
Abstract
Abstract
A novel bacterium capable of simultaneous heterotrophic nitrification and aerobic denitrification was isolated from deep-sea sediment of the North Atlantic and identified as Serratia marcescens CL1502. The aim of this study was to study the ability, influence factors, and application in application to tannery wastewater nitrogen removal by S. marcescens CL1502. Results showed that S. marcescens CL1502 had good ability to remove NH4+-N, NO3−-N, and NO2−-N. High average NH4+-N, NO3−-N, and NO2−-N removal rates of 4.57 mg/(L·h), 4.60 mg/(L·h), and 4.42 mg/(L·h) were observed. The optimum carbon source for heterotrophic NH4+-N removal by S. marcescens CL1502 was glucose. Higher C/N ratios were favorable to heterotrophic NH4+-N removal. S. marcescens CL1502 had a good tolerance to salinity (1–5%) and Cr(III). In addition, application of strain CL1502 in actual tannery industry wastewater resulted in chemical oxygen demand (COD), NH4+-N, and TN removal efficiencies of 63.1%, 76.7%, and 72.9%. This is the first report of S. marcescens carrying out the process of simultaneous nitrification and denitrification. These results demonstrate the potential of the bacterium for use in tannery wastewater treatment for the efficient removal of ammonia.
Introduction
T
Traditional biological nitrogen removal used in wastewater treatment systems involves two separate processes: aerobic autotrophic nitrification and anaerobic heterotrophic denitrification (Joo et al., 2005; Khardenavis et al., 2007). There are some problems in this system, for example, it requires separate treatment processes and strict condition control, and autotrophic nitrifiers are vulnerable to high concentrations of organic matter and ammonium (Liu et al., 2015). Hence, the operation cost is relatively high, and traditional nitrification can be carried out only after reducing the C/N ratio or diluting the wastewater.
Recently, some bacteria capable of combined heterotrophic nitrification and aerobic denitrification abilities have been isolated and intensively studied as potential microorganisms in biological nitrogen removal systems, such as Acinetobacter junii (Yang et al., 2015), Alcaligenes faecalis (Joo et al., 2005; Liu et al., 2015), Acinetobacter calcoaceticus (Zhao et al., 2010), Bacillus subtilis (Yang et al., 2011), Pseudomonas stutzeri (Zhang et al., 2011), Agrobacterium sp. (Chen and Ni, 2012a), and Rhodococcus sp. (Chen et al., 2012b). These special bacterial strains exhibit higher growth rates than autotrophs, can use organic substrates as carbon source and energy for denitrification, and perform simultaneous nitrification and denitrification in the same reactor. However, the performance of these bacteria suffers when pH, temperature, and/or salinity fluctuate.
Dinçer and Kargi (1999) reported that the nitrification and denitrification bioactivity was reduced significantly when salt was more than 2%. Yoshie et al. (2004) reported that salinity decreased the nitrite reductase gene diversity in denitrifying bacteria. Unfortunately, tannery industry wastewaters often contain a range of salts generated from soaking and unhairing operations (Lofrano et al., 2013). It is known that high salinity can cause cell plasmolysis and inhibit the enzyme activity of common microorganism, which lead to poor treatment efficiency (Dinçer and Kargi, 1999; Jang et al., 2013).
Previous research shows that the use of salt-tolerant bacterial strains can enhance wastewater biological treatment over a wide salinity range and improve the resistance or adaptability to salinity shock (Abou-Elela et al., 2010; Shi et al., 2012). Guo et al. (2013) isolated a halophilic strain (Halomonas campisalis ha3) that could carry out heterotrophic nitrification and aerobic denitrification. In their study, they found that Halomonas campisalis ha3 could remove inorganic nitrogen compounds simultaneously and grew well in the medium containing up to 20% NaCl. However, to our knowledge, there are few studies on biological nitrogen removal from tannery wastewater using heterotrophic nitrification–aerobic denitrification bacterium.
In this study, a novel halophilic heterotrophic nitrification–aerobic denitrification bacterium, strain CL1502, was isolated from marine sediments. The strain was characterized by the 16S rRNA gene sequence analysis and its ability of heterotrophic nitrification and aerobic denitrification was evaluated. In addition, its performance in actual tannery industry wastewater was investigated.
Materials and Methods
Mediums
All of chemicals used in this study were analytical grade without any further purification.
