
Abstract
Abstract
Biofouling is a pervasive challenge in industrial and medical settings with enormous economic and health impacts. Use of quorum-quenching phages is a potential solution in either combating the biofilm or inhibiting biofilm formation when an engineered phage is designed for a specific bacterial system. In the event of a real-life application of synthetic phage, it is necessary to consider the effect of environmental conditions on the synthetic phage. This study focused on both (1) stability of phage lysate under different types of carbon sources and (2) competition of the engineered phage in comparison to a wild-type (wt) phage. Optimal results were found (in plaque forming units [PFUs]) with glucose as a carbon source and were kept stable for almost 30 days. Results suggested that competition happens before 4 h. After that time, polymerase chain reaction results from PFU samples showed an increase in T7aiiA, which indicates that these conditions favored the phage replication of the engineered phage over the wt at 27 h in a monoculture system. When both stability and competition were combined, the mix of phages showed a trend similar to T7aiiA only primarily after 8 days.
Introduction
F
To reduce fouling in membranes, mainly in severe pore plugging situations, chemical processes (Kuberkar and Davis, 2001), physical processes such as backflushing (Guizard and Rios, 1996; Le-Clech et al., 2006), biological processes that involve biocides, and even other processes such as the use of nanoparticles (Rosenhahn et al., 2008) can be applied. A potential new approach in biological processes is the use of bacteriophages. Bacteriophages or phages are bacterial viruses that can infect bacteria, the host cell, and can cause lysis to the host cell. Among all phages that are abundant in nature, the T-phages are one of the most abundant types (Fu et al., 2010). T-phages are shown to be able to infect Escherichia coli (Lu and Collins, 2007) in a single and mixed species bacterial biofilms containing Pseudomonas aeruginosa (Pei and Lamas-Samanamud, 2014).
The T7 phage is a complex phage that presents a rapid lysis after 25 min of infecting the host cell. However, the T7 phage may show a delay in enzymatic activity, which can be due to the nonspecificity of phages to the biofilm matrix (Fu et al., 2010; Liao et al., 2012). One disadvantage of using phages is the physical and metabolic characteristics of biofilms, more specifically the extracellular polymeric substance (EPS) matrix of the biofilms, which can limit the antibiofilm effect (Tait et al., 2002). Though many phages naturally carry genes encoding polysaccharide-degrading enzymes that can degrade EPS to gain access to their bacterial hosts buried inside the biofilm (Tait et al., 2002; Glonti et al., 2010; Cornelissen et al., 2012), there is no single enzyme or simple enzyme mixture that can degrade all biofilm polysaccharides (Wingender and Flemming, 2010).
Thus, to be effective, T7 phages require a proper enzymatic system to guarantee biopolymer degradation of the matrix (Sillankorva et al., 2010; Siringan et al., 2011). A study with an engineered T7 phage with polysaccharide depolymerase DspB (Lu and Collins, 2007) has had good results in quorum quenching biofilms of single species while an engineered T7 phage with AiiA has had good results for both singles and mixed species (Pei and Lamas-Samanamud, 2014).
In the application of a synthetic phage in the environment, two aspects should be considered: (1) impact on the phage, that is, its stability in the long run within a given environment and (2) impact on the environment, that is, whether the phage has unintended negative consequences. Results from Gladstone et al. (2012) showed that quorum-quenching phages that have high specificity to a bacterial strain tend to be overgrown by wild-type (wt) phages on a synthetic media, in which, both types of phages would be in constant competition. Thus, the main objectives of this research were (1) to observe stability through phage behavior over a period of time with the addition of host cells and a carbon source, and (2) promote competition for the same host cell by combining both the natural phage and the engineered phage.
Materials and Methods
Bacterial strains
E. coli strain BL21 and TG1 (lacI:kan) were kindly provided by Dr. Timothy K. Lu of Massachusetts Institute of Technology. E. coli BL21(DE3) and P. aeruginosa PAO1 were provided, respectively, by Drs. Tao Wei and Robert Renthal of University of Texas at San Antonio. E. coli MG1655 was provided by Dr. Thomas Woods of Pennsylvania State University. All bacterial strains were cultured overnight at 37°C in Luria–Bertani (LB) broth (Miller; Thermo Fisher Scientific, Waltham, MA) liquid or agar media with no antibiotics added.
