
Abstract
Abstract
A halotolerant chromate [Cr(VI)]-resistant bacterium, CrRPSD40, isolated from marine sediments of Paradip port, Odisha, India, was identified and used in Cr(VI) reduction study. Based on biochemical characterization and 16S rRNA gene sequencing, the bacterium CrRPSD40 was identified as Brevibacillus laterosporus, with 97.7% homology to B. laterosporus. The bacterium has a high Cr(VI) resistance (up to 2,100 mg/L) and a high salt tolerance (up to 11% w/v NaCl) capacity. Bacterial growth in the presence of Cr(VI) (100 and 500 mg/L) showed a relatively longer lag phase (0–12 h) and a shorter exponential phase (12–72 h) compared with growth without Cr(VI). Cr(VI) reduction by B. laterosporus was studied at various ranges of pH (4.0–10.0) and NaCl (1–11% w/v) concentration, at 35°C temperature, and at 130 rpm agitation speed. The optimum saline and pH condition for better growth and Cr(VI) reduction by CrRPSD40 were found to be 3% w/v NaCl and 8.0, respectively. Under the optimum condition, CrRPSD40 exhibited efficient Cr(VI) reduction as much as 92% and >50% of 100 mg/L Cr(VI) in just 120 and 60 h of treatment, respectively. Comparative changes in bacterial cell surface morphology due to interaction with Cr(VI) were evident from the field emission scanning electron microscope-energy dispersive X-ray spectroscopy (FESEM-EDX) and Fourier transform infrared (FTIR) analyses. The present investigation described the significant reduction potential of the native marine B. laterosporus (CrRPSD40) under the wide range of saline and pH conditions for its successful implementation in Cr(VI) detoxification of a saline metal-polluted environment.
Introduction
T
Recently, the use of chromium-resistant bacteria as bio-remediators for the transformation of Cr(VI) to Cr(III) has emerged as a safe and cost-effective technology when contrasted against traditional physicochemical methods (Megharaj et al., 2003; Pal and Paul, 2004; Chandra and Singh, 2014). However, the bioremediation of Cr(VI) from the saline environments such as industrial effluents (contains 1–10% w/v NaCl) is a major challenge since these effluents cannot be treated by nonsaline microbes due to the hindrance of their growth (Subramanian et al., 2012; Chandra and Singh, 2014). Hence, halophilic/halotolerant microbes from marine environments exhibiting salt as well as Cr(VI) tolerance are more suitable for Cr(VI) reduction in the saline environment (Subramanian et al., 2012; Chandra and Singh, 2014). Metal-resistant marine bacteria are potential agents for metal removal from the saline environment, as they are adapted to the most adverse marine environments and have variable temperature, pH, salinity, conductance, etc. (Subramanian et al., 2012; Dash et al., 2013). However, the availability of halotolerant and efficient Cr(VI)-reducing bacteria is a problem since very few halophilic/halotolerant bacteria such as Bacillus subtilis SHB13 (Swapna et al., 2016), Halomonas sp. CSB5 (Chandra and Singh, 2014), Planococcus maritimus VITP21 (Subramanian et al., 2012), Vigribacillus sp. (Mishra et al., 2012), and Exiguobacterium indicum MW1 (Mohapatra et al., 2017) were reported for bioreduction of Cr(VI) in the saline environment.
Further, bacterial Cr(VI) reduction depends on factors such as pH, salinity, and Cr(VI) concentrations. Bacterial growth and its chromium reductase activity are affected by pH changes due to the alternation of the ionic form of the enzyme's active site (Kathiravan et al., 2010). In acidic pH, the binding sites of the cell wall are blocked with hydrogen ions and hinder the interaction of cations with surface functional groups by exerting repulsive forces (Aryal and Liakopoulou-Kyriakides, 2015). Maximum Cr(VI) reduction by most of the saline bacteria such as B. subtilis SHB13, Vigribacillus sp., Nesterenkonia sp. MF2, and E. indicum MW1 was achieved at pH ∼7.0 to 8.0 (Amoozegar et al., 2007; Mishra et al., 2012; Swapna et al., 2016; Mohapatra et al., 2017). Mangaiyarkarsai et al. (2011) and Subramanian et al. (2012) reported that a gradual increase of Cr(VI) reduction by alkaliphilic B. subtilis and P. maritimus VITP21 was observed from pH 6 to 9, confirming that the alkaline condition favors the Cr(VI) reduction compared with a neutral or acidic condition. Cr(VI) reduction by halotolerant Vigribacillus sp. increased marginally with an increase in NaCl concentration as much as 6% w/v and thereafter decreased gradually by a further increase in NaCl concentration (Mishra et al., 2012). Substantial removal of high Cr(VI) concentration by halotolerant P. maritimus was obtained at 4% NaCl (Subramanian et al., 2012). Therefore, isolation of salt and chromate-resistant bacteria and evaluation of their Cr(VI) reduction potential are the essential requirements for bio-reduction-based bioremediation of toxic chromate from the saline waste.
Paradip port harbor, situated on the Bay of Bengal along the Odisha coastal line of Eastern India, frequently comes into contact with various heavy metals, especially chromium, during the transportation of metal ore, including chromite (Raj et al., 2013; Palleyi et al., 2015). This leads to the deposition of metals in the port sediment by settling down through the water column. Therefore, there is a possibility of native marine sediment bacteria developing a resistance toward various heavy metals. Among all the isolated chromate-resistant native sediment bacteria, CrRPSD40 bacterium was found to show high Cr(VI) resistance as well as tolerance to six other toxic metals. Hence, the present study was involved in the molecular identification of the Cr(VI)-resistant CrRPSD40 bacterium and the evaluation of its Cr(VI) reduction potentiality under various saline and pH conditions. Further, the interaction of the bacterium cell with the Cr(VI) metal ion and specific changes in cell surface morphology during reduction was studied by FESEM-EDX and Fourier transform infrared (FTIR) analyses.
