
Abstract
Abstract
Effect of zinc oxide (ZnO) nanoparticles (NPs) size was evaluated in the context of bacterial bioluminescence, seed germination, gene mutation in bacteria, and algal growth. Different sensitivities and half maximal effective concentration (EC50s) were observed, showing orders of toxicity: bacterial bioluminescence (0.31–0.40 mg/L) > algal growth (1.08–2.90 mg/L) > seed germination (6.56–9.98 mg/L). Furthermore, size of NPs plays a critical role in toxicity. The mutation ratio decreases with increase in the size of NPs, with 2.4 for 5 nm and 1.1 for 80 nm, at 100 mg/L of ZnO NPs. Effect ZnO particle size differed according to tested assays. Under exposure conditions with large sizes, toxicity of ZnO NPs increased on seed germination, but decreased on algal growth and gene mutation. Toxicity results of algal growth and seed germination showed statistically significant differences (p < 0.05) between 5 and 80 nm ZnO NPs. In the case of bacterial bioluminescence, no significant differences were observed among each of the sizes tested. These findings show that a toxicity evaluation of NPs needs to consider the different effects with respect to particle sizes and tested organisms for an accurate assessment of toxicity.
Introduction
E
To accurately evaluate environmental impacts, it is important to choose appropriate bioassay organisms, end points, and methods for risk assessments. Bioassays are used for toxicological evaluation of contaminants and have gained widespread attention (Banks and Schultz, 2005). A range of test organisms (e.g., bacteria, algae, protozoa, plants, and fish) have been used to test the toxicity of chemicals. Research has shown that the toxicity of NPs can vary according to the bioassay (Soto et al., 2006; Mohanty et al., 2016). For example, widely used NPs, such as titanium dioxide (TiO2) and zinc oxide (ZnO), have been shown to have toxic effects, and can inhibit the growth of microalgae, crustaceans, and bacteria (Aruoja et al., 2009). On the other hand, contrasting results have been obtained with Vibrio fischeri, Daphnia magna, and Thamnocephalus platyurus (Heinlaan et al., 2008).
From an ecotoxicological perspective, several studies have shown that ZnO NPs are toxic to human colon epithelial Caco-2 cells (Cao et al., 2014). These NPs might have the ability to reach sites inaccessible by larger particles, resulting in accumulation, aggregation, or toxic effects. Different studies have reported a range of toxicity mechanisms (e.g., dissolved ions, interactions with algae, entrapment of algal cells into the NPs aggregates.) for a variety of NPs (Ji et al., 2011). However, there is no report to corroborate the impact of ZnO NPs having similar size and concentration on range of organism tested for environmental studies. Therefore, in this study, we examined the effect of ZnO NPs on bacterial bioluminescence, seed germination, algal growth, and gene mutation.
We choose the seed germination test, because of its many advantages, such as sensitivity, simplicity, and suitability for reactive chemicals and contaminated soil samples (Di Salvatore et al., 2008; El‐Temsah and Joner, 2012). Similarly, bioluminescence assays are time saving and cost effective, thus used as a reproducible and sensitive screening method for determining acute toxicity (Wang et al., 2002). Algae are important organisms in the aquatic ecosystem used as a model organism for examining toxicity of many NPs (Franklin et al., 2007; Wang et al., 2008). Engineered NPs may also have unpredictable genotoxic properties owing to their small size, high surface area, and charged surface (Singh et al., 2009). A commonly used method for the genotoxicity analyses is Ames test (Mortelmans and Zeiger, 2000; Ng et al., 2010).
The primary objective was to examine the effect of the ZnO NP size on bacterial bioluminescence, seed germination, algal growth, and gene mutation. Laboratory synthesized ZnO NPs of four sizes (5, 15, 25, and 80 nm) were used to investigate the effect of size on the toxicity of each bioassay.
Experimental
Experimental plan
This study aims to examine the toxicity of different sizes of ZnO NPs based on bacterial bioluminescence, seed germination, algal growth, and gene mutation. Table 1 outlines the experimental plan. Details of the experiments are described in the following sections.
NPs, nanoparticles; TEM, transmission electron microscopy; ZnO, zinc oxide.
Synthesis and characterization of ZnO NPs
ZnO NPs of four different sizes, ranging from 5 to 80 nm, were synthesized. The smallest particles (5 nm) were synthesized using a methodology suggested by Lv et al. (2013). A 10 mM zinc acetate solution was prepared by dissolving 0.2 mM zinc acetate dihydrate into 20 mL of absolute ethanol along with a 75 mM NaOH solution. The two well-dissolved precursors were mixed together and stirred at 120 rpm for 12 h. As a result, the mixed solution became turbid with a white precipitate. The turbid solution was then centrifuged to obtain the white precipitate and washed several times with ethanol, which was the as-prepared ZnO nanoparticle sample that was used for further characterization. ZnO NPs (15–80 nm) were synthesized using the sol–gel method (Zak et al., 2013; Raliya et al., 2015b). About 11.2 g of zinc nitrate was dissolved in 25 mL of deionized (DI) water and stirred for 30 min. In addition, 5 g of starch was dissolved in 75 mL of DI water and stirred for 30 min at 75°C, followed by the addition of a zinc nitrate solution to starch solution. Furthermore, the mixture was incubated for 10 h at 80°C under stirring conditions. At this point, the sample was divided into three groups: sample A for 15 nm particles synthesis, sample B for 25 nm particles synthesis, and sample C for an 80 nm particles synthesis. The powders of samples A, B, and C were calcined at 500°C, 750°C, and 900°C, respectively, for 5 h to obtain the ZnO NPs with the desired size. After calcination, the samples were obtained as a white powder. All the synthesized NPs were characterized to determine their morphological features, such as shape and size.