Construction of engineered bacteriophages
The engineered T7 bacteriophage was generated by inserting the aiiA gene in the T7select415-1 phage display vector (EMD Millipore, San Diego, CA), which contains a modified T7 genome with deletion of a small portion of the nonessential early genes and incorporation of the multiple cloning site at the 3′ end of gene 10B.
Determination of phage count
Determination of the phage counts in the lysates expressed as plaque forming unit (PFU) were adapted from (Corbin et al., 2001; Sambrook and Russell, 2001). Briefly, 100 μL of host cell (overnight) were infected with the respective phage to the concentration of 103 PFU. Five milliliters of soft agar (0.6% agarose) were mixed with 50 μL of the host cell lysate and poured on LB agar. Plates were kept at 37°C for 3–4 h and plaques were formed in the bacterial lawn. Those plaques were counted and the concentration was given by the number of PFU/mL. Three replicates were conducted for each sample.
Surface water experiments
Surface water samples from Leon Creek in San Antonio, TX were filtered (0.20 μm pore size) and the filtrate was used as media for the growth of E. coli BL21 and the phage. Three samples in triplicate were conducted in parallel: one containing basal media (no carbon source), the second containing 20% R2A in basal media (intermediate media with 2% glucose), and the third one containing 20% glucose in basal media. Basal media was prepared according to protocol adapted from Sambrook and Russell (2001). A volume of 250 mL of surface water was mixed with 250 mL of corresponding media. Fifty milliliters of E. coli BL21 overnight and 10 μL of phage 1011 PFU/mL were added to the samples. Flasks were kept at 22°C and under agitation. Both PFU and chemical oxygen demand (COD) were monitored.
A carbon source was added to the samples every time the COD values reached 50% reduction from the initial concentration of each sampling. Samples were kept running until phage replication (given by PFU) was above or similar to the concentration of PFU added at the beginning of the experiment. Temperature was kept constant at 22°C. Conductivity, pH, and COD were monitored daily throughout the experiment. An unpaired one-sided t-test was performed to determine whether there was a significant difference between the mean values (p < 0.05).
Competition between T7wt and T7aiiA
To verify what type of phage would have greater advantage in a different type of environment, phages from T7aiiA and T7wt were combined, thus mixing both phages in the media. It was expected that both phages present in the media would compete for the same host cell (the concentration of host cell was not changed when phages were added to the system individually), promoting competition. The experiments were run in triplicate.
Using a single culture, E. coli BL21 were grown overnight at 37°C. Ten milliliters of this culture were transferred to 90 mL of LB media and grown for 1 h at 37°C. Ten microliters of T7wt and T7aiiA phage mix (1011 PFU/mL total) were added to the sample and left to grow for 3 h. After 3 h, 10 mL of this solution was transferred to a new flask of 100 mL of media (10 mL E. coli BL21 + 90 mL of LB broth). The total time of the experiment was 27 h, but samples of 1 mL were taken every 3 h to be analyzed in terms of PFU to identify the stability of the gene aiiA in the media after a period of 27 h.
To verify which phage was predominant at the end of each experiment, an end point polymerase chain reaction (PCR) was carried out using primers that would only target the gene aiiA. This approach was used to verify the presence of the engineered phage in relation to samples collected but it does not reflect the real number of phages present in one plaque-forming unit. The design of primers was done to match Bacillus anthracis Ames strain (AiiA protein) including a HindIII gene on the left side and a NotI gene on the right side. The setup of primers used for this PCR reactions were as follows: aiiaf (forward primer) 5′TAG AAGCTT ATGACAGTAA AGAAGCTTTA3′ and aiiar (reverse primer) 5′CTA GCGGCCGC CTATATATATTCAGGGAACAC3′. The approach was to use a simple gene detection method to verify the largest amount possible of PFU units.
To perform the PCR reaction, all PFU units (20–30 plaques/plate) were collected with pipette tips and soaked in enough MasterMix (25 μL total volume) for a PCR run. The MasterMix solution (MasterTaq Kit 1,000 U from 5 Prime) consisted of: 12.15 μL of deionized water, 2.5 μL 10 × buffer, 5 μL 5 × enhancer, 1 μL Mg2+ (25 mM), 0.5 μL dNTP (10 mM), 1.25 μL of 5 μM primer aiiaf, 1.25 μL of 5 μM primer aiiar, 0.35 μL Taq polymerase (5 U/μL), and 1 μL of sample plaque soaked in DI water.