Materials and Methods
Materials
Culture media and reagents used throughout the study were obtained from Himedia laboratory. All chemicals used for preparations of metal stock solutions and chromium analysis are of analytical grade purchased from Merck India Pvt. Ltd. Cr(VI) standard stock solution was prepared by dissolving potassium dichromate in ultrapure Millipore water. All the glasswares used in this experiment were metal free via washing with nitric acid followed by triplicate rinsing with distilled water.
Characterization and identification of CrRPSD40 bacterium
Marine bacterium CrRPSD40 was isolated from the sediment of Paradip port, Odisha, and it showed high tolerance toward Cr(VI), including six other heavy metals (Mohapatra et al., 2016). In a previous study, the bacterium CrRPSD40 was phenotypically identified as Brevibacillus sp. by means of morphological and biochemical characterization as per Bergy's manual of determinative bacteriology (Mohapatra et al., 2016). In the present study, further genotypic identification of CrRPSD40 was done by 16S rRNA gene sequencing and the phylogenetic method. For sequencing, bacterial DNA was first isolated from the strain and its quality was evaluated on 1.2% Agarose gel. A fragment of the 16S rRNA gene was amplified by PCR, and the amplicon was purified to remove contaminants. Forward and reverse DNA sequencing reaction of PCR amplicon was carried out with 8F and 1492R primers by using BDT v3.1 Cycle sequencing kit on ABI 3730xl Genetic Analyzer. All the sequencing procedures described earlier were carried out at Xcelris Genomics Labs Ltd. (Ahmedabad, India; www.xcelrislabs.com). Phylogenetic analysis was done for the 16S rRNA gene sequences by retrieving the similar matches of the selected Cr(VI)-resistant strain from EzTaxon, and multiple sequence alignment was performed by using Clustal_X program (Thompson et al., 1997). The evolutionary relatedness of all the strains was inferred by using the Neighbor-Joining method; distance was calculated by using the p-distance method, followed by a bootstrap test in 1,000 replicates for each cluster; and the Phylogenetic tree was drawn in MEGA5.0 program (Tamura et al., 2011).
Growth of CrRPSD40 under varying NaCl and Cr(VI) concentrations
Effect of salt (NaCl) concentration on the growth of Cr(VI)-resistant CrRPSD40 bacterium was investigated. The bacteria were grown in nutrient broth containing 1%, 3%, 5%, 7%, 9%, and 11% w/v NaCl and their growth was measured till 96 h at postinoculation intervals of 8 h. Similarly, the change in bacterial growth pattern under Cr(VI) stress was evaluated in the presence and absence of Cr(VI). For this study, fresh CrRPSD40 bacterial culture was inoculated to nutrient broth that was supplemented with 100 and 500 mg/L Cr(VI) at 3% w/v NaCl. Nutrient broth without Cr(VI) that served as a control was also taken in a similar condition, and the bacterial growth was measured in a specific interval of time starting from 0 to 120 h of incubation. For both the experiments, bacterial cell growth was measured by taking optical density at 600 nm by using a Uv-Vis spectrophotometer (VARIAN Cary 50 Bio). The growth experiments described earlier were carried out (in duplicate) in a neutral pH (∼7.0) at 35°C temperature, 130 rpm agitation speed, and the mean data with the standard errors were recorded.
pH optimization for enhanced Cr(VI) reduction
The Cr(VI) reduction process is affected by pH due to the influence of H+ ion on bacterial cell surface functional groups (Aryal and Liakopoulou-Kyriakides, 2015). To evaluate the optimum pH for potential Cr(VI) reduction by the strain CrRPSD40, 100 mg/L of Cr(VI) was used in a wide range of pH from 4.0 to 10.0 (maintained in nutrient broth). The experiment was carried out till 120 h by using 3% w/v NaCl, 35°C temperature, and 130 rpm agitation speed. Cr(VI) concentration in the culture was measured at a regular interval of time by drawing the cell-free supernatant sample from each pH condition. This pH dependence reduction experiment was carried out in duplicate, and data were reported as a mean value with standard errors.
Cr(VI) reduction under optimum pH and salinity by CrRPSD40
Optimum values of pH and salinity are the most favorable points at which better Cr(VI) reduction has been achieved. The Cr(VI) reduction potential of the bacterial strain CrRPSD40 was evaluated at optimum pH 8.0 and salinity (3% w/v NaCl) conditions in nutrient broth medium containing 100 mg/L Cr(VI). Reduction of Cr(VI) was carried out till 120 h at 35°C and 130 rpm inside an orbital shaking incubator (REMI-24BL). Concentrations of Cr(VI) and Cr(III) were analyzed at each 12 h interval starting from 0 h of incubation. Bacterial growth was also measured simultaneously at the same interval of time by taking optical density at 600 nm. A reduction experiment was carried out in duplicate, and data were reported as a mean value with standard errors.
Analysis of chromium
Hexavalent chromium in the culture was estimated by the diphenyl carbazide method (Eaton et al., 1995). A fixed volume of culture supernatant was taken out by centrifuging the culture at 10,000 rpm for 15 min at 4°C by using a cooling centrifuge (REMI Compufuge-CPR24). Two milliliters of supernatant was mixed with 2 mL of 3 M H2SO4 and 1 mL of 1,5-diphenyl carbazide (0.25%). After 10 min of incubation at room temperature, the absorbance of the mixed solution was measured at 540 nm by a UV spectrophotometer against known standards. The total chromium of each sample was analyzed by an Atomic Absorption Spectrophotometer (SHIMADZU AA 7000), and the Cr(III) concentration was calculated by deducting the Cr(VI) concentration from the total chromium concentration of the respective sample (Dhal et al., 2010).
