs, critical micelle concentrations; POESH, polyoxyethylene sorbitol hexaoleate.
PAH and Oxy-PAH removal
First-stage treatment of PAH-contaminated soil removed substantial amounts of PAHs and oxy-PAHs, as shown in Supplementary Tables S5 and S6. Second-stage batch treatment with the nonionic surfactant POESH for either 7 or 12 days resulted in substantial removal of the residual PAHs and oxy-PAHs remaining in the effluent from the first-stage bioreactor (Fig. 1). Removal was most substantial for the 4-ring PAHs (FLA, PYR, BaA, and CHR). Removal of naphthalene was low (<20%) and not consistently significant. Second-stage treatment removed significant amounts of the 5-ring PAHs (BbF, BkF, and BaP), but removal was more variable across trials than for the other PAHs. The second trials of both the 7- and 12-day incubations, for example, had significantly less removal of BbF and BkF (Fig. 2). For the 5-ring dibenz[a,h]anthracene (DBA), removal was low (<20%) and not consistently significant. The 6-ring benzo[g,h,i]perylene, which had the highest concentration of any measured PAH in the first-stage bioreactor effluent, was not significantly removed in any second-stage trial.
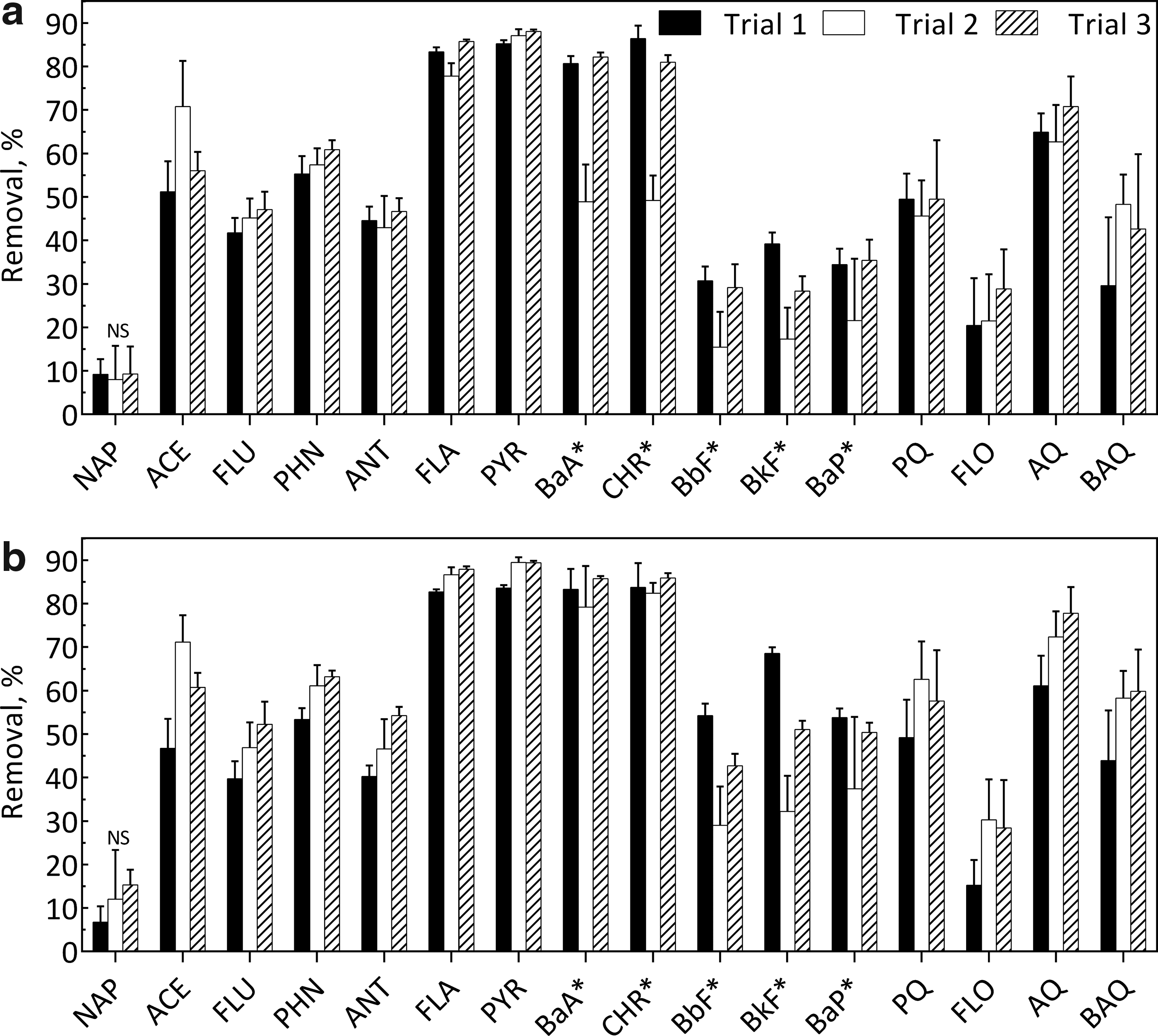
Effect of second-stage treatment on residual PAHs and oxy-PAHs in effluent from first-stage bioreactor at a residence time of
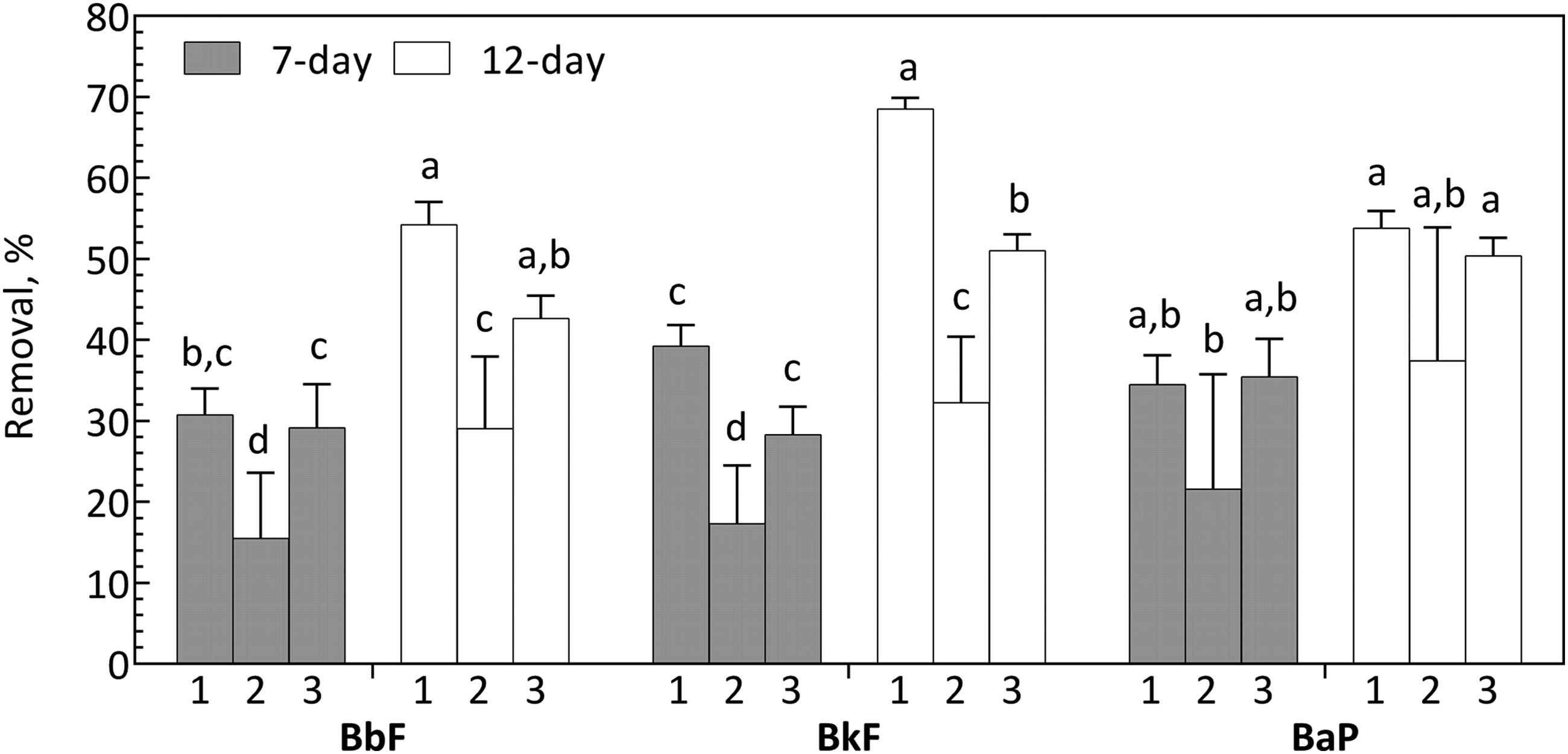
Effect of second-stage residence time on removal of selected 5-ring PAHs (data are the same as shown in Fig. 1 but rearranged). For each PAH, conditions for which there was not a significant difference (α = 0.05) in removal detected by Tukey's method are assigned the same letter. Numbers below the bars correspond to the three trials at a given residence time. PAH, polycyclic aromatic hydrocarbon.