The basic medium (BM) used for bacteria isolation and heterotrophic nitrifying comprised the following ingredients (g/L): NH4Cl, 0.76; glucose, 5.0; K2HPO4·3H2O, 7.9; KH2PO4, 1.5; MgSO4·7H2O, 0.1; NaCl, 30; trace element solution, 2 mL; pH, 7.0-7.2; and salinity, 3% (w/v).
The aerobic denitrification medium (ADM-1) used for nitrate reduction included (g/L) the following: NaNO3, 1.22; glucose, 5.0; K2HPO4·3H2O, 7.9; KH2PO4, 1.5; MgSO4·7H2O, 0.1; NaCl, 30; trace element solution, 2 mL; pH, 7.0-7.2; and salinity, 3% (w/v).
The ADM-2 used for nitrite reduction included (g/L) the following: NaNO2, 0.98; glucose, 5.0; K2HPO4·3H2O, 7.9; KH2PO4, 1.5; MgSO4·7H2O, 0.1; NaCl, 30; trace element solution, 2 mL; pH, 7.0-7.2; and salinity, 3% (w/v).
Trace element solution contained (g/L) the following: EDTA, 50; ZnSO4·7H2O, 3.9; CaCl2, 5.5; MnCl2·4H2O, 5.06; FeSO4·7H2O, 5.0; Na2MoO4·2H2O, 1.0; CuSO4·5H2O, 1.57; and CoCl2·6H2O, 1.61.
Sample collection
The deep-sea sediment sample was collected using a TV-guided multicorer from the North Atlantic (26°13′N, 44°82′W; 3,667 m water depth), during cruise of DY125-26 R/V “Da-Yang-Yi-Hao” on October 29, 2009. The sediment sample was kept immediately at 4°C after being collected until use.
The tannery wastewater utilized in this experiment was taken from a tannery factory in Quanzhou, China. The characteristics of the wastewater were as follows: pH = 8.0, chemical oxygen demand (COD) = 4,500 mg/L, NH4+-N = 179 mg/L, and TN = 220 mg/L.
Isolation of bacteria
For the enrichment cultures, 10 g of marine sediment was added to a 500-mL Erlenmeyer flask containing 100 mL of BM. The flask was incubated at 30°C with shaking at 170 rpm for about 3 days. Then, 10 mL of cell suspension was used to inoculate 100 mL of fresh BM. These flasks were again incubated at 30°C for 3 days. After the second round, serially diluted cultures were spread onto the solid BM agar plates. After 1 day of incubation at 30°C, bacterial colonies with a distinct morphology were subcultured in fresh agar BM repeatedly for purification.
Identification and phylogenetic analysis of isolates
Colonies were assigned to the species level by amplification of rRNA gene region. Bacterial 16S rRNA genes were amplified with universal primers 27F (5′-AGA GTT TGA TC(C/A) TGG CTC AG-3′) and 1492R (5′-GGT TAC CTT GTT ACG ACT T-C′). Two microliters of whole cells from an exponentially growing culture of each isolate was used for amplification. Amplification program began with an initial cycle at 95°C for 102 min; followed by 25 cycles of 1 min at 95°C, 1 min at 54°C, and 2 min at 72°C. A final step of 10 min at 72°C was included to accomplish as described previously (Jin et al., 2012). Alignment of the 16S rDNA sequences, clustering by the neighbor-joining and analysis of the stability of clusters (bootstrap analysis with 1,000 replicaters) were performed with the software MEGA version 4.0 (Tamura et al., 2007).
Performance of heterotrophic nitrification and aerobic denitrification by CL1502 strain
To evaluate the ability of the isolated microorganism to heterotrophic nitrification and aerobic denitrification, 1 mL of preculture CL1502 strain was inoculated into 100 mL BM and ADM-1 and ADM-2 in 250-mL Erlenmeyer flasks, and was incubated aerobically at 30°C with shaking at 170 rpm. Samples were taken from the flasks periodically to determine the OD600, COD, NH4+-N, TN, NO3−-N, and NO2−-N.
To determine the effect of carbon source, sodium acetate, sodium citrate, sodium succinate, glucose, and sucrose were used as the carbon sources in the BM. To determine the effect of C/N ratio, the content of carbon source (glucose) in the BM was changed to adjust C/N ratio to 2, 10, 15, 20, and 25, respectively. To determine the effect of salinity, the salinity was varied at 1%, 3%, 4%, 5%, 6%, and 8%, respectively. To determine the effect of Cr(III), the different initial Cr(III) concentrations (0, 20, 60, 100, and 150 mg/L) were added into the BM. All of the above experiments were carried out with inoculation of 1 mL CL1502 strain in 100 mL BM and incubated at 30°C with shaking at 170 rpm.