FESEM-EDX and FTIR analysis of reduced residues
During the Cr(VI) reduction process, the bacterial cell association with a reduced product and the assessment of the interaction between Cr(VI) and the cell surface were investigated by using a field emission scanning electron microscope-energy dispersive X-ray spectroscopy (FESEM-EDX). The culture of CrRPSD40 bacteria during the mid-exponential phase (72 h old) from both Cr(VI)-treated (100 mg/L) and -untreated (control) cells was taken and centrifuged at 10,000 rpm for 15 min at 4°C. Cell pellets were washed with phosphate-buffered saline (PBS, pH 7.2) three times to remove the extra culture media. Harvested cells were fixed with 3 mL of 2.5% glutaraldehyde in PBS buffer for 3 h followed by postfixation and washing thrice with PBS (Nithya et al., 2011). Then, the cells were dehydrated with 30%, 50%, 70%, 90%, and 100% ethanol consecutively and placed on the aluminum stubs (Mangaiyarkarasi et al., 2011). The specimens were mounted on the sample holder with carbon-conductive adhesive tapes and coated with gold before viewing by FESEM (Carl Zeiss, SUPRA GEMINI55) at 5,000× and 10,000× magnifications with simultaneous EDX analysis (Dhal et al., 2010; Nithya et al., 2011). For FTIR spectroscopy analysis, Cr(VI)-treated (100 mg/L) and -untreated (as control) bacterial cells were pelleted out from respective cultures by centrifugation, as described earlier for the FESEM study. Then, the bacterial cells with a reduced product were dried in an oven at 60°C overnight and kept in a desiccator before FTIR analysis. The FTIR spectra of bacterial cell residues with the KBr (1:100) phase were recorded under analysis conditions of 45 scans at a resolution of 4 cm−1 that was measured between ranges of 400 and 4,000 cm−1 by using an FTIR spectrophotometer (SHIMADZU IRPrestige-21) (Pei et al., 2009; Dhal et al., 2010; Chandra and Singh, 2014).
Results and Discussion
Characterization and identification of isolated CrRPSD40 bacterium
As reported earlier (Mohapatra et al., 2016), the bacterial isolate CrRPSD40 selected in the present study exhibited the highest minimum inhibitory concentration value (2,100 mg/L) for Cr(VI) and also showed resistance against six other toxic metals, including Zn, As+3, Cu, Cd+2, Pb+2, and Ni. Bacteria that were isolated from metal-polluted toxic environments became metal resistant for their adaptation, and survival is a natural phenomenon (Rani et al., 2010). As Paradip port harbor is frequently contaminated with different metals during ore-handling activities, the bacteria grown in this environment can reflect the resistance characteristics of multiple metals. As mentioned in our previous report, a detailed morphological and biochemical characterization of CrRPSD40 was studied and it revealed that this strain was a Gram-negative, rod-shaped, and motile bacterium showing TSI alkaline, nonfermentative, and oxidative reactions. CrRPSD40 did not utilize Indole; it also showed negative results for the MR-VP test and positive results against the hydrolysis of enzymes such as casein hydrolase, gelatine hydrolase, alkaline phosphatase, oxidase, and catalase (Mohapatra et al., 2016) (data not shown).
In the present study, the bacterium CrRPSD40 was subjected to 16S rRNA gene sequencing and phylogenetic analysis revealed that CrRPSD40 belonged to the Brevibacillus cluster, with the closest relative being Brevibacillus laterosporus (Fig. 1). The sequence homology of the test organism to B. laterosporus CrRPSD40 was 97.7%, and its consensus sequence was submitted to Gene Bank with an association number of KX905162. Bacillus brevis was first described by Migula in the year 1900 and it was further reclassified as a novel genus Brevibacillus along with nine other different species in this genus (Baek et al., 2006). According to Ruiu (2013), Brevibacillus genus is a new genus within the Brevibacillus brevis cluster. As described in various reports, Brevibacilus sp. is a Gram-positive, rod-shaped, spore-forming, and motile bacterium. In this study, phenotypic and genotypic characteristics of the identified bacterium CrRPSD40 were supported by the findings of Mallick and Mukherjee (2015), Arya and Sharma (2014), Song et al. (2011), and Baek et al. (2006).

Evolutionary relationship and phylogenetic analysis of isolated marine CrRPSD40 bacterium.
Growth of CrRPSD40 under different NaCl and Cr(VI) concentrations
Since the CrRPSD40 is a saline bacterium, it grows well at an elevated NaCl concentration. It has been suggested that bacterial Cr(VI) reduction is directly related to its growth in the presence of Cr(VI) and NaCl (Chandra and Singh, 2014). Thus, the influence of Cr(VI) and NaCl concentration on Cr(VI) reduction was evaluated. From the NaCl-dependent bacterial growth study, the strain CrRPSD40 showed optimum growth at 3% w/v NaCl and there was no significant variation in the growth of CrRPSD40 from 1% to 5% w/v NaCl, but after 5% w/v NaCl, the bacterial growth gradually decreased (Fig. 2). The decreasing order of bacterial growth in different NaCl concentrations (1–11% w/v) was found to be 3% > 5% > 1% > 7% > 9% > 11% (Fig. 2). An increased substantial reduction of higher Cr(VI) concentration by halotolerant Vigribacillus sp. and P. maritimus was achieved between 4% and 6% w/v NaCl (Mishra et al., 2012; Subramanian et al., 2012). Hence, the obtained result in this experiment regarding the role of NaCl concentration in bacterial growth and simultaneous Cr(VI) reduction was in accordance with the findings of the authors mentioned earlier.