Comparing removals between the two residence times, differences were most substantial for the 5-ring PAHs, BbF, BkF, and BaP (Fig. 2), with generally greater removal at the 12-day residence time. Cumulative removals relative to the untreated feed soil for the combined first- and second-stage treatment are presented in Supplementary Tables S5 and S6 to illustrate the overall impact of the two-stage treatment concept.
Cytotoxicity and genotoxicity
Cytotoxicity and genotoxicity were determined with the DT-40 chicken lymphocyte assay; cytotoxicity was evaluated with the parental cell line, and genotoxicity by comparison with the DNA repair-deficient mutants, Rad54−/− and Rev1−/−. All second-stage trials reduced the cytotoxicity of the soil relative to the first-stage effluent (Fig. 3a). The average relative LC50s of the first-stage treated soil across the 6 trials were 0.71 ± 0.08 for the Rad54−/− mutant and 0.79 ± 0.09 for the Rev1−/− mutant. Both relative LC50 values were significantly less than 1, indicating that the effluent from the first-stage bioreactor was genotoxic. Relative to first-stage treated soil, all second-stage trials at the 7-day residence time increased genotoxicity as measured with the Rev1−/− mutant (Fig. 3c). Only trial 2 of the 7-day treatment trials significantly increased genotoxicity as measured with the Rad54−/− mutant (Fig. 3b). Trials 1 and 2 of the 12-day second-stage