
Abstract
Abstract
A combination of methods was applied in this study to evaluate the removal of pentachlorophenol (PCP) from an Indian agricultural soil. Treatment involved PCP-resistant microorganism Vibrio sp. (SK1); their activity was combined with the adsorbing capacity of biochar. First, PCP-resistant microorganism was used to transform PCP at batch level and further extended to farmland soil. Experimental results indicated that bacterial activity was found to have pronounced positive effect on PCP removal, and decreased the PCP level up to 75% of soil spiked with 100 mg/kg, compared to control within 1-week period. Remaining PCP in soil was stabilized probably by sorption using biochar prepared from Pistia stratiotes; biochar was added at concentrations of 1%, 3%, and 5%. We observed that the 5% biochar strongly influenced the stabilization of PCP in the soil. 16s rRNA gene analysis indicated that bacterial strain belongs to Vibrio sp., which was not reported earlier for the PCP degradation. In our study, a parameter estimation method was used in mathematical models along with kinetic models. By using the nonlinear graphical regression method with the logistic growth curve, Monod, Contois, Moser, and the Tessier models, kinetic parameters were estimated.
Introduction
P
Chlorophenols and their derivatives are highly toxic to living organisms due to their carcinogenic, mutagenic, and cytotoxic properties. The International Agency for Research on Cancers and World Health Organization declared that chlorophenols have the potential to induce cancer in humans (Igbinosa et al., 2013). Several chlorophenols are in the list of priority pollutants by the United States Environmental Protection Agency (USEPA) as well.
Different physicochemical treatments are available for remediation of PCP; however, compared to other methods, bioremediation is largely favored for PCP cleansing due to its sustainable nature. The degradation of a particular compound required a particular microbe and its enzyme, which depends upon the chemical structure and the presence of the enzyme systems in bacteria. Therefore, knowledge regarding chemicals such as structure, environmental properties, toxicity, risk to human, distribution, and obvious have idea about efficient microbial removal (microbes, enzymes, genes, proteins) which can improve bioremediation process. PCP degradation mechanism involved the following: oxygenolysis, hydroxylation, or reductive dehalogenation (Field and Sierra-Alvarez, 2008).
Aerobic and anaerobic microorganisms were reported to degrade PCP under a variety of conditions and at different concentrations. Microbial species or strain like Sphingomonas sp., Pseudomonas sp., Arthrobacter sp., Mycobacterium sp., Flavobacterium sp., Serratia sp., and Bacillus sp. showed various rates and extent of PCP degradation (Singh et al., 2007; Bosso and Cristinzio, 2014). Biodegradation is a cost-effective and environment-friendly option for the treatment of various hazardous pollutants that may exist in the solid, aqueous, or gaseous form, but they also have certain limitations (Karn et al., 2011). Microorganisms are able to remove PCP at a certain extent (fixed concentration), after that, microbes are unable to transform PCP even in laboratory-level experiments, which provide all the necessities of physical condition. The removal efficiency decreases eventually due to the varying environmental conditions in the field-level or in situ remediation (Karn and Geetanjali, 2014). Therefore, it is needful to find a two step process first remove by using efficient isolate and second application of biologically or biochemically or chemically derived material for the complete removal of contaminant from the soil during field-level remediation.
Thermochemical conversion of biomass/waste has been considered a feasible option for soil remediation. After thermochemical conversion of biomass, biochar is formed as a byproduct of pyrolysis, its optimization and application is necessary to prepare the entire process economically viable. Biochar can also be used as a precursor of value-added activated carbons. By steam activation and fast pyrolysis, biochar greatly enhances the surface area, sorption capacity, and porous structure for water contaminants (Lima et al., 2010). Lehmann and Rondon (2006) also proposed the application of biochar (BC) to the degraded soil for sustainable development. BC enhances plant growth, retains nutrient, improves physical, chemical, and biological properties, and performs as soil conditioner. BC influences the porosity and consistency through changing the bulk surface area, reducing the contaminant from the polluted soil (Downie et al., 2009). PCP has a negative impact on the soil and water to human health; therefore, it is necessary to remove the PCP from contaminated soil completely or up to the safe limit.
Kinetic parameter identification technique was conducted under controlled conditions by using a nonlinear multiple response regression algorithm (Marquardt, 1963; Lin and Miller, 2004; Sudipta and Somnath, 2010; Eswari and Chimmiri, 2013). The integration of the set of differential equations, for the calculation of the parameters used to adjust the experimental data, was performed with the aid of the fourth-order algorithm of Runge–Kutta. Parametric identification is indicated when there is good agreement between the experimental values and those supplied by the model. This challenge is to develop innovative and cost-effective solutions to decontaminate polluted sites, to make them safe for human habitation and consumption, and also to protect the ecosystem that can support healthy life.
Therefore, in this investigation, PCP was first removed efficiently by a newly isolated organism, which is not reported earlier for PCP degradation. Kinetic and mathematical modeling has also been introduced to predict the effect of remediation of PCP-contaminated soil. Further addition of biochar to the bacteria-treated soil was done for its complete removal of PCP. The benefits of these approaches used are functional for PCP-contaminated soil and the development of sustainable technology.