PCR conditions were as follows: the first cycle (hotstart) was carried out at 94°C for 5 min; the second cycle was repeated 35 × (denaturation step at 94°C for 1 min, annealing step at 55°C for 1 min, and elongation step at 72°C for 2 min); and the last cycle (final elongation) was performed at 72°C for 6 min. Samples were then analyzed under UV on electrophoresis gel and the number of positive bands were counted corresponding to T7aiiA.
Competition in surface water experiments
Similar to what was described in the competition between T7wt and T7aiiA section, competition between T7wt and T7aiiA was promoted to verify which type of phage would have greater advantage in the surface water. The experiments were run in triplicate. Filtrate from Leon Creek was used in the experiments as described in the Surface Water Experiments section. Three samples were run in parallel containing no carbon source, R2A, and glucose. A volume of 250 mL of surface water was mixed with 250 mL of corresponding media. Fifty milliliters of E. coli BL21 overnight and 10 μL of phage 1011 PFU/mL were added to the samples.
For the competition flask, a mix of phages of equal concentration that added up to 1011 PFU/mL was added to the samples. Flasks were kept at 22°C on the shaker. The same parameters conductivity, pH, and COD were monitored daily. PFUs were used to identify phage infectivity. These plaques were soaked into MasterMix and PCR was performed on each of these plaques following conditions described in the competition between T7wt and T7aiiA section. An unpaired one-sided t-test was performed to determine whether the difference between the mean values was significant (p < 0.05) and an analysis of variance (ANOVA) to determine whether the ration between T7wt and T7aiiA was significant.
Water parameters
Conductivity was read using a Mettler Toledo Seven Easy. pH was read with a pH meter (Accumet Basic AB15plus). COD vials were acquired from HACH for both low and high ranges. COD vials were digested for 2 h and cooled down before they were read in a spectrophotometer at the HACH method 8000 specified wavelengths (420 nm for 3–150 mg/L and 620 nm for 200–1,500 mg/L).
Statistical test
Statistical significant differences were determined by using an unpaired one-sided Student's t-test using Microsoft Excel. The p-value smaller than 0.05 indicates statistical significance. p-Values lower than 0.01 indicates highly significant differences. ANOVA was run on ration between T7wt and T7aiiA using JMP12.
Results and Discussion
Construction of engineered phage
AHL lactonase gene aiiA from B. anthracis Ames strain (Ulrich, 2004) was inserted and packaged into the T7 DNA to generate the T7aiiA phage. Such a design enables the expression of the free AiiA enzyme that could be used to produce the quorum-quenching enzyme with antibiofilm potential.
The stability of the synthetic phage includes at least two aspects: infectivity and stability. First, infectivity of the synthetic phage, that is, the ability of the phage to lyse the host, measurable by formation of plaques in the lawn of the host E. coli BL21. Since T7 phages are able to infect the F plasmid-containing E. coli, enzymes that show specificity can be inserted into the T7 genome and applied as antifouling reagents.
In a previous study, the T7aiiA phage caused significant reduction in the biofilm (initial OD600 nm was 0.656 and 1.76, for 4 and 8 h postplating, respectively) with 74.9% (final OD600 nm was 0.165) and 65.9% (final OD600 nm was 0.6) for 4 and 8 h postplating. A reduction in T7wt of 23.8% (final OD600 nm was 0.5) and 31.7% (final OD600 nm was 1.2), in a mixed-species biofilm consisting of P. aeruginosa PAO1, E. coli TG1 and E. coli BL21 (Pei and Lamas-Samanamud, 2014) was found.
Second, the stability of the foreign gene over time in a given environment is a common concern for all genetically engineered microorganisms (GEM). The survival and persistence of GEM are known to be affected by various interrelated factors including biotic factors such as nutrients, predators, GEM density, competing microorganisms, host cell density, and abiotic factors such as temperature, pH, oxygen demand, water, salt content, and soil characteristics (van Elsas, 1992; Tappeser et al., 2002).
Therefore, when using a GEM like T7aiiA it is important to consider four main areas. The first one is whether T7aiiA works in controlling biofilms. Based on the results shown above, T7aiiA exhibited antibiofilm characteristics and the engineered phage had better performance than the T7wt. The second is to determine how long the T7aiiA remain infecting host cells without losing its specificity (infectivity). The third is to monitor and control abiotic factors that may compromise the infectivity of T7aiiA. And the fourth step is to verify whether there was competition for the same host cell between T7aiiA and T7wt (stability). The second, third, and fourth areas are the focus of this study.