treatment significantly increased genotoxicity as measured with both mutants, while no significant effect was observed in trial 3. There was insufficient evidence of genotoxicity for the untreated feed soil as measured with either of the DT-40 mutant cell lines, because the relative LC50s were not statistically significantly different than 1 (Fig. 3b, c). In general, effluent soil from both first- and second-stage treatment was more genotoxic than the untreated feed soil, a result that was not substantially improved with increased second-stage residence time from 7 to 12 days.

Effect of first-stage (black bar) and second-stage (white bar) treatment on toxicity to the parental DT40 cell line
Ecotoxicity
The potential for ecotoxic effects of the soil was evaluated using the phytotoxicity test with the lettuce (L. sativum) seed, which has been shown to be among the most sensitive of all soil ecotoxicity tests for PAHs (Eom et al., 2007; Manzo et al., 2008). The feed soil was highly toxic, causing no seeds to germinate and thus a percentage germination index (%GI) value of 0.0 (Table 2). Treatment with the first-stage bioreactor allowed some seed germination and growth with a %GI of 4.1 ± 2.7, which was significantly different from the feed soil. Subsequent second-stage treatment caused a further decrease in toxicity with %GI values of 13.2 ± 5.2 and 14.2 ± 4.1 for 7- and 12-day residence times, respectively. There was no significant difference between the 7- and 12-day residence times, but both were significantly different than the feed soil and the first-stage treated soil.