Materials and Methods
Sample enrichment and isolation of microbe
Sample was collected from a river at Yamunanagar (30.133°N, 77.288°E), Haryana, India, in sterile conical flasks and stored at 4°C. In this place, all the waste from the cities and industries are disposed off. PCP-tolerant bacterial strains were isolated on a mineral salt medium (MSM) (Sharma et al., 2009) using an enrichment method supplemented with 50 mg/L PCP (0.185 mM) as the sole carbon source for energy. The MSM contain the following constituents (in mg/L): KH2PO4, 800; Na2HPO4, 800; MgSO4.7H2O, 200; CaCl2. 2H2O, 10; NH4Cl, 500; plus 1 mL of trace metal solution, which includes FeSO4.7H2O, 5; ZnSO4.7H2O, 4; MnSO4.4H2O, 0.2; NiCl.6H2O, 0.1; H3BO3,0.15; CoCl2.6H2O, 0.5; ZnCl2, 0.25; and EDTA, 2.5. The pH of the medium was adjusted to 7.5. Bacteria were isolated by the serial dilution method; further repeated subculture was done on nutrient agar plates to get the purified culture. Colonies that appeared after incubation at 37°C for 48 h were selected for further screening. The potent isolate were screened for the degradation studies and one of the efficient isolate was selected and designated as SK1 for further studies. PCP was added to the medium after autoclave. Synthetic PCP was purchased from Sigma Aldrich chemicals. All solutions were prepared in sterile Milli-Q water (Millipore direct Q3, Bangalore, India). PCP was dissolved in small amounts of methanol (0.2%) and purified water (Mili1-Q water).
Growth, degradation, and chloride release analysis by using SK1
The bacterial strain SK1 was inoculated into 250-mL Erlenmeyer flasks containing 50 mL of MSM supplemented with different PCP concentrations in mg/L: 50 (0.185 mM), 100 (0.37 mM), 200 (0.75 mM), 400 (1.5 mM), and 600 (1.8 mM) of PCP. Next, the flasks were incubated under shaking conditions by fixing temperature at 37°C for a168-h duration. The growth of SK1 was measured from culture filtrate collected at different time intervals and was determined by measuring the optical density at 600 nm.
Before starting PCP removal experiment, inoculation was prepared and degradation was started by inoculating 1% inoculum (105 colony-forming unit [CFU]/mL) of the bacterial strain SK1 in 250-mL Erlenmeyer flasks containing 50 mL of MSM medium added with 100 mg/L (0.37 mM) of PCP. The flasks were incubated at 37°C under shaking conditions (120 rpm) up to 168 h. Growth of the bacterial cells was observed by measuring the optical density at 600 nm and the PCP degradation was analyzed by gas chromatography (GC). Chloride ion released in MSM was estimated at every 24-h interval up to 168 h from culture filtrate. Chloride ion concentration was further analyzed with an Orion ion analyzer mode l940 using calibrated selective chloride ion electrode. The electrode was calibrated by adding known concentrations of sodium chloride standards to the MSM.
Degradation studies of PCP in soil
First, we collected farmland soil and characterized for different parameters such as pH, C, H, N, P, S, and O analysis. One gram of soil sample was taken and oven dried at 105°C for 24 h. The sample was ground to a particle size of about 0.1 mm; further, dried soil sample (10–50 mg) was taken for C, H, N, and O analysis using an Elemental analyzer (Thermo Scientific). Next, farmland soil was sieved and sterilized to remove entire microorganism. Degradation experiment was carried out in 250-mL flask containing 200 g soil mixed with PCP from stock solution of 250 mg/L (100 mg/kg PCP prepared in soil because the initial PCP concentration in the soil was negligible). Approximately, 80 mL of the PCP solution was mixed with 200 g of soil and kept for further 2 days; SK1 (5%) was inoculated and mixed properly, and moisture required was maintained with deionized water. For each experiment, we kept three beakers. Un-inoculated control was conducted simultaneously; next, the soil container/beaker was incubated at 30°C for 2 weeks by maintaining the moisture of the soil between 60% and 65%. Survival and growth of SK1 were monitored by CFU/mg of soil. After 2 weeks, experiment was terminated and PCP was extracted from the soil by the method described in National Council for Air and Stream Improvement [NCASI] CP-85.01 method, 1986.
Preparation and application of biochar into soil
Biochar is a carbon-rich material, prepared by thermal decomposition of biomass in the absence of oxygen (O2) at a relatively low temperature (<700°C). The biochars used in this study were obtained from Pistia stratiotes leaves (about 200 g of sample was placed in a ceramic cup) through a pyrolysis method. Methods for the preparation of biochar have been described previously (Cao and Haris, 2010). Briefly, the dried Pistia stratiotes leaves were put in a ceramic cup and heated in a Muffle Furnace under O2-limited condition at 650°C for 5 h. The solid residue left after heating was C rich and called as biochar (Lehmann, 2007). The mineralogical composition of biochar was analyzed using scanning electron microscope using Zeiss EVO 18 Special edition for imaging and elemental analysis. Now the biochar was applied to the bacteria-treated soil separately for the stabilization of PCP. For this, we added biochar at the concentration of 1%, 2.5%, and 5% into the soil sample and mixed properly and incubated for 7 days at temperature 30°C ± 2°C and soil moisture was about 75%. About 2 mm size of biochar was applied to the soil. Furthermore, PCP was extracted from the soil using NCASI CP-85.01 method, 1986, and the concentration was evaluated from the soil.
GC analysis
PCP was extracted and analyzed by using NCASI CP-85.01 method, 1986, with a little modification, First, for extraction of PCP from the soil, we sonicated the soil sample in a Sonicator (SG-25) (Roop Telsonic Ultrasonix Ltd., India) by keeping in ice (power intensity 3, 7.4 W/cm2; frequency 24 kHz). Soil sample were kept in ice and sonicated for 10 min with 1.5 min burst at a certain interval of 5 m by keeping in ice. Next, 1.3 mL K2CO3 (pH 11.5) was added, further sonicated for 2 m with addition of 1.5 mL of acetic anhydride, and sonicated for 2 min again by adding 0.5 mL acetic anhydride. Next, 5 mL of hexane was added, sonicated, centrifuged, and extracted with hexane. After centrifugation at 10,000 rpm for 15 min, the supernatant was collected and used to analyze the chlorophenols.