Treatment of tannery wastewater by CL1502 strain
To investigate the application potential of the CL1502 strain in actual tannery industry wastewater treatment, 20 mL of CL1502 preculture was inoculated into 150 mL tannery wastewater in 500-mL Erlenmeyer flasks and was incubated aerobically at 30°C with shaking at 170 rpm. During treatment, samples were taken from the flasks periodically to determine the COD, NH4+-N, TN, NO3−-N, and NO2−-N.
Analytical methods
Concentrations of NH4+-N, TN, NO3−-N, NO2−-N, and COD were determined according to the standard methods (State Environmental Protection Administration of China, 2002). The OD600 was determined by using a UV-9600 spectrophotometer at 600 nm. All data shown in this study were the mean values of duplicate or triplicate experiments.
Results and Discussion
Identification of CL1502 strain
Colonies of strain CL1502 on the BM agar plate were white, opaque, and glossy, as well as a regular edge. The strain was gram negative and appeared as rod shaped with a size of ∼0.9–1.2 μm in length and 0.6–0.8 μm in width. The 16S rRNA gene sequence of CL1502 obtained was compared against the sequences available from GenBank database (www.ncbi.nlm.nih.gov/blast) as well as the EzTaxon database (www.eztaxon.org). Both comparative sequence analyses revealed that strain CL1502 belonged to the genus Serratia, with Serratia marcescens (similarity up to 99%) as its closest phylogenetic relative. In addition, a neighbor-joining phylogenetic tree of strain CL1502 also revealed that strain CL1502 and S. marcescens were in the same group (Fig. 1). The above results revealed that the screened strain was identified as S. marcescens CL1502.

Unrooted neighbor-joining phylogenetic tree of partial 16S rRNA sequences showing relationships between bacterial strain CL1502 and closely related reference organisms.
Heterotrophic nitrification–aerobic denitrification ability of strain CL1502
The growth and nitrogen removal characteristics of S. marcescens CL1502 in the heterotrophic nitrification medium were investigated and the experimental results are shown in Fig. 2. It could be seen from Fig. 2a that, when NH4+-N was provided as the sole nitrogen source, the NH4+-N concentration decreased significantly from 200 to 7.9 mg/L after 42 h of cultivation and the average NH4+-N removal rate was 4.57 mg/(L·h). Meanwhile, the cell growth achieved a stationary phase in 24 h as the OD600 increased from 0.15 to 1.22. Chen et al. (2012b) reported that Rhodococcus sp. CPZ24 removed NH4+-N (50 mg/L initial concentration) at a rate of 3.1 mg/(L·h). In the study of Padhi et al. (2013), the NH4+-N removal rate of Klebsiella pneumoniae CF-S9 strain was 4.3 mg/(L·h) at an initial concentration of 120 mg/L NH4+-N.