Single asterisk indicates significant difference from feed soil (α = 0.05), double asterisks indicates significant difference from both feed soil and first-stage treated soil (α = 0.05).
Bacterial community
The bacterial community in the first-stage bioreactor slurry used as influent for the POESH-amended second-stage bioreactor was examined by DGGE and appeared stable, with little variance observed across the six samples serving as influent to the second-stage reactor (Supplementary Fig. S5). The community profile was also highly similar to that of the first-stage bioreactor observed during our previous study of surfactant amendment at a smaller scale (Singleton et al., unpublished data) (Supplementary Fig. S5). Any variability in the bacterial community during second-stage treatment was therefore presumed not to be heavily influenced by the inoculum. A brief description of methods used to set up and analyze incubations for the smaller scale study is provided in Supplementary Data.
The scale and residence time of the second-stage treatment significantly impacted the apex community, with several clear differences between previous small-scale surfactant-amended incubations (Fig. 4, lanes 1–3) and the second-stage bioreactor samples of the present study (lanes 4–15). Differences between the POESH-amended bacterial communities at differing residence times (7 and 12 days) in the secondary bioreactor were also apparent, although some dominant bands were shared among all conditions and replicates. Of particular interest was the lack of one prominent band in the second trial of the 12-day residence time (Fig. 4, indicated arrow), which was present in both the other 12-day trials and therefore may have been related to the reduced removal of 5-ring PAHs observed in that sample (Fig. 2). The equivalent band from the third 12-day trial profile was excised and identified as representing organisms within the Terrimonas genus of the family Chitinophagaceae, similar to OTU-14 from the prior study (Singleton et al., unpublished data) (Supplementary Fig. S6). In that study, similar sequences were highly represented in 16S rRNA gene amplicon libraries of small-scale POESH-amended incubations (9% average relative abundance), but not in slurry from the first-stage bioreactor or in incubations with other surfactants (Singleton et al., unpublished data).