GC analysis was performed with Nucon GC-5765 (Centurion Scientific, India) in electron capture detection mode. A capillary column DB-5 (30 m length 0.025 mm i.d.) was used and initial temperature was 50°C (for 2 min) further increased to 10°C/min to 280°C where it was kept for 10 min. Helium as the carrier gas was used at a flow of 1.2 mL/min. The samples were analyzed in split less mode at an injection temperature was fixed 250°C and temperature of detector was 280°C. The injected volume was 0.1 μL.
Molecular identification and phylogenetic analysis
For identification, whole DNA was isolated according to standard protocol; furthermore, 16S rRNA gene was amplified and sequenced as described in Karn et al. (2010a). Next, 16S rRNA gene sequence was checked to NCBI (www.ncbi.nlm.nih.gov) by using BLASTN program (Altschul et al., 1997) and also compared to Ribosomal Database Project-II (RDP-II) (Cole et al., 2005). A further sequence was aligned by CLUSTALW program (Thompson et al., 1997). The evolutionary distance was measured by Kimura 2; next, phylogenetic tree was constructed by the neighbor-joining method using MEGA5.0 package. Bootstrap analysis was based on 1,500 resamplings (Tamura et al., 2007).
Kinetic growth models and parameter estimation
The kinetic model was prepared on the basis of three rate equations: biomass growth, substrate utilization, and product formation. Model described the rate of increase in biomass as a function of biomass only. Thus
where X is the biomass and μ is the specific growth rate.
Logistic growth rate equation
Maximum specific growth rate (Logistic-μmax) estimated by using Logistic growth curve is given in Equation (2), and then, the corresponding specific growth rate values were calculated by using Logistic growth curve mentioned in Equation (2).
Monod kinetic model
Monod kinetic model is considered one of the unstructured models, which are dependent on substrate concentrations. The Monod equation has been widely used in the literature to describe microbial growth kinetics. It describes the relationship between the microbial specific growth rate, p, and the limiting substrate concentration, S, as:
The linearized Monod equation is as follows:
So that, the first rate equation was obtained as follows:
Generally, Monod kinetics is valid when growth and population density are low. The constants (Monad)-μmax and Ks are referred to as the maximum specific growth rate and the half saturation coefficient, respectively, and are intrinsic parameters for a given microbe substrate system. It follows from Equation (1) that for any microbe–substrate system described by the Monod equation, the specific growth rate is a function of the limiting substrate concentration.
Contois model
It is another unstructured model with a slight modification of Monod model is a Contois kinetic model depends on two terms such as substrate and cell concentrations. The following equations are nonlinear and linear forms of Contois model:
where Ksis is the Contois kinetic constant and Contois-μmax, is referred to as the maximum specific growth rate. The effective substrate concentration is proportionally related to the cell growth, where μ is inversely related to the cell growth at high cell density.
Moser model
Moser kinetic model is strictly related to substrate concentration. The following equations are nonlinear and linear forms of Moser model:
where Moser-Ksis is the saturation kinetic constant. Moser-μmax is referred to as the maximum specific growth rate.
Tessier model
Tessier kinetic model is strictly related to substrate concentration. The following equations are nonlinear and linear forms of Tessier model.
where Tessier-Ksis is the saturation kinetic constant. Teisser-μmax is referred to as the maximum specific growth rate.
Mathematical modeling (theoretical)
Assuming constant temperature, the material balances of the biomass, product (PCP degradation), and substrates (PCP leftover) can be written in the following form:
In the above equations, the notation X, S, and P denote biomass, substrate (PCP remaining), and product (PCP degradation), respectively. Equation (2) is classical growth equation. In accordance with the quasi-steady-state assumption, constant specific uptake and excretion rates are assumed during the exponential growth phase of metabolites. The experimental data of substrate (PCP remaining) and product (PCP degradation) as well as the biomass obtained (during the exponential growth phase) have been reported in the previous section. The classic study of Luedeking and Piret (1959) on the PCP degradation by Vibrio sp. indicated that the product formation kinetics combined both growth- and nongrowth-associated contributions.
The reduced form of Luedeking–Piret product is
Finally, substrate utilization kinetics may be expressed as the reduced form of Luedeking–Piret substrate, which is expressed as follows:
The model (Luedeking–Piret substrate and product) are used for fitting the PCP degradation data, especially, how substrate is involved in the product formation can be interpreted. The model parameters were incorporated in the models and integrated using Runge–Kutta method.
Kinetic modeling and parameter estimation
The logistic model which was shown in Equation (2), solved and the kinetic parameters estimated by nonlinear graphical regression method. The Monod, Contois, Tessier, and Moser models were fitted for the growth in the process of PCP degradation. The model parameters were first evaluated by solving the Equations (3)–(10). These values were then used to calculate the kinetic parameters. Substrate utilization (PCP remaining) and product formation (PCP degradation) parameters were calculated from Equations (14) and (15), respectively. These model equations were written in linear form as in Equations (11)–(15) for the determination of the constants.