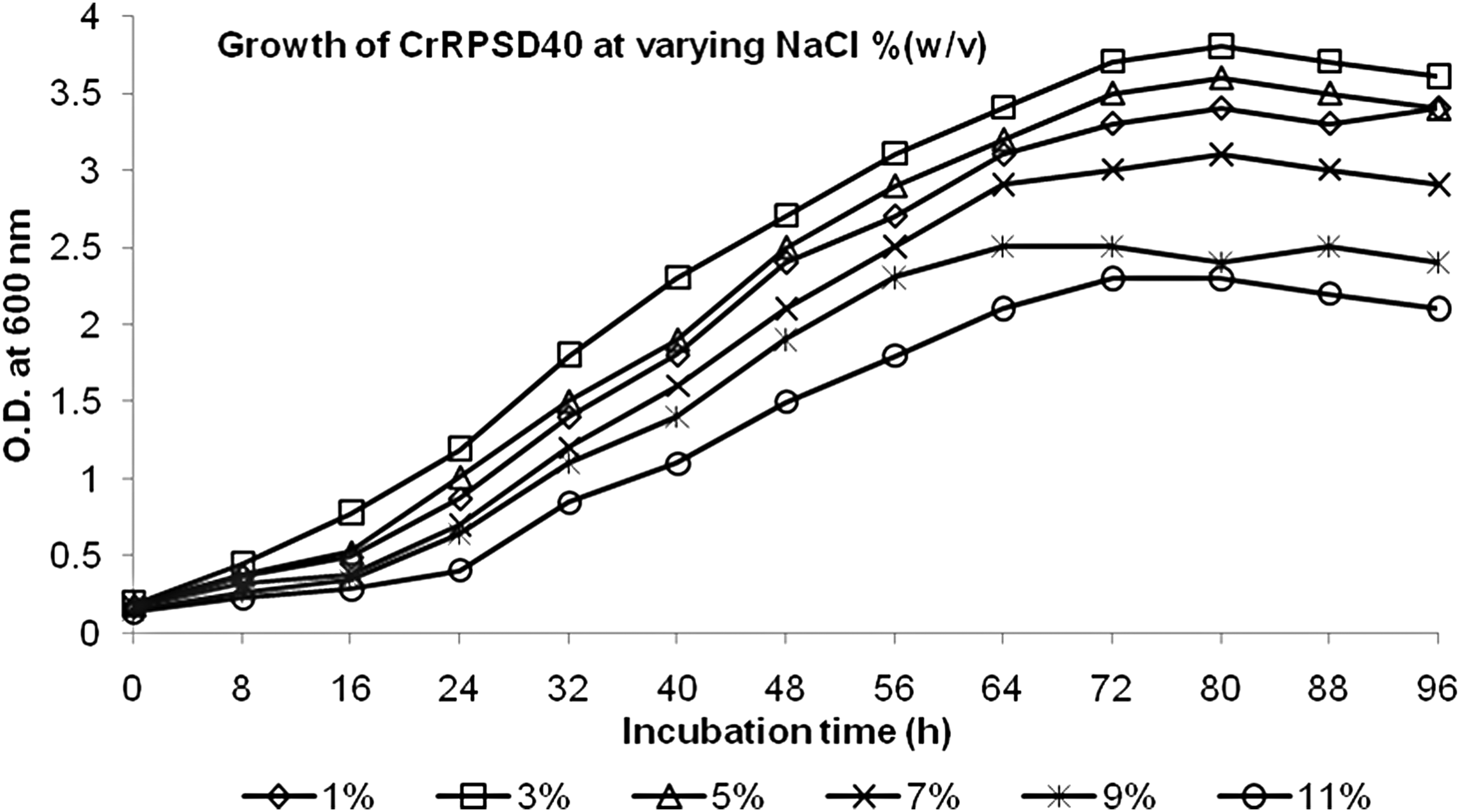
Growth of Brevibacillus laterosporus CrRPSD40 bacterium in nutrient broth supplemented with a varying concentration of NaCl.
The growth of CrRPSD40 in the presence of Cr(VI) (100 and 500 mg/L) and the absence of Cr(VI) (0 mg/L, control) was studied in nutrient broth medium supplied with 3% w/v NaCl. The growth pattern of the bacterium exposed to different Cr(VI) concentrations (100 and 500 mg/L) became slower and showed concentration-dependent growth as compared with the control (Fig. 3). In case of the control, the lag phase became shorter (0–8 h), followed by the exponential phase (8–84 h), and the stationary phase was reached after 84 h of incubation (Fig. 3). However, growth with 100 and 500 mg/L Cr(VI) showed a relatively longer lag phase (0–12 h) and a shorter exponential phase (12–72 h) as compared with the control, whereas the stationary phase was achieved after 72 h of incubation (Fig. 3). Overall, bacterial growth with Cr(VI) was not as such affected by the 100 mg/L Cr(VI) due to the high resistance capacity of the strain but in the case of 500 mg/L Cr(VI), it was slightly affected. This observed growth property of CrRPSD40 was also well supported by the findings of Mishra et al. (2012), during Cr(VI) reduction by moderately halophilic Vigribacillus sp.
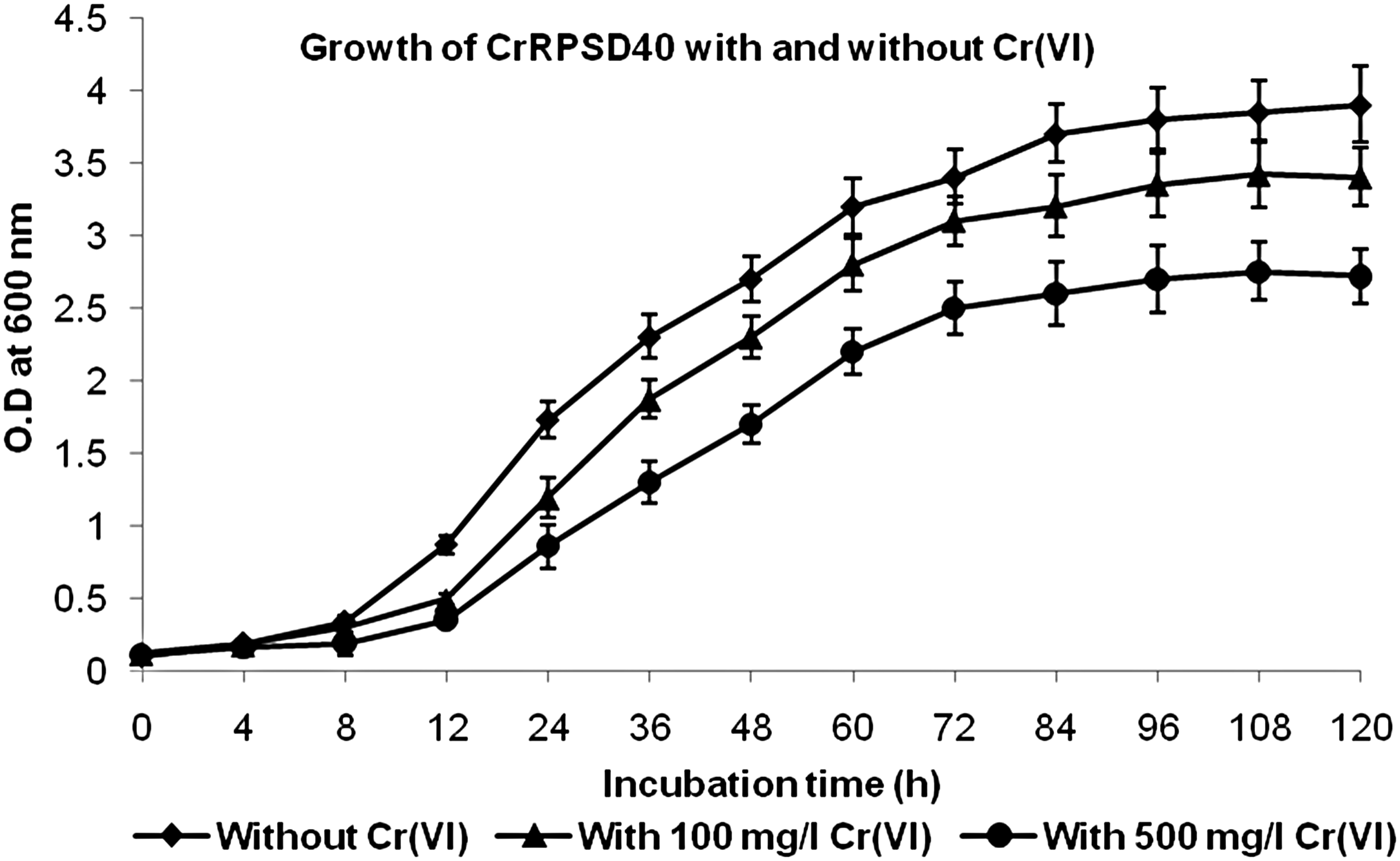
Growth of B. laterosporus CrRPSD40 bacterium without Cr(VI) and with Cr(VI) (100 and 500 mg/L) in nutrient broth medium containing 3% w/v NaCl.
Effect of pH on Cr(VI) reduction
Reduction of 100 mg/L initial Cr(VI) concentration by B. laterosporus CrRPSD40 under different pH range (4–10) is shown in Fig. 4. Higher reduction was observed between pH 7.0 (76%) and 8.0 (88%) whereas pH 8.0 was considered as optimum for Cr(VI) reduction. When the pH value became <6.0 and above pH 9.0, the reduction efficiency was drastically decreased. The decreasing order of Cr(VI) reduction with respect to pH after 120 h of treatment is as follows: pH 8.0 (88%) > pH 7.0 (76%) > pH 9.0 (64%) > pH 6.0 (51%) > pH 10.0 (49%) > pH 5.0 (19%) > pH 4.0 (7%) (Fig. 4). A similar type of observation was