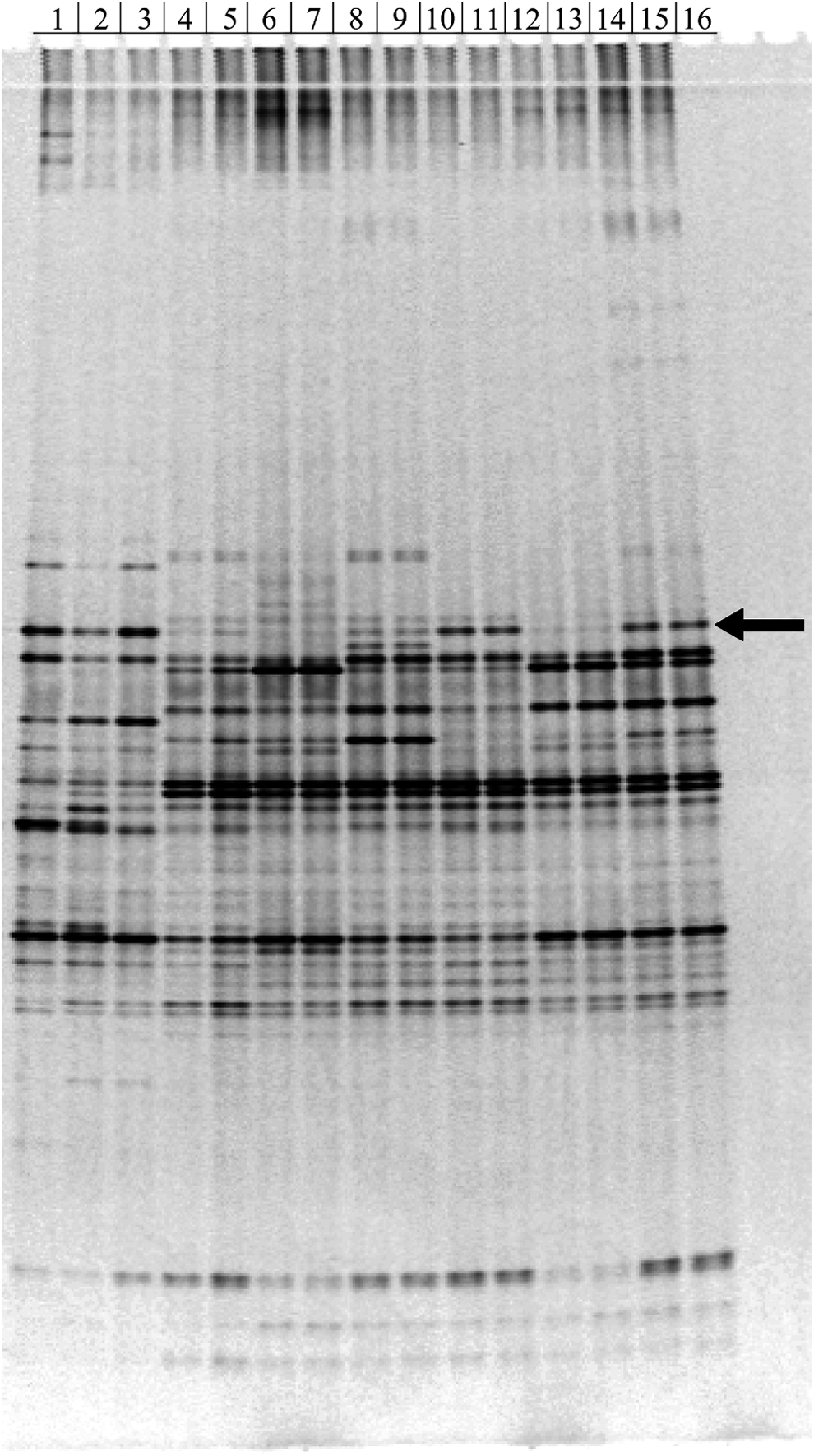
DGGE gel showing bacterial community profiles of POESH-amended soils. Lanes 1–3, triplicate small-scale POESH-amended soil incubated for 14 days from prior experiments (Singleton et al., unpublished data); lanes 4–5, 7-day second-stage treatment (first trial); lanes 6–7, 7-day second-stage treatment (second trial); lanes 8–9, 7-day second-stage treatment (third trial); lanes 10–11, 12-day second-stage treatment (first trial); lanes 12–13 12-day second-stage treatment (second trial); lanes 14–15, 12-day second-stage treatment (third trial); and lane 16, no-template PCR control. The arrow indicates the band that was excised and identified. Colors in the image were inverted and contrast adjusted to best visualize the bands using the GNU Image Manipulation Program (v2.8.0). DGGE, denaturing-gradient gel electrophoresis; POESH, polyoxyethylene sorbitol hexaoleate.
Discussion
Although the addition of surfactant has been suggested as a means of enhancing the biodegradation of hydrophobic contaminants such as PAHs in soil, few studies have emphasized that the benefits are most likely to be manifested in systems limited by low bioaccessibility. In separate studies using PAH-contaminated soils from two different sites, we reported that nonionic surfactants substantially improved the desorption and biodegradation of residual PAHs from contaminated soil that had already undergone aerobic treatment in a laboratory-scale bioreactor (Zhu and Aitken, 2010; Adrion et al., 2016). We have consistently found the fraction of PAHs remaining after first-stage treatment to be of low bioaccessibility (Zhu and Aitken, 2010; Hu et al., 2014; Adrion et al., 2016) and in previous studies found that further treatment without surfactant led to limited PAH removal (Zhu and Aitken, 2010; Adrion et al., 2016). In this study, we extended the concept by evaluating the reproducibility of a semicontinuous two-stage treatment process at the bench scale.