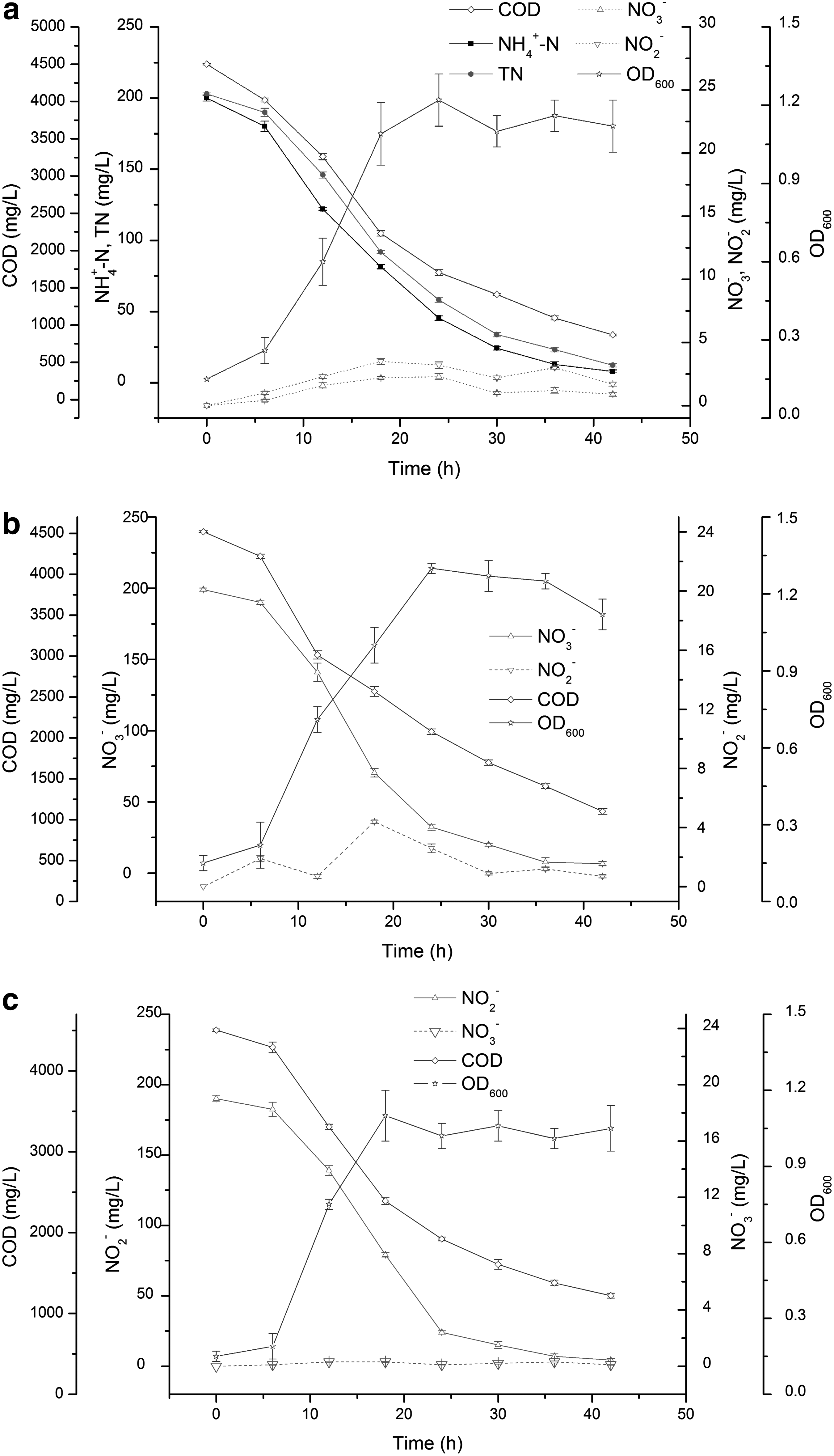
Characteristics of heterotrophic nitrification and aerobic denitrification by CL1502 strain when ammonium
In addition, the COD concentration also decreased rapidly with the growth of S. marcescens CL1502, and ∼80.7% of the initial COD was reduced during 42 h of cultivation. The COD removal profile, similar to that of NH4+-N, indicated that the consumption of organic matter and the degradation of NH4+-N occurred simultaneously (Shi et al., 2013). Furthermore, during the whole process of batch tests, only a few of nitrification products nitrate and nitrite were observed at maximum accumulation concentrations of 2.3 and 3.5 mg/L, respectively. In addition, it also could be found that neither ammonium removal nor the growth of CL1502 strain occurred in the absence of organic carbon (data not shown).
To confirm the ability of aerobic denitrification by S. marcescens CL1502 strain, nitrate and nitrite were utilized as the nitrogen sources, respectively, and the results are depicted in Fig. 2b, c. As shown in Fig. 2b, after 42 h of inoculation, the concentration of NO3−-N decreased rapidly from 199 to 6.6 mg/L, corresponding to a removal efficiency of 96.7%, and the average NO3−-N removal rate was 4.6 mg/(L·h). Meanwhile, the removal of COD and growth of CL1502 strain were accompanied by the removal of NO3−-N. Approximately 75.6% of initial COD was degraded after 42 h of cultivation. The cell growth achieved the maximum phase at 24 h with an OD600 value of 1.3. It also could be seen that NO2− slightly accumulated (maximum concentration of 4.4 mg/L) in this process. As shown in Fig. 2c, the removal profile of NO2−-N was similar to that of NO3−-N, but there was no noticeable NO3−-N accumulation. After 42 h of inoculation, the NO2−-N removal efficiency and removal rate were 97.7% and 4.42 mg/(L·h), respectively. The COD concentration was decreased from 4,505 to 1,221 mg/L, corresponding to a removal efficiency of 72.9%. These experimental results indicated that S. marcescens CL1502 strain had a capacity of simultaneous heterotrophic nitrification and aerobic denitrification.