reported by Chandra and Singh (2014), that Cr(VI) reduction by halotolerant Halomonas sp. CSB5 was maximum at pH 8.0 and declined at pH 6.0, 9.0, and 10.0. A Cr(VI) reduction study in different pH ranges by using various alkaliphilic bacteria such as B. subtilis, B. subtilis SHB13, Vigribacillus sp, Nesterenkonia sp. MF, P. maritimus VITP21, etc. revealed that Cr(VI) reduction occurred only at alkaline condition (pH > 6.0) and a pH range from 6.0 to 9.0 mostly favors the Cr(VI) reduction by marine bacteria as compared with a neutral or acidic condition (Amoozegar et al., 2007; Mangaiyarkarsai et al., 2011; Mishra et al., 2012; Subramanian et al., 2012; Swapna et al., 2016). Less number of reports are available on Cr(VI) reduction in alkaline conditions (Swapna et al., 2016); hence, the present investigation on Cr(VI) reduction by halotolerant B. laterosporus CrRPSD40 was very significant and may be helpful for Cr(VI) reduction in saline waste.

Reduction of 100 mg/L Cr(VI) by the B. laterosporus CrRPSD40 bacterium in varying pH ranges (4.0–10.0) at 3% (w/v) NaCl.
Cr(VI) reduction under optimum pH and salinity(NaCl)
Cr(VI) reduction by the CrRPSD40 bacterium and its growth at optimum pH (8.0) and salinity (3% w/v NaCl) were measured simultaneously. Under optimum pH and saline condition, nearly 92% of 100 mg/L initial Cr(VI) was reduced to Cr(III) in 120 h of treatment (Fig. 5). A close observation of Fig. 5 shows that more than 50% reduction was achieved in just 60 h of treatment. As shown in Fig. 5, a correlation exists between bacterial growth and Cr(VI) reduction. The rate of Cr(VI) reduction was slower during the lag phase (0–12 h) and increased with a simultaneous increase in the bacterial cell growth during the log phase (up to 84 h). After 84 h of incubation, when bacterial growth reached its stationary phase, reduction rate was slow and became constant. A similar observation was reported by Xu et al. (2015) during Cr(VI) reduction by Bacillus anthracis Cr-4. Further, it was observed that Cr(VI) concentration gradually decreased whereas Cr(III) concentration increased with bacterial growth and residence time.