Monod kinetic model: Equation (3) represents the nonlinear model. The graph was plotted between 1/s versus 1/μ by using linearized Equation (4). The Lineweaver–Burk plot was used for determination of kinetic parameters. The constant μmax was determined by intercept and Ks were obtained from slope of the plot. Contois kinetic model Equations (6) and (7) are the nonlinear and linear forms of Contois model. The graph was plotted between x/s versus 1/μ by Lineweaver–Burk plot, where constants are determined from slope and intercept. Moser kinetic model: Moser kinetic Equations (8) and (9) represent the nonlinear and linear forms. The plot of 1/s2 versus 1/μ was used to obtain the constants by using the slope (Ks/μmax), the intercept (1/μmax), and the exponent of substrate concentration for n = 2. The parameters are shown in Table 1. Luedeking–Piret equation: substrate utilization (PCP remaining) and product formation (PCP degraded) were determined by using linearized Equations (14) and (15). The product formation constant α, β and substrate use parameters γ and δ were determined by using plot between μ versus Qp and μ versus Qs in (Fig. 5). The parameters are in (Table 1) PCP degradation kinetic studies. The kinetic data also showed that substrate consumption was due to both growth and nongrowth metabolic activities. The kinetic parameters estimated by the nonlinear graphical regression method by logistic growth curve, Monod, Contois, Moser, and the Tessier kinetic model obtained are given in Table 1. To determine product, Equations (11)–(13) were solved numerically using a fourth-order Runge–Kutta algorithm with a time step of 1 day. The model obtained has proven very effective in describing the kinetic behavior for PCP degradation and PCP leftover, as a function of cellular growth. Better fitting was observed with logistic growth curve method when comparing with the remaining kinetic models. Hence, the corresponding logistic model with estimated parameters is shown in Fig. 6. Since the logistic model was fitted better, PCP degradation and leftover exponentially depend on the substrate depletion and growth.

Luedeking and Piret kinetic parameter estimation of Vibrio sp. (SK1).
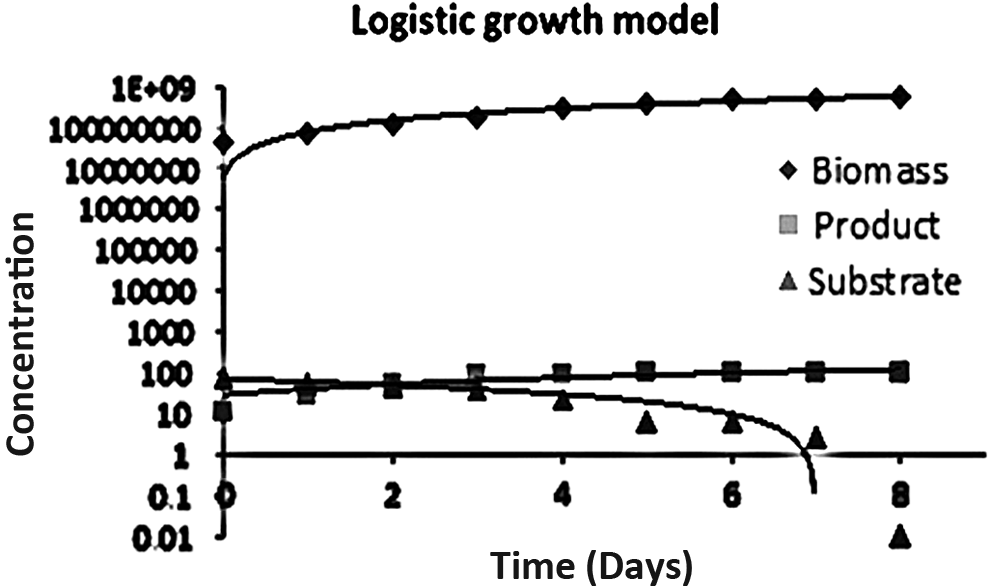
Logistic growth model of Vibrio sp. (SK1).
μmax = maximum specific growth rate; KS = saturation constant; α = growth-associated parameter for the product formation; β = nongrowth-associated parameter for the product formation; γ = growth-associated parameter for the substrate depletion; δ = nongrowth-associated parameter for the substrate depletion.
Data analysis
Experimental data were subjected to analysis of variance, and the mean differences were compared by Tukey–Kramer Multiple Comparison Test (p < 0.05) using GraphPad Prism (v 4.03) software. Three replicates were maintained for each treatment.
Results and Discussion
PCP removal by Vibrio sp. (SK1) in batch experiment
Several microorganisms showed excellent ability to tolerate and remove PCP, by using two mechanisms: to avoid PCP the toxicity excluding it from the cell and after that converting it into a nontoxic compound or by using PCP as the carbon or energy source (McAllister et al., 1996). PCP is more toxic in nature compared to other chlorophenols because the toxicity of chlorophenols increases with relative degree of chlorination (Reineke and Knackmuss, 1988). Present strain was able to grow and use PCP as an energy source and also convert into less toxic compounds by releasing chloride into the medium. The growth of bacterial strain was observed in the PCP-supplemented medium and we observed a significant increase in growth up to 6 days, further attaining a stationary phase, and decreased thereafter. The bacterial growth curve with correspondence to liberation of chloride ion was determined and shown in Fig. 1a, discovered that the strain achieved good growth with simultaneous liberation of chloride ion into the medium. The PCP was completely mineralized at 100 mg/L (0.37 mM) in 6 days of incubation (Fig. 1b). The liberation of chloride ion in the culture filtrate can be considered the result of mineralization of PCP. It was observed that growth of Vibrio sp. (SK1) significantly reduced as the concentration of PCP increased in the MSM. Up to 200 mg/L (0.75 mM), 95% removal of PCP was observed and slightly decreased at 400 (1.5 mM), which were about 80%, moreover, at 500 mg/L (1.8 mM), removal was about 75%, shown in Fig. 2a, b. Previously, Karn et al. (2010b, 2011, 2014) reported that Pseudomonas sp., Bacillus sp., Kocuria sp., and Enterobacter sp. also released chloride ion and produced biomass using PCP as a carbon source. Verma and Singh (2013) isolated and identified Brevibacterium casei (TVS-3) and observed the release of 900 mg/L chloride ions by using 720 mg/L PCP in 168 h.