Effect of different carbon sources on heterotrophic nitrification by S. marcescens CL1502 strain
Carbon compounds usually serve as energy and electron source for heterotrophic nitrifying bacteria. The effect of different carbon compounds that serve as carbon source on heterotrophic ammonium removal by S. marcescens CL1502 strain was investigated, and the result is depicted in Fig. 3. It could be observed that, when sodium acetate, sodium citrate, sodium succinate, glucose, and sucrose were used as the carbon sources, after 42 h of inoculation, the NH4+-N removal efficiencies were 76.6%, 61.7%, 90.0%, 96.9%, and 85.2%, respectively, and the TN removal efficiencies were 75.0%, 58.9%, 89.0%, 94.6%, and 82.5%, respectively. This implied that the glucose was the most suitable carbon source for heterotrophic ammonium removal by S. marcescens CL1502 strain. Chen et al. (2015) also found that a heterotrophic nitrifying bacterial strain (Anoxybacillus contaminans HA) exhibited the best NH4+-N removal ability when glucose was used as the sole carbon source.

Effect of different carbon sources on heterotrophic ammonium removal by CL1502 strain.
Effect of C/N ratio on heterotrophic nitrification by S. marcescens CL1502 strain
NH4+-N removal by S. marcescens CL1502 strain was affected not only by organic carbon sources but also by the ratio of C/N. Thus, the effect of C/N ratio on heterotrophic ammonium removal by S. marcescens CL1502 strain was determined, and the results are shown in Fig. 4. It could be seen that the removal efficiencies of NH4+-N and TN were increased first and then kept constant with the increase of C/N ratio. When the C/N ratio was increased from 2 to 15, the NH4+-N removal efficiency increased from 50.1% to 94.9% and the TN removal efficiency increased from 37.2% to 93.9%.
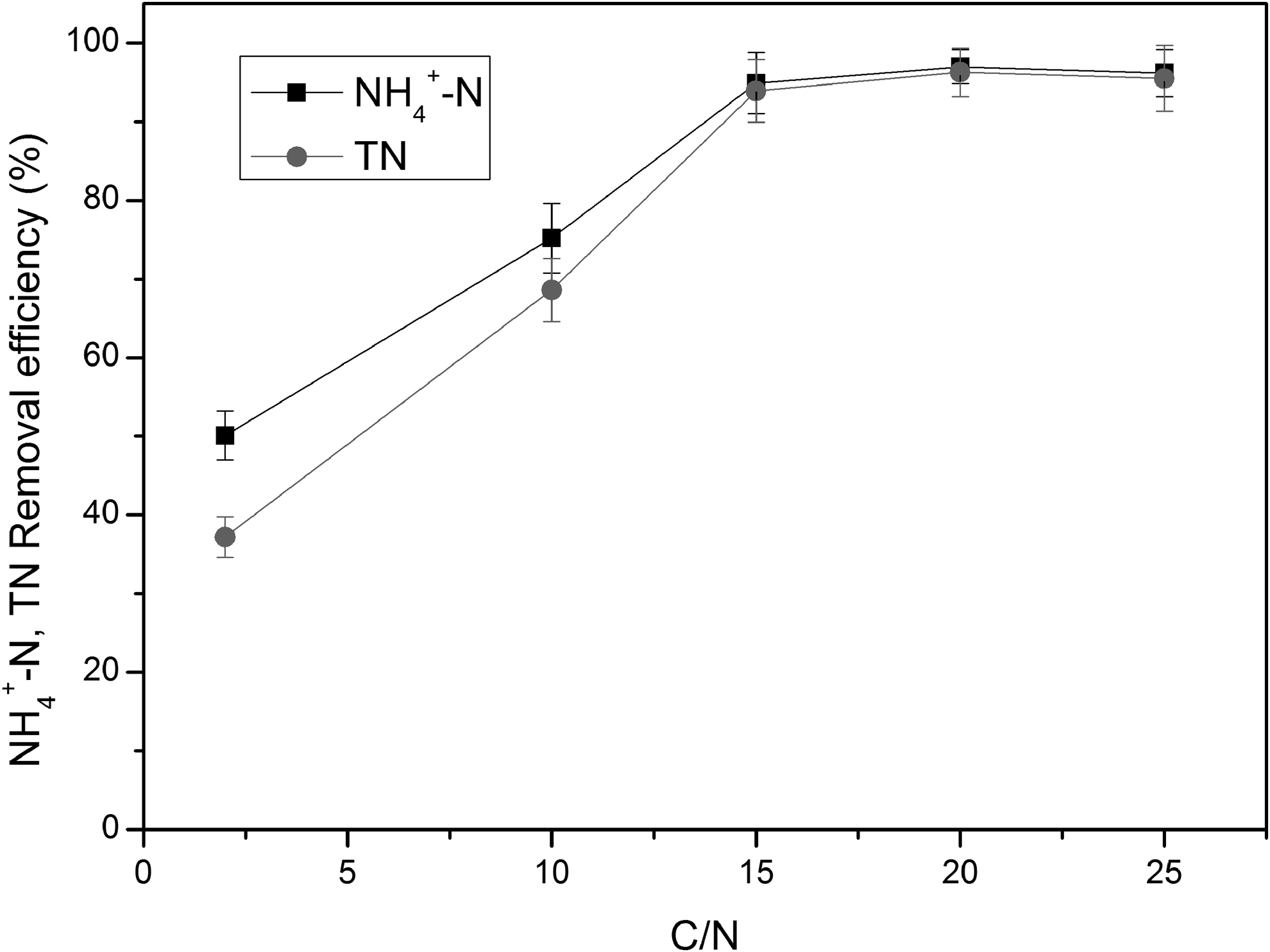
Effect of different C/N ratios on heterotrophic ammonium removal by CL1502 strain.