Surface water experiments
Stability of host cell lysate was determined by how well phages could persist and multiply in a surface water environment with host cells added to the media. The hypothesis was that phages would remain infective as long as a certain concentration of the host cells was achieved and kept constant in the media to promote constant phage infectivity. The phage lysis would then be determined by the number of PFUs seen during the duration of the experiment.
Parameters such as conductivity, pH, and COD were monitored to determine whether variations in such parameters could affect or influence infectivity. Temperature was kept constant at 22°C to determine the influence of carbon source solely on infectivity. Both COD values in mg/L and phage counts were used to determine when the carbon source or host cells had to be added to the system. Table 1 shows parameters for basal media, R2A, and glucose, respectively.
COD, chemical oxygen demand; wt, wild type.
Results from Table 1 showed that the pH value was neutral on all types of media and even between both phages, the maximum pH was 7.3 observed in R2A media. Since surface water samples were at a neutral pH, little differences in the values of pH were observed (Table 1) and demonstrated that phages might not impact the acidity or alkalinity of the system but a buffer system is probably in place since phages naturally occur in environmental waters.
Conductivity was not as constant for both types of phages in different carbon sources. It was about 1,000 μS/cm for both basal and R2A and about 750 μS/cm for glucose. These conductivity values represent the mean value from all the data monitored for conductivity. Initial and final values are described in Table 1. Under microbial corrosion and in closed systems in which biofouling is often found, variations in these parameters have to be taken into consideration because they can be associated with physiological conditions of microbes (Beech, 2004). Future studies can be applied to look at the effect of changes on pH and conductivity but were not within the scope of this work instead focusing on the effect of carbon sources (Videla, 2002; Videla and Herrera, 2009).
As described previously, a carbon source was added to the samples every time COD values reached a reduction of 50% from the initial COD value. Even with the addition of the carbon source, COD values kept decreasing with time. COD values were different for each type of media. Basal media showed COD values in a low range (3–150 mg/L COD), while R2A showed an intermediate range and glucose a high range (20–1,500 mg/L COD). As shown in Table 1, COD values decreased to 80%, 50%, and 40% for basal, glucose, and R2A media, respectively. PFU values showed an increase in infectivity when the carbon source was added to the system and phages appeared stable for almost 30days.
It was expected that an increase in infectivity when the host cells were added would be observed and it was also expected that an increase in the host cell numbers would occur when the C source was added. However, the main finding in this study was to identify how stable the phages were throughout the experiment.
Previously, it was found by Pei and Lamas-Samanamud (2014) that phages were stable for up to 24 h when no carbon source was added to the system. Also, in that study, samples were analyzed both in terms of PFU and CFU. CFU values decreased after 12 h and were considerably low at 24 h. Since CFU measured the amount of host cells in the system, it was difficult for phages to persist if there were no host cells available. Thus, adding a carbon source periodically promotes an increase in bacterial growth and more available host cells to be lysed. Considering that most likely without a C source, there was less available energy to be metabolized by the host cell and the latter to be infected by the phage.
To verify which media showed higher phage counts and consequently, more infectivity, graphs were plotted for each system (Fig. 1). Arrows indicate when host cells and the carbon source were added after the beginning of the experiment. For basal media, only host cell diluted with basal media was added on days 4, 8, and 17. For R2A, C source was added on days 4, 8, and 15. Finally, for glucose-enriched media, C source was added on days 6, 10, and 17.