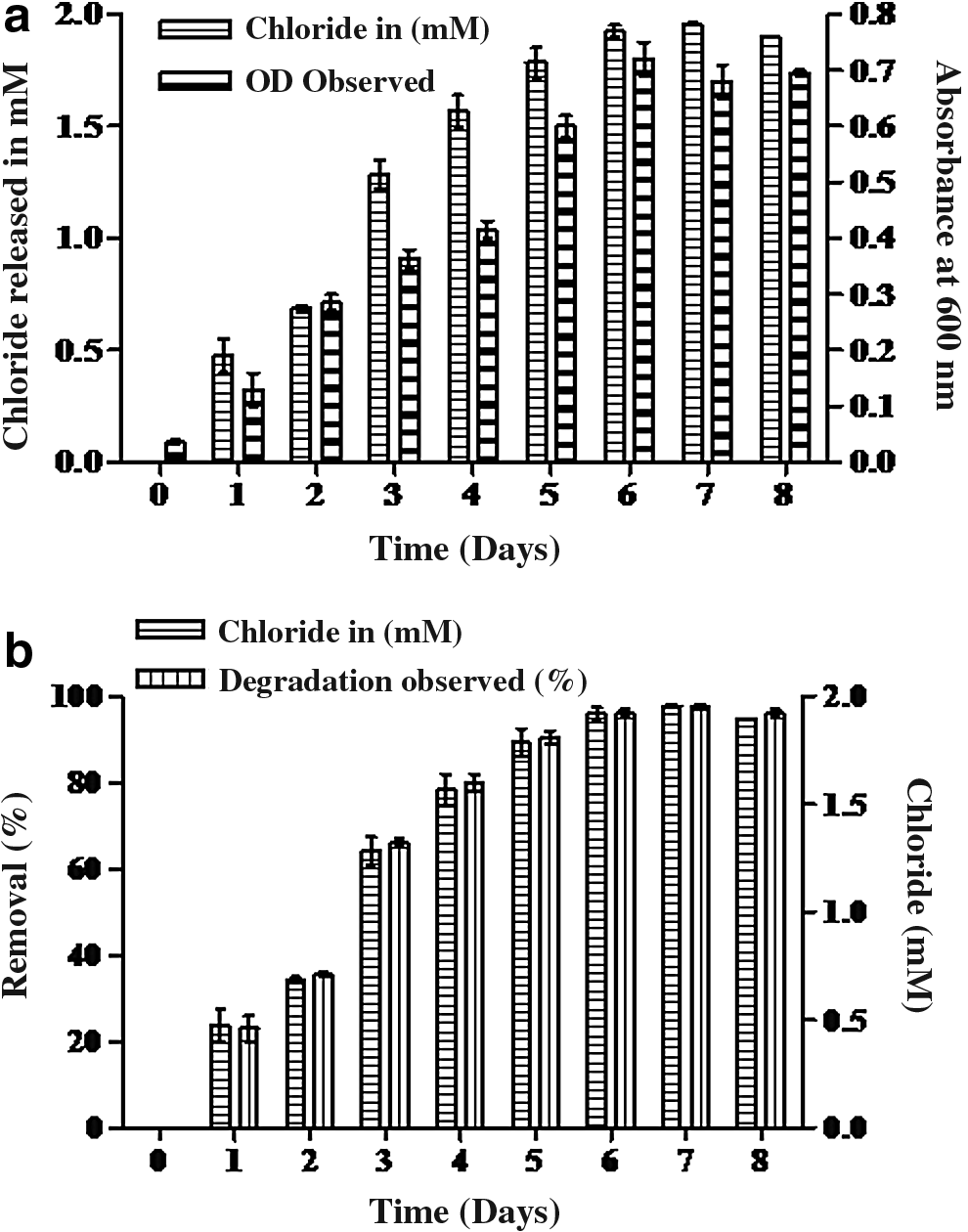

Different kinds of bacteria are known, which can use chlorophenols as carbon and energy sources under aerobic conditions. Like Lee et al. (1998) observed about Pseudomonas sp. (Bu34) grown up to 4,000 mg/L of PCP, Shah and Thakur (2002) reported 72% of transformation of PCP by Pseudomonas fluorescence (TE3) at 96 h when grown with 100 mg/L PCP. Sharma and Thakur (2008) reported that Pseudomonas aeruginosa (PCP2) removed 60% of PCP within 96 h of incubation. Singh et al. (2007) found that Serratia marcescens from pulp and paper mill waste was able to degrade 90.3% of PCP within 168 h when grown with 300 mg/L of PCP. Recently, Verma and Singh (2013) observed that Brevibacterium casei (TVS-3) degraded 1,000 mg/L PCP. This bacterium (SK1) degraded 75% PCP at concentration of 500 mg/kg within 168 h at pH 7.5 and 35°C temperature, which is superior to previous findings. Khessairi et al. (2014) found that Janibacter sp. FAS23 degraded PCP at a concentration of up to 300 mg/L supplemented with 1% of glucose in neutral pH and temperature of 30°C. With recent study indicating that there are still many unknown bacteria in nature that may have more survival and degradation ability for PCP, it is very important to screen the potential bacterial strain from PCP-contaminated soil and water. From this investigation, we report for the first time that Vibrio sp. (SK1) are able to metabolize PCP as a carbon/energy source. This strain tolerated up to 500 mg/L (1.8 mM) of PCP and also degraded 75% of PCP into the culture medium. Commonly, polychlorinated phenols(3 to 5 chlorine containing compounds) converted to chlorohydroxyquinones will probably be intermediate (hydroxyquinone pathway); subsequent reaction progressively removes chlorine, whereas the lower chlorinated phenols are attacked by monooxygenase and product may be chlorocatechol intermediates (chlorocatechol pathway), which are subjected to ring cleavage before dechlorination (Solyanikova and Golovleva, 2004). The results of this study clearly demonstrate that Vibrio sp. strain is able to use PCP by increasing its biomass and also chloride ion concentration that increased significantly into the medium. The liberation of chloride ion in the medium is considered to be the mineralization of chlorinated compounds probably by the hydroxyquinone pathway. Present results supported by the previous finding of other researchers indicated that chlorinated phenols are mineralized to chlorine-free end products (Mohn and Kennedy, 1992; Karn et al., 2010a).