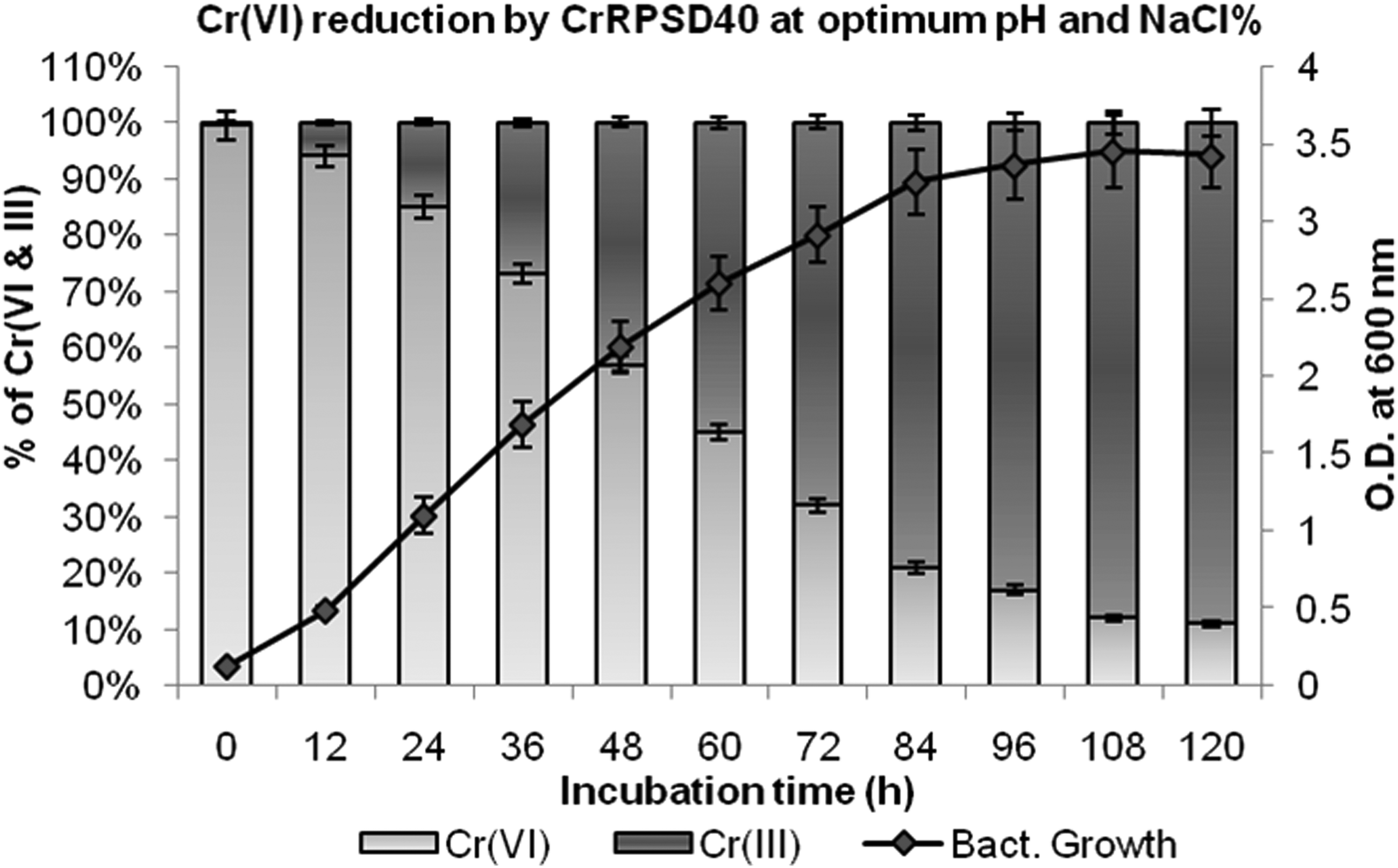
Reduction of 100 mg/L initial Cr(VI) to Cr(III) with respect to growth of the bacterium B. laterosporus CrRPSD40 at optimum pH (8.0) and NaCl (3% w/v).
When compared with previous reported works (Holail et al., 2011; Ibrahim et al., 2011; Mangaiyarkarasi et al., 2011) during Cr(VI) reduction using saline chromate-resistant bacteria such as Amphibacillus sp. KSUCr3, Bacillus sutilis, and B. laterosporus strain I4, the reduction efficiency of the present strain, CrRPSD40 was found to be better and was able to reduce a higher concentration of Cr(VI) with less time in a high saline condition.
FESEM-EDX and FTIR analysis
FESEM-EDX
The FESEM-EDX analysis of a 72 h-old culture of Brevibacillus sp. (CrRPSD40) grown in nutrient broth without and with 100 mg/L Cr(VI) is presented in Fig. 6. The SEM image shows cells of CrRPSD40 with spores and surrounding protective coat. The coat (Exosporium) is usually composed of protein with small amount of carbohydrate and lipid (Allievi and Mariano, 2011). The spore surface of the marine CrRPSD40 bacterium is porous and dominated by negatively charged particles that are mainly contributed by carboxylate and phosphate groups; these help in binding with heavy metals (He and Tebo, 1998). Here, a canoe-like structure (Fig. 6) was formed by B. laterosporus CrRPSD40 due to a possible interaction of spore surface charges with metal ions (He and Tebo, 1998; Azizbekyan et al., 2015). At both 5,000 × and 10,000 × magnification, the untreated bacterial cell surface became a little bit smooth, slender, nonporous, and nonfilamentous (Fig. 6a, b); whereas in the Cr(VI)-treated bacterial cell, distinct changes in cell surface such as rough, porous, bulged, and filamentous structures were observed (Fig. 6d, e). Similar observations were mentioned by Das et al. (2014) and Swapna et al. (2016) when Bacillus amyloliquefeciens and B. subtilis SHB13 were treated with Cr(VI). Morphological changes occurred, may be due to the formation of huge spores or surfactin having the chelating property of a toxic metal ion for protecting the cell from contact with Cr(VI) (Swapna et al., 2016). Further EDX spectral analysis of the bacterial residues both before and after Cr(VI) treatment showed that distinguishing chromium peaks were observed for bacterial cells that were grown with Cr(VI), unlike those that were nontreated (Fig. 6c, f). These Cr peaks occurred either due to the presence of precipitated reduced Cr(III) species around the cells or due to the complexion of Cr(III) species with cell surface functional elements (Dhal et al., 2010; Das et al., 2014). In addition to Cr peaks, some coexisting trace elements such as C, O, Na, P, Ca, and Al were also observed in both treated and untreated samples (Fig. 6c, f). Except for Al, all other peaks were previously reported elsewhere. The Al-peak obtained here may be due to a foil stub in the experimental system on which the samples were fixed.