In addition, it should be noticed that the TN removal efficiency was smaller than NH4+-N removal efficiency, and the removal efficiencies of NH4+-N and TN were low at a low level of C/N ratio. The reason for this phenomenon might be that the exhaustion of the carbon source caused the insufficient carbon supply for both microbial growth and electron donor for denitrification (Taylor et al., 2009; Yang et al., 2016). When the C/N ratio was further increased to 20, the removal efficiencies of NH4+-N and TN remained stable at around 97% and 96%, respectively.
Effect of salinity on heterotrophic nitrification by S. marcescens CL1502 strain
Effect of salinity on heterotrophic ammonium removal by S. marcescens CL1502 strain is demonstrated in Fig. 5. It was observed that the NH4+-N removal efficiency could be stabilized at a relatively high level when the salinity was below 5%. In addition, when the salinity was more than 5%, the removal efficiency of NH4+-N decreased significantly. This might be due to the high salinity that caused cell plasmolysis and activity loss of microorganism (Uygur and Karg, 2004). After 42 h of cultivation, the NH4+-N removal efficiencies were 97.0%, 96.1%, 96.7%, 95.2%, 83.6%, and 51.7% when the salinity was 1%, 3%, 4%, 5%, 6%, and 8%, respectively.
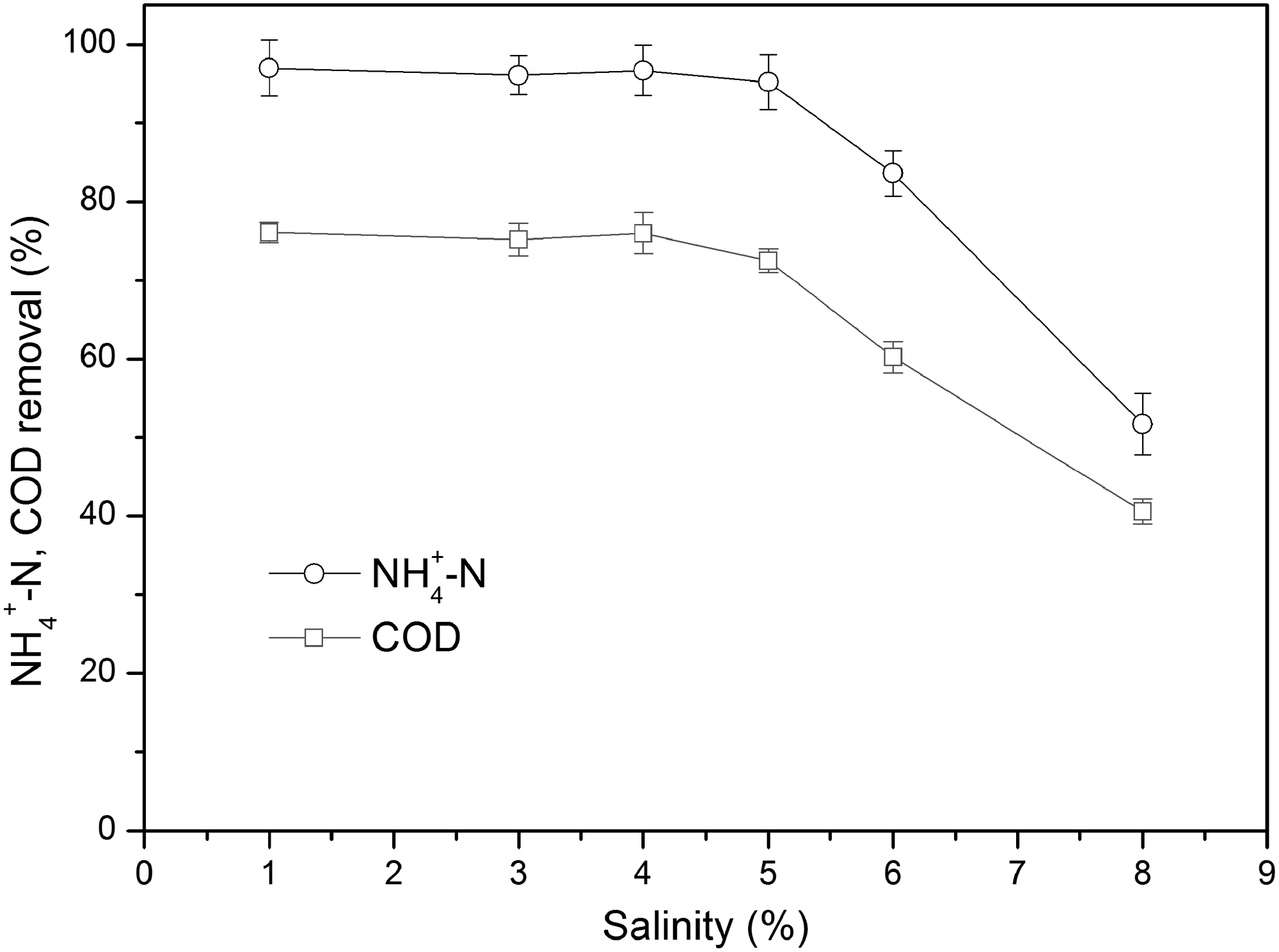
Effect of salinity on heterotrophic ammonium removal by CL1502 strain.
In addition, it also could be observed that the COD removal showed similar trend of NH4+-N removal. After 42 h of cultivation, the COD removal efficiencies were 76.1%, 75.2%, 76.0%, 72.5%, 60.2%, and 40.6% when the salinity was 1%, 3%, 4%, 5%, 6%, and 8%, respectively. This experimental result indicated that the S. marcescens CL1502 strain had a high tolerance to salinity.