PCP removal in soil by using Vibrio sp. (SK1)
Physiochemical characteristics of soil was also done pH 6.4 ± 0.5; carbon 21.24% ± 2%; hydrogen 2.1 ± 0.7; nitrogen 2.9 ± 0.5; sulfur 0.5 ± 0.01; phosphorus 1.7 ± 0.4; oxygen 32.0% ± 2.1%. PCP removal in soil was analyzed by inoculating the Vibrio sp. (SK1) into soil. The initial PCP concentration in the soil was very less and therefore 100 mg/L (0.37 mM) of PCP was artificially increased in the soil, to observe the efficacy of this strain. The survival of this strain was checked by CFU. The growth of the bacterial strain increased with time, in both soil supplemented with PCP and soil alone. The growth of Vibrio sp. was higher in soil alone compared to soil amended with PCP (Fig. 3). Vibrio sp. (SK1) was able to remove up to 72% PCP from the soil, whereas in control, the PCP concentration remains same. This result suggested that the isolated bacterial strain (SK1) has the potential to grow well, survive, and remove PCP from soil in the short span of time. Therefore, this strain is found be very effective for the removal of PCP from the soil. Previously, some research also has been carried out, like Dai and Copley (2004) used microcosm to remove PCP from the real soil, but observed that degradation capacity was very less, about 10% in 28 days of incubation. Alber et al. (2000) used Sphingomonas sp. UG30 for PCP mineralization and degradation at three different PCP concentrations (100, 225, and 500 mg/kg of PCP) for 22 days in statically incubated soil. For the first two concentrations, 100 and 225, they found a degradation of 25% and 65%, respectively. At 500 mg/kg of PCP soil, there was no degradation. Colores and Schmidt (2005) used Sphingomonas chlorophenolica RA2 in a microcosm soil contaminated with the following concentrations: 0, 10, 50, 100, or 300 ppm of PCP. S. chlorophenolica RA2 degraded only 10% in the soil contaminated with 10 ppm PCP, reaching 30% after 3 weeks. No degradation was observed at any other concentrations. By observing the previous finding by various researchers, we found that SK1 has much better efficacy for PCP removal and for survival in PCP-contaminated soil. Early in situ remediation of PCP studies was carried out in compost, manure, and soil by Laine and Jorgensen (1997). Gautam et al. (2003) applied Pseudomonas sp. IST103 two sets of soil microcosms containing 20% and 40% moisture, each having the following PCP concentrations: 0, 10, 100, 500, and 1,000 mg/L. Pseudomonas sp. showed significant use of PCP, about 80%, with higher cell growth after 45 days, the highest being when PCP was applied up to levels of 100 mg/L, and at a concentration of 20% and 40% moisture, about 70% of PCP was used. At 500 and 1,000 mg/L PCP showed inhibitory effects on the growth and PCP utilization. Pu and Cutright (2007) evaluated PCP biodegradation in soils from Columbia. The soils were incubated for 56 days with Flavobacterium sp. (ATCC 21918) as well as the mixed culture at concentrations of 539 mg/kg. After 56 days in Columbia soil, the biodegradation efficiencies were 12% and 25% for both Flavobacterium sp. and mixed culture. A strain of Pseudomonas fluorescens was found in South Africa by Lin et al. (2008) and was found to be able to degrade 200 μM PCP after 4 days. Vitkova et al. (2011) considered the degradation capacity of bacterial strain Pseudomonas testosterone CCM 7530 by means of bioaugmentation and biostimulation in PCP-contaminated soil. The biodegradation of PCP in soil was carried out under laboratory conditions at concentrations of 10 and 100 mg/kg of PCP. For each experiment, three sets of soil samples were used and analyzed after 7, 14, and 21 days; the soil samples with concentrations of 10 mg/kg PCP revealed higher degradation in comparison to soils with concentrations of 100 mg/kg PCP, especially chernozem where the highest degradation was 78% in nonamended soil and 55% in lignite-amended soil. Biodegradation of PCP in bioaugmented soils was evaluated after 21 days. By comparing with other strains, we found that this strain (SK1) is much more efficient for PCP removal in the short period of time by showing good survival into the natural condition (soil) because for many strains, this becomes a major limitation for the in situ remediation.
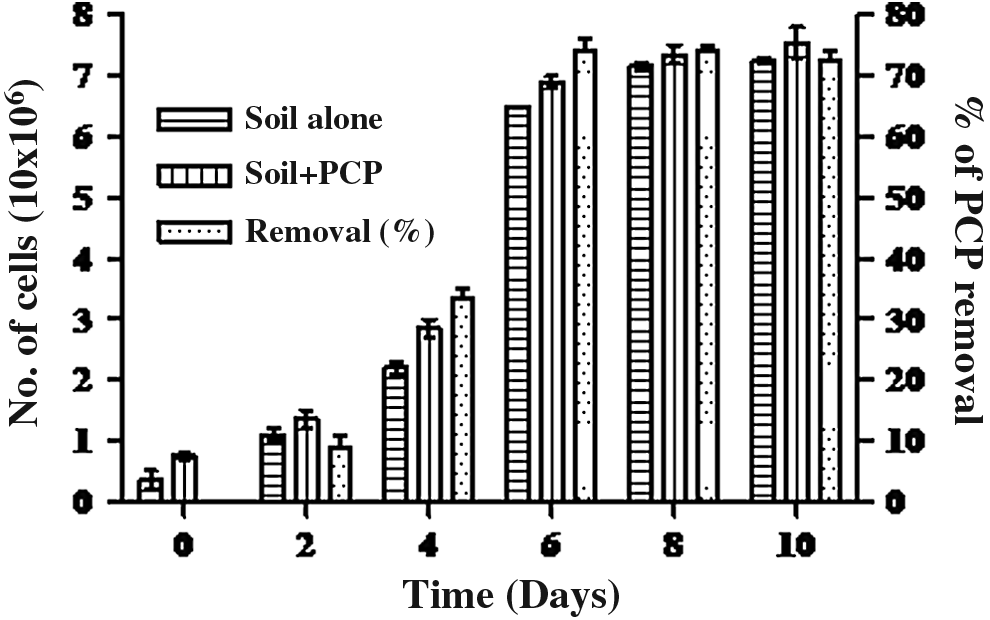
Survival, growth, and removal capability of Vibrio sp. (SK1) in soil alone and soil with PCP. Values are represented in mean ± SD (n = 3). Error bars represent standard deviation of three independent experiments.