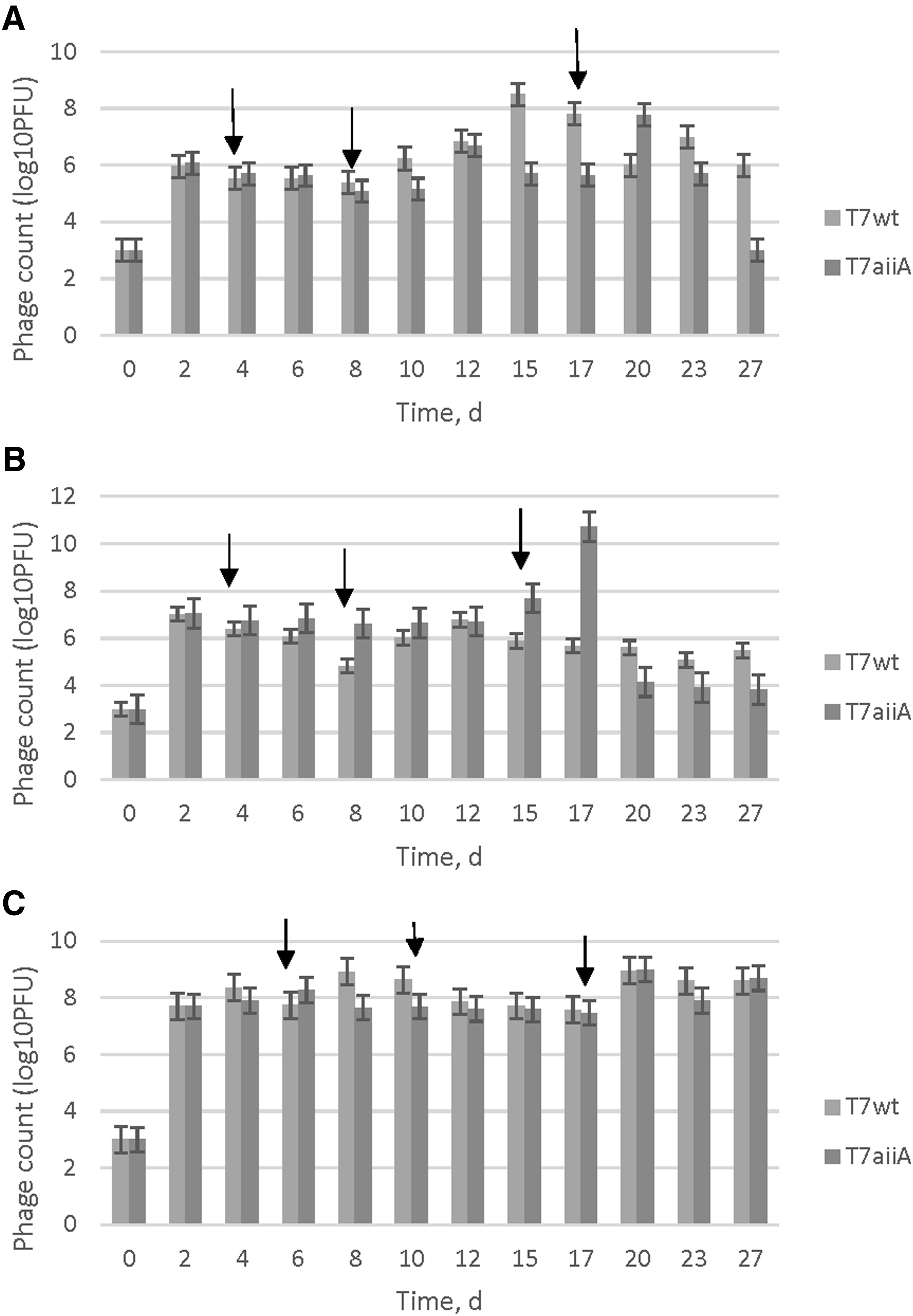
Effect of phage counts (log10PFU) on surface water samples using T7wt and T7aiiA with addition of basal media for 27 days:
As shown in Fig. 1, the carbon source and host cells were added during the first 2 weeks of the experiments. Parameters such as conductivity, pH, and COD were also monitored during that time. Experiments verified how samples would respond to no addition of a carbon source and how stability would be affected due to the presence or absence of a carbon source. Thus, samples were kept on shaker and phage counts were monitored for more than 2 weeks with no extra addition of a carbon source, up to 30 days. There was a decrease of at least onefold to colony forming units (data not shown) that indicated the host cells still persisted in the system. This could indicate that phages could still remain for a period over 30 days.
In Fig. 1A, the addition of basal media to the surface water showed an increase in infectivity from 4 log10PFU to 6 log10PFU on the first day and back to 3 log10PFU at the end of the experiment. There was an increase in infectivity of T7aiiA during the first 10 days and an increase in T7wt at the end of the experiment. This suggests that both natural phage and synthetic phage can infect host cells as long as these host cells are at a threshold concentration capable of multiplication of corresponding phage virions. Additionally, T7aiiA showed an increase in infectivity when compared to the wt phage as long as parameters like COD or phage counts were being kept at steady state. Otherwise, T7wt had a better chance of infectivity than T7aiiA. Thus, controlling COD and host cells favored T7aiiA than T7wt.