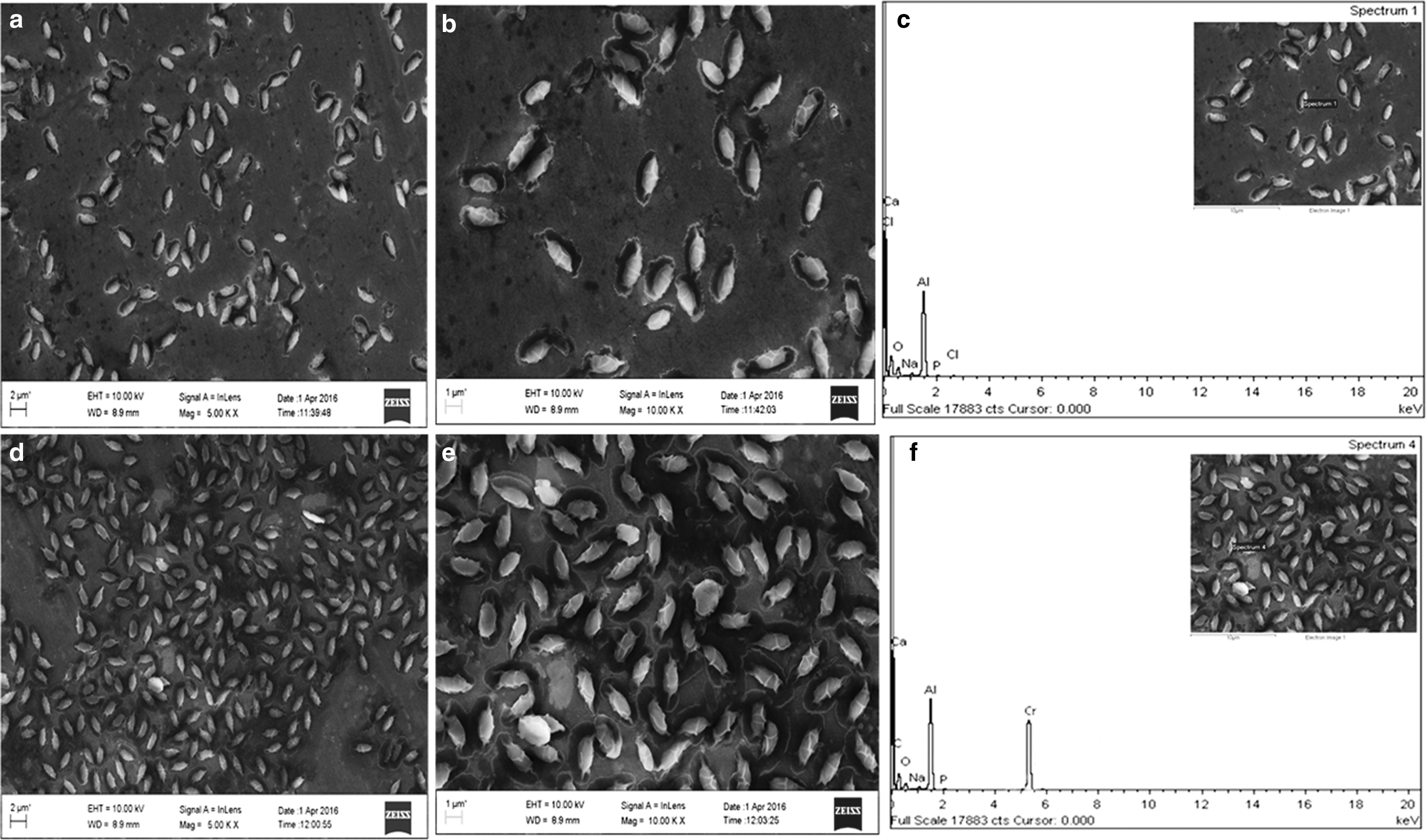
FESEM-EDX image of B. laterosporus CrRPSD40 bacterial cell growing without Cr(VI) at 5,000×
FTIR analysis
The analysis of FTIR spectra of Cr(VI)- treated and -untreated (control) B. laterosporus CrRPSD40 shows a possibility of metal ion interactions with the cell surface (Fig. 7). In both the cases, a broad absorption spectral peak that appeared at ∼3,250 to 3,350 cm−1 is attributed to the hydroxyl (–OH) and amines (–NH) stretching, most probably from polysaccharides and protein. The peak number 3,268 cm−1 in the control was slightly shifted to 3,295 cm−1 after treatment with Cr(VI), indicating a direct interaction of bacterial cell-surface hydroxyl and amines with the chromium metal ion (Mangaiyarkarasi et al., 2011; Chandra and Singha, 2014). The absorption bands that appeared at ∼3,175, ∼2,960, and ∼2,887 cm−1 represented the C—H stretching of the alkyl group and alkanes. The peaks obtained at ∼1,659 and ∼1,540 cm−1 correspond to Amide-I (Protein C═O stretching) and Amide-II (Protein N—H bending and C—N stretching) groups, respectively (Mangaiyarkarasi et al., 2011). In addition to these, there are also other small intensity peaks existing at ∼1,442, ∼1,236, ∼1,072, and ∼837 cm−1 representing the C—C— aromatic stretching, aromatic C—N stretching, PO2− asymmetric stretching, and C—O polar and C—H aromatic stretching, respectively (Pei et al., 2009; Mishra et al., 2012). In the case of the Cr(VI)-treated bacterial cell, there is a slight shift in the band frequency of ∼1,236 to ∼1,230 cm−1, indicating the involvement of phosphate linkage in chromium binding activity. Similarly, a slight shift of ∼1,659 to ∼1,653 cm−1 represents an intervention of C═O group of amide I bond. The peaks observed at 1,072 and 1,442 corresponding to C—C— and C═O stretching remained unchanged in both treated and untreated cells. In the present study, no such distinct significant difference in the appearance of various characteristic peak positions among treated and control cells was observed. Hence, from the FTIR peak analysis, it is confirmed that there is a certain role of hydroxyl, amine, phosphate, and amide-I cell-surface functional groups of B. laterosporus in interaction with Cr(VI) during the reduction process. Similar types of findings were reported by the authors Das et al. (2014) and Mangaiyarkarasi et al. (2011) during Cr(VI) reduction by Bacillus amyloliquefaciens and alkaliphilic B. subtilis, respectively.