PCP stabilization/removal by using biochar
An additional same soil treated with Vibrio sp. was supplemented with biochar 1%, 2.5%, and 5% produced from Pistia stratiotes leaves. The mineralogical characteristics of biochar was also analyzed and are as follows: carbon 73.54% ± 2%; hydrogen 3.1 ± 0.8; nitrogen 0.7 ± 0.02; sulfur 0.1 ± 0.03; and phosphorus 0.5 ± 0.04. In this experiment, biochar plays an important role to fix the PCP in the soil, probably by sorption. Vibrio sp. removed PCP up to 72%. Further PCP was left (18% PCP) in the soil that was basically fixed/sorption by biochar in 1 week period (Fig. 4). Soil treated with 5% biochar has no PCP left in the soil. Previously researcher observed that biochar used for sorption of organic contaminants to carbonaceous materials to treat contaminated soil/sediment, carbonaceous material or biochar easy to prepare and also stabilizing the pollutant high level. Lou et al. (2012) suggested that biochar is better for controlling organic pollutants present in sediment and observed sorption of PCP from sediments supplemented with rice straw biochar by adding 2% black carbon to the sediments. The sorption capacity of PCP onto black carbon sediment increased with increasing amounts of black carbon. The PCP adsorbed onto the black carbon supplemented sediments by 10%. Black carbon sorption varied with the amount and source of the black carbon at room temperature (25°C ± 1°C). It was reported that the surface area and sorptive ability of sediment supplemented with char was higher compared with sediment alone. Xu et al. (2012) used column leaching experiments by the leaching ability of PCP in soil amended with bamboo biochar (0%, 1%, 2%, and 5% w/w). After leaching (methanol and distilled water), he found that residual bioavailability of PCP by the addition of 5% bamboo biochar decreased the cumulative leach-loss content of PCP by 42% and also reduced PCP concentrations by 56 and 65% in methanol and distilled water. Therefore, the use of carbon-containing material for soil remediation is an important component to decrease the bioavailability of organic pollutants.
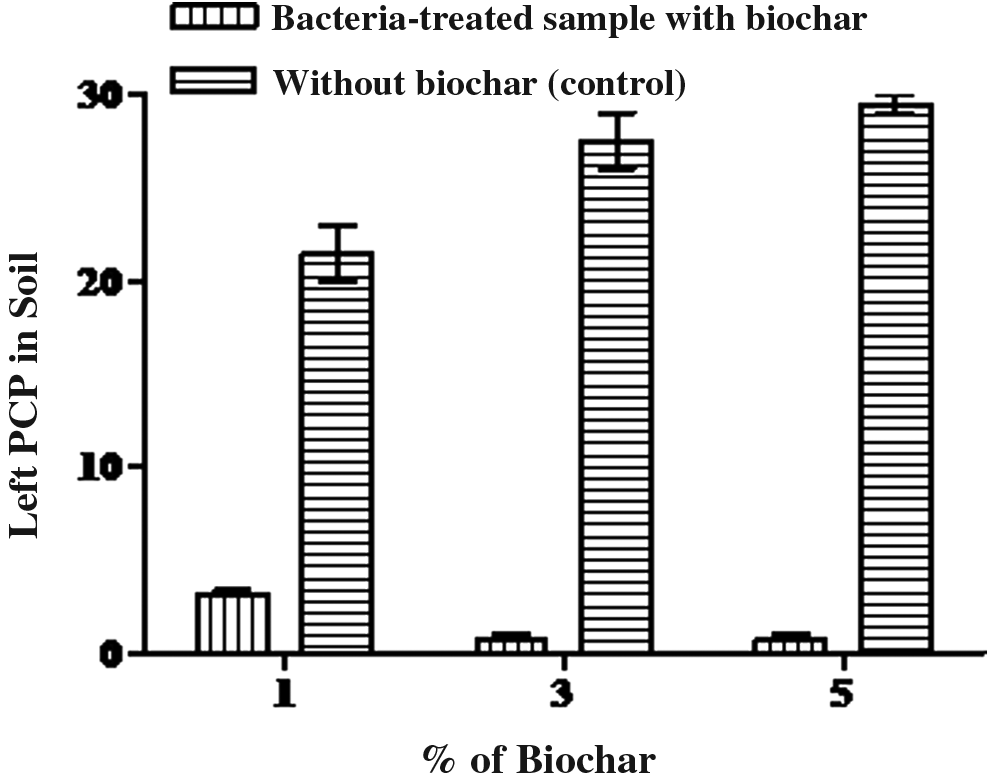
Observation of application of biochar to the bacteria-treated soil and ability to stabilize or immobilize the PCP. Values are represented in mean ± SD (n = 3). Error bars represent standard deviation of three independent experiments.