Effect of Cr(III) on heterotrophic nitrification by S. marcescens CL1502 strain
It is known that chromium is one of the major contaminants in the tannery effluent, and hence, the effect of Cr(III) on heterotrophic ammonium removal by S. marcescens CL1502 strain is presented in Fig. 6. From Fig. 6a, it is evident that in comparison to the control [no addition of Cr(III)], the heterotrophic NH4+-N removal was not affected by the addition of Cr(III) at concentrations up to 60 mg/L, while a slightly negative impact was observed at Cr(III) concentration of 100 mg/L. When the Cr(III) concentration was further increased to 150 mg/L, the NH4+-N removal efficiency decreased up to ∼10%. When the initial Cr(III) concentrations were 0, 20, 60, 100, and 150 mg/L, after 42 h of reaction, the NH4+-N removal efficiencies were 96.7%, 97.0%, 96.3%, 91.9%, and 86.9%, respectively.
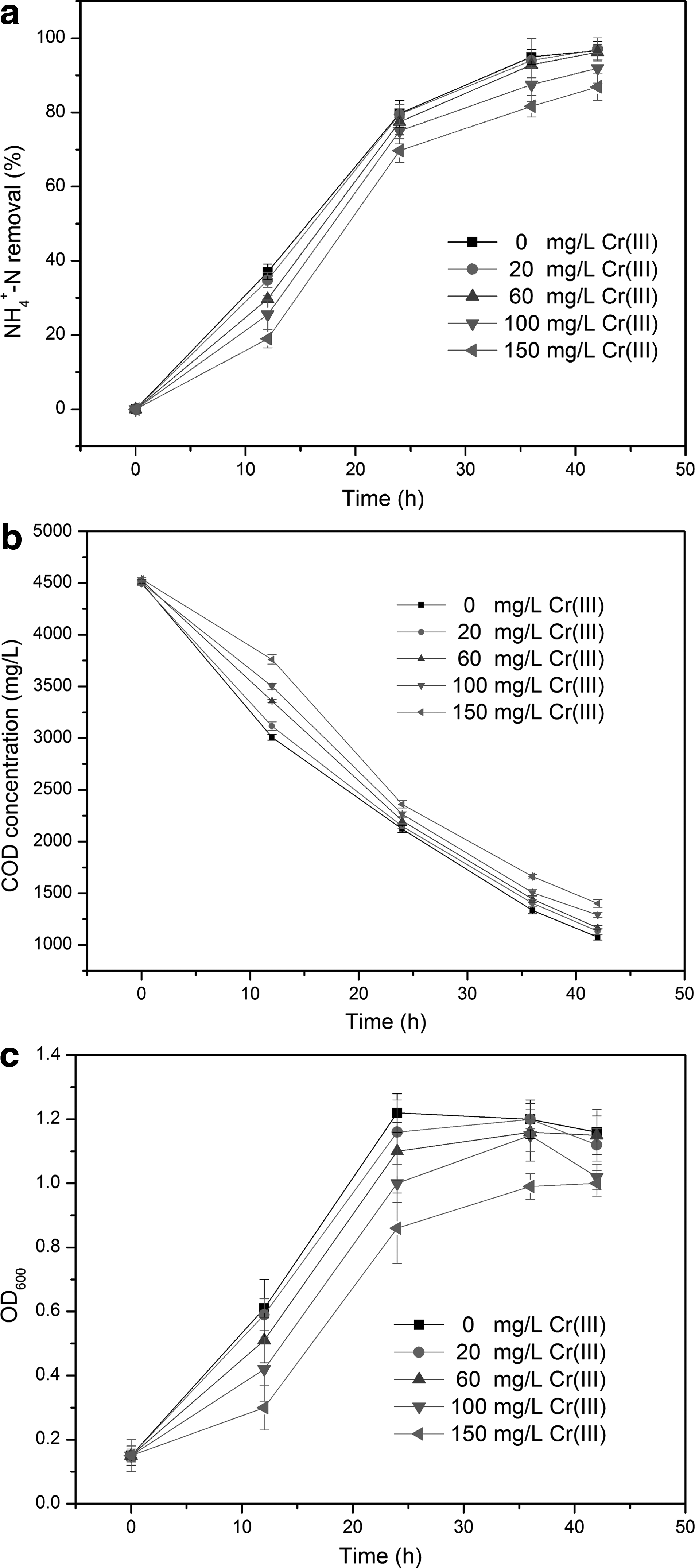
Effect of different initial concentrations of Cr(III) on heterotrophic ammonium removal by CL1502 strain.
In addition, it could be seen from Fig. 6b that the COD removal was slightly inhibited by the addition of Cr(III). Fig. 6c shows that increasing the concentration of Cr(III) from 0 to 60 mg/L had little effect on the growth of CL1502 strain. When the concentration of Cr(III) was increased to 150 mg/L, the growth of CL1502 strain showed a lag phase. These experimental results indicated that S. marcescens CL1502 strain had a good tolerance to Cr(III).
Bioremediation of tannery wastewater by S. marcescens CL1502