Addition of R2A to the media is shown in Fig. 1B revealing predominance of one phage over the other at different times. T7aiiA was higher than T7wt for longer periods than when compared to Fig. 1A. T7aiiA phage counts remained at about 6 log10PFU for almost 15 days of the experiment. T7wt phage counts increased whenever the carbon source was added to the system and they varied between 4 and 6 log10PFU.
Figure 1C shows the addition of glucose as a carbon source. The addition of phage in glucose-enriched media showed an infectivity of 8 log10PFU for almost all the experiments with some even reaching a maximum of 9 log10PFU. Phage in glucose-enriched media showed an increase in phage titer and, consequently, an increase in infectivity for both natural and synthetic phages that were kept constant throughout the experiment. When compared to R2A and to no carbon source, phage lysates present in the glucose-enriched media had both higher infectivity and stability. Mean values of glucose were significantly different (p < 0.05) to mean values of R2A and highly significant (p < 0.01) to mean values of basal media.
Preliminary experiments (data not shown) demonstrated that samples from surface water without filtration would show a decrease in infectivity during the first week. In fact, PFU counts did not pass the initial concentration of 3 log10PFU. This initial concentration was established by Pei and Lamas-Samanamud (2014), to be the minimum infectivity that could be found in a system to promote biofilm inhibition. In this sense, samples from surface water without filtration did not show phage titer high enough to show that phages were infecting host cells. This suggests that without filtration, surface water samples may contain bacterial cells that inhibit multiplication of phages or that other microorganisms can produce toxins that interfere with the phage lysate (Letarov and Kulikov, 2009).
When samples were filtered but no carbon source was added periodically to the system, infectivity remained at the initial concentration for 10 days before dropping considerably. Thus, the addition of a carbon source could influence the stability and infectivity of phages in surface water samples. Optimal conditions, however, vary with respect to the concentration or type of carbon source, frequency of its addition, and the importance of monitoring parameters related to both phages and carbon source.
Addition of a carbon source is common in wastewater treatment plants to remove nitrogen content and works as a way to trigger bacterial cell multiplication and substrate consumption (Grady Jr et al., 2011; Ribera-Guardia et al., 2014; Alvarino et al., 2016). In this case the carbon source favored host cell multiplication and, consequently, infectivity of phages. In these experiments, the dosage of host cells was the same over all the processes. The phages did not show significant peaks of infectivity, but were shown to be stable for a longer period of time.
Glucose is a common carbon source for bacterial cell growth but other carbon sources can be added. One example is the addition of maltose. Changes in the concentration of the carbon source can also play a role in the infectivity of phages. A maximum concentration that host cells can reach to be infected by phages is expected (OD600 = 0.5). Too many or too few cells can determine the efficiency of the process.
Optimal results, that is, phage titer increase, were found when dosage concentration of the host cell, phage concentration, and COD were taken into account. Variations on water quality parameters are important to real life applications since these values can vary constantly depending on the media.
Competition between T7wt AND T7aiiA
Upon inoculation into the microbial community, one issue that determines the fate of the synthetic phage is its relation with the wt phages in the community. Therefore, stability was studied in a short-term competition experiment between the synthetic and wt phage. A recent study indicated that synthetic T7 phages expressing the foreign polysaccharide-degrading enzyme endosialidase as a free enzyme were evolutionarily less favorable than the wt phage (Gladstone et al., 2012). One explanation is that the free enzyme was released to the biofilm, benefiting not only the enzyme-producing phages but also phages, which do not incur the metabolic cost to produce the enzyme (Sutherland et al., 2004). To determine whether the synthetic phage behaves similarly, the synthetic phages were mixed with the wt phages of the same dosage and an overnight culture of E. coli BL21.
It was important to monitor whether phages remain at the same ratio throughout the duration of the experiments. To verify this, a longer experiment was conducted in liquid media. Samples were kept for 27 h with the same initial concentration of mix phages of 1011 PFU. To maintain phage multiplication, fresh host cells were added every 3 h as adapted from Kay et al. (2011) and Gladstone et al. (2012). In their study, the genetically engineered phage is at a disadvantage when in competition with natural phages since the media favors the multiplication of natural phages (Fig. 2).