Additional PAH removal observed during POESH-amended treatment of effluent slurry from the first-stage bioreactor was consistent with our previous study, particularly the substantial removal of 4-ring PAHs (Adrion et al., 2016). In addition to the PAHs, removal of the four oxy-PAHs we evaluated was also enhanced by the addition of POESH. Oxy-PAHs are of concern because some are known to exhibit toxic or genotoxic effects (Chesis et al., 1984; Bolton et al., 2000; Zielinska-Park et al., 2004; Luo et al., 2011) and may also inhibit the biodegradation of parent PAHs (Kazunga and Aitken, 2000; Kazunga et al., 2001). Of these four compounds, AQ was present in the initial (untreated) soil at the highest concentration, 24 μg/g (Supplementary Table S1); overall removal of AQ in the two-stage process was 95% (Supplementary Tables S5 and S6).
Despite the fact that the oxy-PAHs were previously found to be more desorbable than the unsubstituted PAHs in the effluent soil from the first-stage bioreactor (Hu et al., 2014), the addition of POESH appeared to improve their bioaccessibility as well. Although none of the oxy-PAHs increased in concentration during biological treatment in this experiment, net increases of some oxy-PAHs have been observed during bioremediation (Lundstedt et al., 2003) and researchers have inferred the formation of oxy-PAH based on ratios of oxy-PAH to parent compound (Wilcke et al., 2014).
The most substantial removal of 3- and 4-ring PAHs occurred within the first week of second-stage treatment, as evidenced by the limited differences between 7- and 12-day batch treatments. Extending the residence time of the second-stage bioreactor to 12 days from 7 days led to greater average removal of the 5-ring compounds BbF, BkF, and BaP, although variability in the data (particularly in trial 2) made statistical comparisons less conclusive. As discussed in our previous work, we assume that POESH sorbs to and facilitates PAH desorption from the nonpolar domains found in manufactured gas plant soil (e.g., coal tar and black carbon) (Adrion et al., 2016). Hypothesized effects of surfactants on these soil domains include increased PAH diffusivity within the coal tar matrix (Yeom et al., 1996), increased interfacial surface area caused by wetting (Dong et al., 2003), and dispersion of nonpolar matrices (Kile and Chiou, 1989; Zhang and Miller, 1992; Churchill et al., 1995). It is also possible that surfactants can influence the release of soil organic matter with which PAHs associate (Markiewicz et al., 2013).
There were unidentified factors affecting aqueous-phase surfactant and PAH concentrations during second-stage treatment. Preliminary test-tube-scale incubations with whole slurry from the first-stage bioreactor agreed well with our previous test-tube-scale work (Adrion et al., 2016), suggesting that the greater aqueous-phase surfactant concentration observed in the larger scale, second-stage reactor could be due to the method of mixing. Mixing in the test-tube-scale incubations (orbital shaking) was more turbulent than that achieved during second-stage treatment using a magnetic stir bar. The second trials of both the 7- and 12-day residence times had higher aqueous-phase surfactant and PAH concentrations at 48 h, and this may have impacted the bacterial community, affecting PAH removal (trial 2 had lower PAH removal than trials 1 and 3 in each case). Reduced PAH removal at higher aqueous-phase surfactant concentrations is consistent with the significantly lower removal of 5-ring PAHs (BbF, BkF, BaP, and DBA) observed in the supra-CMC incubation compared to the sub-CMC incubation in preliminary experiments (Supplementary Fig. S3). Prior test-tube-scale experiments comparing two sub-CMC doses of a different surfactant, Brij 30, also led to significant changes in the bacterial community, and it is probable that this would be the case for POESH as well (Singleton et al., unpublished data). Since adequate interaction of the surfactant with the solid phase is important before commencing aerobic treatment, the adequacy of mixing is an important issue to consider for scale up in the two-stage treatment concept.