The ability of S. marcescens CL1502 to remove nitrogen and COD from actual tannery industry wastewater was investigated (Table 1). It could be observed from Table 1 that S. marcescens CL1502 strain had a good treatment performance in COD, NH4+-N, and TN removal from actual tannery wastewater. After 3 days of cultivation, the removal efficiencies of COD, NH4+-N, and TN were 63.1%, 76.7%, and 72.9%, respectively. This experimental result indicated that S. marcescens CL1502 strain had a great potential promising for tannery industry wastewater treatment.
COD, chemical oxygen demand.
Conclusions
This work focused on the study of nitrogen removal characteristics by a novel heterotrophic nitrifying bacterium, S. marcescens CL1502, isolated from deep-sea sediment. The S. marcescens CL1502 possessed excellent heterotrophic nitrification and aerobic denitrification capacity with NH4+-N removal rate of 4.57 mg/(L·h), NO3−-N removal rate of 4.60 mg/(L·h), and NO2−-N removal rate of 4.42 mg/(L·h) when ammonium, nitrate, and nitrite were used as the nitrogen sources, respectively. The optimum carbon source for heterotrophic NH4+-N removal by S. marcescens CL1502 was glucose. The higher C/N ratio was favorable to the removal of NH4+-N and TN. The S. marcescens CL1502 had a good tolerance to salinity (1–5%) and Cr(III). Treatment of actual tannery wastewater by CL1502 resulted in COD, NH4+-N, and TN removal efficiencies of 63.1%, 76.7%, and 72.9% within 3 days.
Footnotes
Acknowledgments
This work was supported by the Natural Science Foundation of Henan Province (No. 132300410139) and the Science and Technology Research Key Project of Education Department of Henan Province (No. 13A610528).
Author Disclosure Statement
No competing financial interests exist.