Effect of phage counts (log10PFU) on host cells for 27 h:
As seen in Fig. 2A, phage multiplication was present throughout the experiment. At 3 h, it can be observed that the number of phages present in the media were the highest. This is also true at about 6, 9, 18, and 27 h. The highest peak at 3 h could be an outlier since most experiments showed phage counts considerably lower than values found at 3 h. However, the mix of phages may indicate that at this point, both phages are multiplying at the same rate and the peak observed was the addition of both phage counts. This did not repeat again during the experiment suggesting that after 4 h if there was competition between both phages, it happened before that period. Phages can infect host cells, until there are no more host cells available. After 3 h, all host cells were infected and only the phages that would persist and consequently, present faster infectivity, would infect new host cells that were added after the 3 h period. In Fig. 2, this is true for T7aiiA. Since they are more available in the media, they infect the new batch of host cells more quickly.
It is important to point out that addition of host cells was carried out every 3 h to all samples. However, previous experiments (data not shown) established that 4 h was the amount of time required for phage at this concentration to infect all host cells in the media. After that period, the addition of host cells would boost the phage infectivity until 8 h. From 12 h, it only decreased. Data from Fig. 1 showed that this infectivity can be kept stable if both PFU and C source are considered and the decrease in phage titer happens after 15 days when no C source is added.
The behavior that is seen in Fig. 2A suggests that in a mix of phages, competition plays a major role in phage multiplication. For a single type of phage as shown in Fig. 1, there is a direct relationship between the addition of host cells and phage multiplication. However, in Fig. 2A, phage multiplication does not increase simply because host cells were added to the system. Under competition, host cells can be fully depleted or phage multiplication is not viable for a large concentration of different phages. Moreover, one type of phage can become more dominant when the number of phages present after competition decreases considerably.
As it can be seen in Fig. 2A, after 21 h, an increasing trend in phage titer can be seen. Thus, in a natural environment in which more types of phage can be present, it was important to note that there is an optimal time for phage multiplication, which was also affected by competition. Some studies (Chibani-Chennoufi et al., 2004; Griffiths, 2005) have shown that multiple infection of the same host cell can occur, in which multiple phages competed to lyse the same host cell. When two of them actually infected the same host cell, it is called mixed infection. Once the predominant phage was in the media, the phage infectivity increased and it was directly affected by the addition of host cells.
Figure 2B shows results of identification of gene aiiA through PCR. The value 1.0 indicated in the left axis is the total amount or representation of 100% of samples analyzed from PFU results. Within this amount, the ratio of each phage is indicated by markers. Both phages start at 50% and then change after 12 h. At 15 h, phage counts dropped considerably, lower even than the initial concentration, though they increased in the following hours. It is possible that the concentration of host cells, at this point, was too low to assure phage multiplication was occurring. Since new host cells were constantly added after 3 h, PFU values that follow can be associated with growth of new host cells.
Both ratio of phages were found at about the same proportion at 18 h (30% difference) and 6 h (20% difference). Differences between both phages increases at 12 h (60%) and at the end of experiment (100%). It is possible to see a trend in which the ratio of T7aiiA outnumbers the T7wt after 6 h, which suggests that the engineered phage may be better adapted to persist in this media than the natural phage.
Since there was a decline in phage counts at 18 h, it is possible that under limited circumstances, in which little or low host cells are found in the media, T7aiiA has an advantage over T7wt to infect host cells. Thus, the competition in a mix of phages did not appear to occur for T7aiiA in this scenario but it appeared for natural phages in contact with this engineered phage.
In terms of competition, the use of a broad-range activity type of enzyme showed that the competition between natural phage with the engineered phage, actually favored the multiplication of T7aiiA. An ANOVA was run on the data for the ratio between T7wt and T7aiiA. This assumption was highly significant (p < 0.001). After 27 h of experimentation, PCR results showed that T7aiiA prevailed over T7wt constituting almost 100% of all PFU samples analyzed at the end of the experiment. This result could be explained by the fact that the engineered phage showed a faster infectivity rate than the natural phage, and therefore, the engineered phage was more efficient in the application of quorum quenching to inhibit biofilm formation.
Use of PCR in this study rather than real-time polymerase chain reaction (q-PCR) is given by the fact that at least three-generation-subcultures were achieved in the 27 h experiment. In this sense, every 4 h a new generation of phage would be present in the media (time to infect all host cells). Thus, for the 27 h experiment, the final data value would be comprised of fifth or sixth generation phages and believed to be enough to determine a possible ratio of T7wt and T7aiiA.