It is also possible that the liquid phase of the effluent from the first-stage bioreactor influenced the liquid-phase behavior of POESH and/or PAHs in the second-stage bioreactor during the anoxic mixing period. In our earlier work at test-tube scale (Zhu and Aitken, 2010; Adrion et al., 2016), the liquid phase from the first-stage bioreactor was replaced with a fresh buffer before the surfactant was added. The liquid phase of the first-stage bioreactor slurry could contain dissolved or colloidal organic matter that can act like a surfactant (reducing surface tension) and can facilitate the apparent solubilization of hydrophobic compounds (Akkanen et al., 2005; Grolimund and Borkovec, 2005; Markiewicz et al., 2013). However, if such factors did influence the apparent liquid-phase concentrations of POESH and/or PAHs in the second-stage bioreactor, we would have expected to observe this effect in the preliminary experiment at the test-tube scale (i.e., it should have been independent of mixing intensity).
Increased genotoxicity resulting from treatment of the feed soil in the first-stage bioreactor observed in this study is consistent with our previous studies on the same contaminated source soil (Hu et al., 2012, 2014; Chibwe et al., 2015). As summarized in our previous work (Hu et al., 2012; Adrion et al., 2016), the Rad54−/− and Rev1−/− mutants are sensitive to damage caused by the major mechanisms of PAH-induced genotoxicity, and Rad54−/− in particular is sensitive to a broad range of genotoxic compounds. Although second-stage treatment removed substantial amounts of 4- and 5-ring PAHs, including some considered to be human carcinogens, in the majority of trials, second-stage treatment increased genotoxicity relative to the effluent soil removed from the first-stage bioreactor. While the bacterial community in the first-stage treated slurry appeared stable over time, variability in the effect of bioreactor treatment on soil genotoxicity between treatment trials could be due to other unmeasured fluctuations in bioreactor conditions occurring over this several-week-long experiment.
Overall, the genotoxicity of remediated soil will depend both on the remaining parent compounds (including PAHs and other contaminants) and the formation or removal of any products of incomplete microbial metabolism. Reduction of parent-PAH concentrations does not always correspond to a reduction in genotoxicity (Hughes et al., 1998; Gillespie et al., 2007; Lemieux et al., 2009; Hu et al., 2012). Because second-stage treatment made the soil less cytotoxic, but more genotoxic, it is possible that surfactant treatment causes transformation of cytotoxic compounds that are not genotoxic themselves into genotoxic products. For example, Zielinska-Park et al. (2004) found that bacterial transformation products such as pyrene- and fluoranthene-quinones can cause oxidative DNA damage in vitro using calf thymus DNA and HeLa S3 cells. The authors speculated that during biodegradation, bacteria may metabolically activate PAHs such as pyrene, which might not otherwise be activated to an appreciable extent by mammalian metabolic systems. Such bacterial activation can, therefore, transform relatively nongenotoxic PAHs such as pyrene, or other nongenotoxic contaminants in the soil, into redox-active products.
We found a small but significant reduction in the potential ecotoxicity of soil following the first-stage bioreactor treatment, as indicated by the seed germination assay. A larger reduction in toxicity was observed following 7 days of batch treatment in the second-stage reactor, but no additional significant improvement was seen after 12 days. Even after the second-stage reactor, the GI was only 13–14% of the control soil, indicating that substantial toxicity remained. However, the PAH concentrations we measured were one to two orders of magnitude higher than those measured by others who reported similar %GI values for the same plant species (Eom et al., 2007; Manzo et al., 2008). The reason for the lower apparent soil toxicity at higher PAH concentrations we observed compared to others is not known, but is likely related to differences in the bioavailability of the PAHs.