Different kinds of adsorbents were used in context to remove PCP from soil water like activated carbon, peat clay, and biological sorbents. Jianlong et al. (2000) observed that using activated sludge biomass to determine the adsorption strength of PCP in aqueous solution indicated the initial biomass concentration affects the adsorption capacity maximum in the range of about 5 g/L. Recently, Devi and Saroha (2014) prepared and used zero-valent iron magnetic biochar composites (ZVI-MBC) from the paper mill sludge biochar ZVI-MBC, combining the advantages of biochar and ZVI particles for the simultaneous adsorption and dechlorination of PCP in the effluent, and the complete removal of PCP was obtained from the effluents. Currently, biochar has been focused in research and commercial interest because of its role in sequestering carbon and increasing nutrient in the soils. Earlier, biochar used to remove various other organic or recalcitrant compounds by different researcher to remediate the soil like Cao et al. (2011) used dairy manure biochar and incubated with contaminated soils at rates of 2.5%, and 5.0% by weight for 210 days. After 210 days, soils treated with the 5.0% biochar showed more than 66% reduction of atrazine concentration; this showed biochar as a unique amendment for immobilization of organic contaminants in contaminated soils. Chen et al. (2012b) also used biochar prepared from orange peel, pine needle, and sugarcane bagasse; two different temperatures were used to prepare: low-temperature biochars at 300°C and high-temperature biochars at 700°C. Adsorption of 1-naphthol to high-temperature biochars was more. Low-temperature biochars had shown exceptional sorption behavior where partition and adsorption were concurrently promoted by a cosolute and elevated temperature. Chen et al. (2012a) used pine wood biochars to remove naphthalene; naphthalene exhibited a fast rate of sorption. Sun et al. (2013) observed sorption of phenanthrene by plant-residue derivative biochar, animal waste-derived biochar and their corresponding deashed biochar. Immobilization and stabilization processes observed in soil are of great environmental importance because of considerable reduction in the bioavailability of toxic compounds.
Kinetic modeling and parameter estimation
The performance and control of biological processes can be elaborately studied by using modeling. Execution of model-centered approaches for process development stay incomplete at industrial level, primarily since the process benefit/product cost ratio is not clear till now smart for such improvements in process procedure. Biochemical processes are highly complex and frequently unwell implicit on a mechanistic origin mostly in concern with the microorganism's growth mechanisms. In our work for procedure analysis, mathematical models, based on first principles, are used to represent the mechanistic preceding information about the process in study. Frequently, mathematical process models are deliberated to represent the a priori information. The models in biochemical engineering and in particular with respect to process optimization and control, consideration of relationships, which explains the simple features of the real process that, are greatest significant to process performance. In biotechnology, mathematical models are built using some mechanistic concepts, which are embedded into mass balances, that is, a relatively simple set of differential equations (e.g., Sonnleitner and Käppeli, 1986; Lübbert and Simutis, 1994; Thomson and Kramer, 1994).
In our study, a parameter estimation method was used for mathematical models along with kinetic equation to investigate the most important phenomena influencing the biochar production and biodegradation process. Experimental data were used to numerically estimate the model parameters and the quality of fit was quantified. Results obtained reveal that the Contois and Moser models are better fitting with these data when compared with the Monod, Tessier, and logistic growth curve. The Monod kinetic equation describes specific growth rate as a function of substrate concentration, R2 value obtained is 0.567, given in Table 2. The R2 values obtained with logistic growth curve, Contois-Ksis, and Moser-Ksis are 0.879, 0.985, and 0.921, also given in Table 2. Hence, Contois model is better fitting with these data since with the feature of the Contois growth model, the cell-mass growth rate depends on the concentrations of both substrate and cell mass, with growth being inhibited at high concentrations of the cell mass. Here, biodegradation is going on while cell mass is growing, whereas Moser model showing a better fitting since introducing exponent, called “the shape factor,” allows modulating the shape of the plot at the cost of introducing an additional parameter. Logistic growth curve giving R2 value of 0.879 can be interpreted as growth is increasing exponentially, not giving any information about substrate. Thus, both Contois and Moser kinetics are considered to be partially applicable models to explain kinetics behavior of Vibrio sp. for PCP biodegradation.
Isolate identification and their phylogeny
The 16S rRNA product, after PCR amplification, was purified and then sequenced using Applied Biosystem automatic sequencer. Sequence was checked in the GeneBank using BLASTN and RDP-II. Pairwise alignment revealed that 16s rRNA gene of the bacterial isolate (SK1) had 99% identity with Vibrio sp. The 16S rRNA gene sequences determined in this study have been deposited in GenBank of NCBI database under the accession number (KT626450).
Conclusion
This article introduced a new bacterial sp. Vibrio sp. (SK1) having the ability to tolerate up to 500 mg/L of PCP; more importantly, this microorganism has a strong ability to survive in the soil, grows well, and degrades PCP from the contaminated soil (100 mg/kg) within 1 week. Biochar played a better role in stabilizing the PCP in the soil by sorption concurrently; biochar addition also increases the health of soil. This is first principle models in conjunction with kinetic models are built, further nonlinear graphical regression method with the logistic growth curve, Monod, Contois, Moser and the Tessier models, the kinetic parameters were predicted. The logistic model has shown as an efficient model for kinetic reciting for PCP degradation and PCP leftover. Future studies will be completed using aged soil from contaminated field sites to determine whether desorption limits PCP removal during treatment with SK1 and also will be tested for bioaugmentation on unsterilized soil to reveal whether the isolate is more effective than native PCP degraders under similar laboratory conditions.
Footnotes
Acknowledgments
The authors are thankful to Department of Biotechnology Engineering, Ambala, College of Engineering and Applied Research, Ambala, and National Institute of Technology (NIT), Raipur (CG), India,for providing facility for this work.
Author Disclosure Statement
No competing financial interests exist.