However, this study did not take into consideration that the generation of phages is also associated with the plaque-forming unit itself. In other words, samples analyzed in the electrophoresis gel could be a mix of the engineered phage and the wt phage along with a mix of both engineered phages. Since the gene targeted was the engineered phage, only T7aiiA would be identified on the gel even though it could possibly be a mix of more phages.
A simple technique has been used to identify and verify the stability and competition of engineered phages in this work. To better investigate the environmental behavior of engineered bacteriophages in the future, q-PCR could be used to identify a specific gene in a multiple or mixed infection. Its advantage over an end point PCR is that it can target both genes and determine the exact concentration of each type of phage used at that particular sample. Nonetheless, statistical results showed that this ration for this preliminary study was significant.
Competition in surface water experiments
Parameters such as conductivity, pH, and COD were also monitored to determine whether variations in such parameters could affect or influence infectivity when both types of phages were competing with each other in the same media. Temperature was kept constant at 22°C to determine the influence of carbon source solely on infectivity, which was added following the procedure described in the Construction of Engineered Phage section. Table 2 shows parameters for basal media, R2A and glucose, respectively.
Results in Table 2 showed that values for conductivity, pH and COD are close to values presented in Table 1 for T7wt and T7aiiA separately. Though phages did not appear to impact the acidity and alkalinity of the system, the competition between both phages seem to interfere with conductivity and COD depending on what type of phage was more predominant in the system at a particular time. Values showed in Table 2 are mean values and thus, some differences may be applied through time. Since the scope of this work is on how phages behave when a specific amount of carbon source is added (Videla, 2002; Videla and Herrera, 2009) and how stable the infectivity of phages can be through time, PFUs were monitored under competition of phages for at least 2 weeks (Fig. 3).

Effect of phage counts (log10PFU) on surface water samples:
Figure 3a, c, d show a similar trend to Fig. 1. T7aiiA was favored with time in terms of infectivity except for the media in which no carbon source was added to the system. What is interesting to note is that a mix of phages have a different type of behavior from single phages until 8 days of the experiment. After this time, it showed a behavior very similar to T7aiiA. Results shown in Fig. 2, revealed that T7aiiA was favored in the system after 15 h when only the host cell was present.
In a more complex system as shown in Fig. 3b, d, f in which surface water is used, the same behavior is found to be true for T7aiiA after 10 days. Addition of a carbon source on days 4 and 8 did not seem to interfere with the outcome of the predominant phage in the media. Thus, phages could be applied in industry settings and competition among natural phages does not appear to have a negative impact on the success of the engineered phage.
The scale-up process and the implementation of the engineered phages in wastewater treatment plants or in other applications in which biofilm is a pervasive challenge must undergo a more rigorous study and assessment of risks related to the possible effects on public health and water bodies. As described in Avise (2004); Letourneau et al. (2002); Rocha et al. (2015); Romeis et al. (2011), the identification of these possible risks is gaining a lot of attention with new research techniques and better approaches to guarantee the biosafety of the environment and public health.
Conclusions
The novelty of this study was to show how persistent the engineered phage can be alongside wild phages in surface water. Results have demonstrated that the engineered phage can remain infective even under competition. The enzyme released by these type of phages can be designed to target a specific biofilm system and this management setting can have positive implications in several industrial settings in which biofouling is a pervasive challenge.
This research focused on the stability of the engineered phage in different carbon sources and in the presence of natural phages. Best results were found when dosage concentration of the host cell, phage and COD were taken into account. Addition of glucose showed better results in both stability and infectivity. These samples were kept stable for almost 30 days considering that host cell was added periodically. COD values were an indicator of when host cells or the carbon source should be added. The engineered phage appeared to be stable and showed better infectivity when in competition with wt phages.
Results suggest that competition happens before 4 h and after this time, T7aiiA was the predominant phage in the counts determined by gene identification. After 27 h, all phages were identified as T7aiiA. In surface water experiments, T7aiiA was also the predominant phage, mainly after 8 days of the experiment and was not directly influenced by the addition of a carbon source. This work demonstrated the potential for using synthetic phages in the reduction of biofilms in water treatment and other applications.
Footnotes
Acknowledgments
This study was supported by funds from the University of Texas at San Antonio. We are grateful for the technical assistance of Dr. Ruoting Pei on this project.
Author Disclosure Statement
No competing financial interest exists.