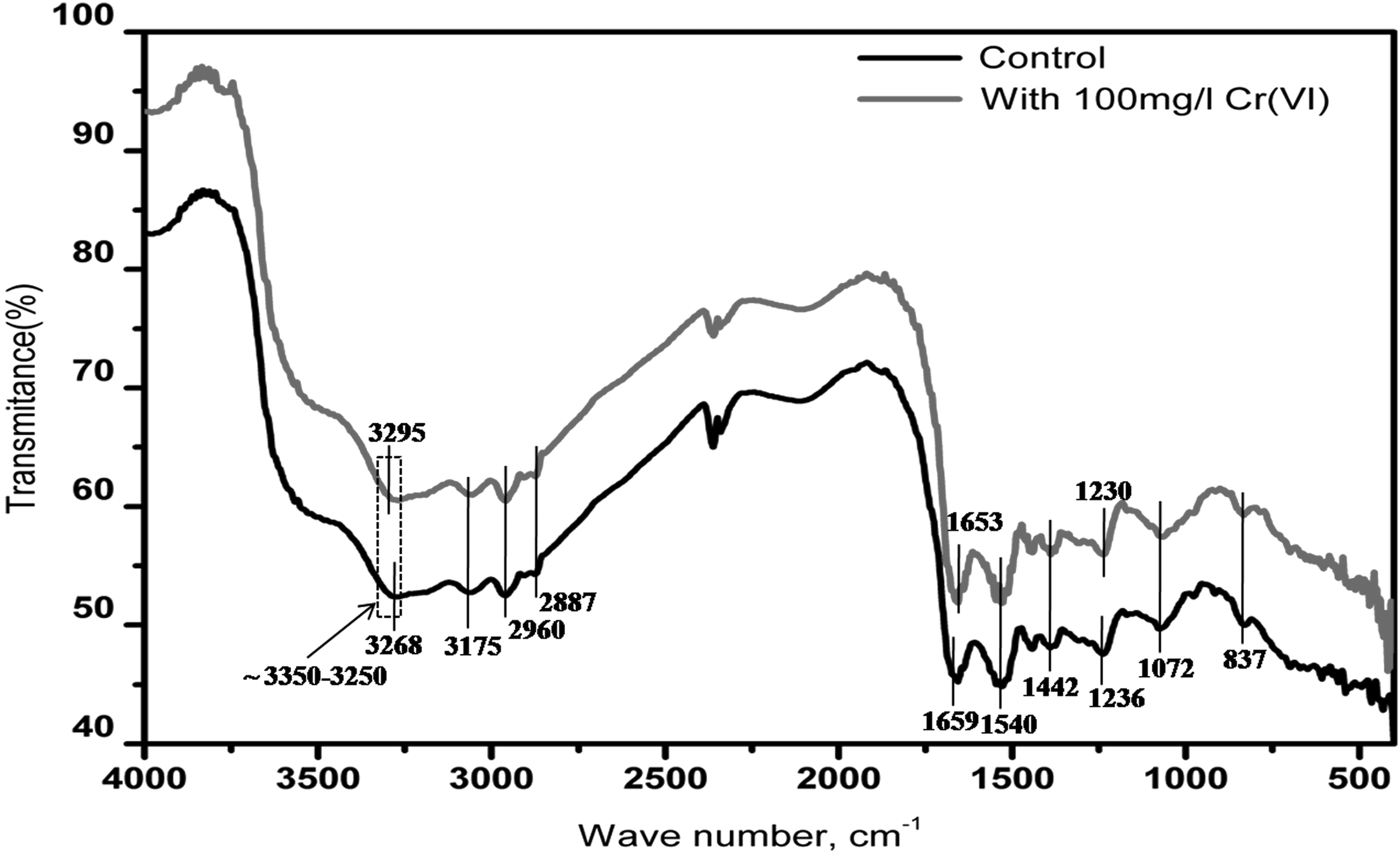
Fourier transform infrared (FTIR) spectral analysis of the B. laterosporus CrRPSD40 bacterial cell before treatment (control) and after treatment with Cr(VI).
Conclusion
A wide variety of metal-resistant and salt-tolerant bacteria from marine sources can be considered suitable candidates for Cr(VI) reduction, as they adapt quite readily to changing sea conditions. The isolated native marine sediment bacterium CrRPSD40 showed a 97.7% sequence homology to B. laterosporus during 16S rRNA gene sequence analysis. The B. laterosporus CrRPSD40 is a high Cr(VI)-resistant (2,000 mg/L) and salt (NaCl)-tolerant (11% w/v) bacterium, and its growth was not greatly affected by the presence of ≤500 mg/L of Cr(VI) and ≤5% w/v of NaCl. The isolated bacterial strain showed an effective Cr(VI) reduction ability by reducing 92% of 100 mg/L initial Cr(VI) to nontoxic Cr(III) in 120 h under optimal pH (8.0) and NaCl (3% w/v) at 35°C. This reduction rate increased with the simultaneous increase in the bacterial cell growth during the log phase between 12 and 84 h of treatment. The decrease of Cr(VI) concentration in the bacterial-treated culture was mainly due to the bacterial reduction rather than biosorption (confirmed by measuring Cr(VI) and Cr(III) concentration in the culture supernatant). FESEM-EDX and FTIR analysis revealed the morphological changes of the bacterial cell surface due to the attachment of Cr(VI) and Cr(III) particles during reduction processes Hence, B. laterosporus CrRPSD40 can be potentially exploited for developing a suitable bioremediation process in both the saline environment and industrial effluents containing multiple toxic metals.
Footnotes
Acknowledgments
The authors would like to thank Prof. B.K. Mishra, Director, CSIR-Institute of Minerals and Materials Technology, Bhubaneswar, India, for providing the necessary facility to carry out this work and also for his kind permission to publish their research findings.
Author Disclosure Statement
No competing financial interests exist.