Bacterial community profile analyses by DGGE indicated substantial changes during second-stage treatment with POESH. The scale of the incubation also appeared to impact the structure of the community, with substantial differences between the DGGE banding patterns of DNA isolated from our previous small-scale POESH-amended incubations and those of DNA isolated from second-stage treated soils in the present study. Prior work, in which we evaluated differences among the bacterial communities between different surfactants and between doses of the surfactant Brij 30, illustrated how small changes in surfactant concentration, even at sub-CMC doses, resulted in the increased relative abundance of gene sequences from some bacterial genera and the apparent inhibition of others (Singleton et al., unpublished data). Of particular interest in the present study was one DGGE band represented in two of the 12-day trials but absent in the remaining 12-day trial that coincidentally displayed reduced HMW PAH removal (BbF-BaP) and had a substantially higher aqueous-phase surfactant concentration. The specific organism(s) represented by that band were determined to be probable members of the Terrimonas genus. Sequences associated with the Terrimonas genus were among a few prominent groups of bacteria that significantly increased in relative abundance in previous small-scale incubations with POESH, which had enhanced HMW PAH removal compared to incubations without surfactant amendment (Singleton et al., unpublished data).
Several years before the current study, Terrimonas sequences were very abundant (about 7% of the total community sequences) during initial startup of the first-stage bioreactor, but their relative abundance decreased over the first several months of bioreactor operation (Singleton et al., 2011). Members of the Terrimonas genus have been linked to PAH degradation in soil from a municipal solid waste composting site that was spiked with anthracene (Zhang et al., 2011) and more significantly were recently linked to the metabolism of BaP in forest soil (Song et al., 2015). As the Terrimonas DGGE band was the only prominent band missing from one 12-day trial profile in comparison to the other 12-day trial replicates, there is strong circumstantial evidence for its involvement in HMW PAH removal; however, further investigation is required to confirm its relevance.
This work demonstrated the effectiveness of a semicontinuous two-stage process for the enhanced removal of PAHs from contaminated soil. In the first stage, soil was treated in a bioreactor without surfactant to remove the most readily bioaccessible fractions of PAHs and oxy-PAHs. Effluent from the first-stage bioreactor was further treated in a second stage, comprising bench-scale batch, surfactant-amended bioreactors that removed substantial amounts of the residual PAHs and oxy-PAHs. The observation that parent PAH removal did not necessarily correspond to a reduction in genotoxicity, however, highlights the need for further research to identify genotoxic products to improve risk management and remediation strategies.
Increasing the residence time of the second-stage bioreactor from 7 to 12 days had limited effect on removal of the targeted contaminants except for several five-ring PAHs, highlighting the need for research to determine whether further removal is limited by desorption or by undetermined microbial factors. Although we propose that enhanced PAH removal is primarily due to improved bioavailability, it is also possible that surfactants could enhance PAH removal by acting as cosubstrates for PAH-degrading bacteria. In the previous smaller scale study, however, we did not find an association of HMW PAH removal with probable surfactant degraders isolated from the soil (Singleton et al., unpublished data). PAH removal and changes in soil cytotoxicity and genotoxicity during bioreactor treatment are likely affected by the bacterial community, which was shown to change as a result of second-stage treatment with surfactant.
Overall, the two-stage treatment concept we evaluated may be a promising method of maximizing the removal of PAHs during bioremediation. Factors to consider for scale-up include the adequacy of initial anoxic mixing and the effect of residence time in both stages. Although we found from screening experiments (Adrion et al., 2016) that the optimum surfactant was POESH, in other cases the optimum surfactant and its dose may be site specific and therefore should be evaluated in preliminary bench-scale treatability studies. In addition, since surfactant might leach from the treated soil, potentially altering contaminant bioavailability, field applications should consider the possible impacts of returning surfactant-laden soil to the remediated site or of disposal in a landfill.
Footnotes
Acknowledgments
This work was supported by the National Institute of Environmental Health Sciences under the Superfund Research Program (grant 5 P42 ES005948) and grant P30 ES010126. We thank Margaret Bertoni of the North Carolina School of Science and Mathematics for conducting the supra-CMC preliminary experiment.
Author Disclosure Statement
All authors confirm that no competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.