Physical diameter of the synthesized NPs was determined by transmission electron microscopy (TEM; FEI Tecnai G2 Spirit, Hillsboro, OR). In addition to the size, the spread in particle sizes is also reported. Due to the controlled methodologies used, variation in size was minimal, thus allowing us to infer a dependence of the properties on mean size. Further, the synthesized NPs were also characterized by X-ray Diffraction (XRD) for crystal structure confirmation and X-ray Photon Correlation Spectroscopy (XPS) for surface elemental composition. The synthesized ZnO NPs were suspended in DI water (pH 7.2 ± 0.4) and dispersed using ultra-sonication for 10 min prior use to avoid possible agglomeration.
Toxicity of ZnO NPs to bacterial bioluminescence
Escherichia coli strain RB1436 is a DH5 strain obtained from Dr. Burlage (Concordia University, Mequon, WI) (Rogowsky et al., 1987). The plasmid with a constitutive promoter to express the lux genes produces bioluminescence in a growing culture. The strain was used to detect inhibition conditions that result in a measureable decrease in bioluminescent output (Ko and Kong, 2014). For the bioassay, 1 mL of the diluted bacterial suspension was mixed with 9 mL of the ZnO solutions at different concentrations, after which the bioluminescence activity was measured every 0.5 h during incubation. The vials were incubated at 130 rpm and 27°C. The bioluminescence activity was measured using a Turner 20/20 luminometer (Turner Design, San Jose, CA), which had a maximum detection limit of 9,999 relative light units (RLUs). To determine toxicity (EC50), an average of two measurements (1.0 and 1.5 h) was used. The duplicate relative values of the control from each of the two measurement times were averaged.
Toxicity of ZnO NPs to seed germination
We used seeds Lactuca sativa L. for seed germination test due to the importance of the food crops, globally. Before the germination test, all seeds were surface-sterilized in 3% H2O2 for 10 min and then rinsed with distilled water. Filter paper was then placed in a sterilized Petri dish and moistened with 5 mL of an aqueous solution (distilled water for control) containing ZnO. Twenty seeds of each species were placed in each plate, which was covered with a lid and incubated in the dark at 23 ± 2°C. The germinated seeds were counted after 3 days of incubation. An extension of both the plumule and radical to longer than 2 cm from their junctions was considered to be indicative of germination and the seedling root length was measured. Triplicate sets were performed for each treatment.
Toxicity of ZnO NPs to algal growth
Toxicity of the ZnO NPs was assessed using the green algae species, Chlorella vulgaris (KCTC AG10002), obtained from the Korean Culture and Tissue Collection. The algae were cultured for 3 days in BG11 medium at 30°C and 150 rpm under 5,000 lux and diluted to a final OD730 of 0.3 for the toxicity test. For the ZnO NPs inhibition test, 19 mL of the algal culture was mixed with 1 mL of the NP solution (final concentration 0, 0.5, 1.0, and 3.0 mg/L ZnO) and incubated for 3 days at 30°C and 150 rpm. Growth inhibition based on the chlorophyll content and cell counts were measured after 72 h incubation. Chlorophyll was extracted with 90% (v/v) acetone and determined using standard methods. The algal cells were counted with a counting chamber (Marienfeld, German) under an optical microscope.
Toxicity of ZnO NPs to gene mutation
A mutagenicity test was performed using the plate incorporation procedure described by Maron and Ames (1983) with Salmonella typhimurium TA98. The strains were maintained and stored using standard methods (Mortelmans and Zeiger, 2000). A control of 0.05 mL dimethyl sulfoxide or a 0.05 mL sample in 2 mL of top agar (NaCl 6 g, agar 6 g, 0.5 mM histidine/biotin solution 100 mL, and water 900 mL) was placed on a growth medium agar plate with a mixture of 0.01 mL of ZnO NPs solution (distilled water for control), 0.1 mL of the culture, and 0.5 mL of buffer. Each sample was plated in triplicate, the positive revertants counted after 48 h incubation at 37°C. The positive controls used were 2-nitrofluorene (2.5 μg/plate) for TA98 to monitor the sensitivities of the bacterial strains. A 100 mg/L ZnO NPs (1 μg/plate) sample of each size was evaluated for mutagenicity without an S9 mixture (mammalian liver homogenate for metabolic activation).
Statistical analysis
Effective concentration (EC50) (concentration of chemical at which 50% of its effect is observed) values of the NPs used in the treatment were determined by expressing each treatment as a proportion of the control activity. The SPEARMAN computer program, which is distributed by the U.S. EPA's Center for Exposure Assessment Modeling (CEAM), was used to estimate the EC50 values. Statistical analysis of the experimental groups utilized the Student's t-test (www.graphpad.com). Each of the experimental values was compared with the corresponding control. Statistical significance was accepted when the probability of the result assuming the null hypothesis (p) was <0.05.
Results and Discussion
Nanoparticle characterization
Size and morphology of the ZnO NPs were investigated by TEM. Typical TEM images (Fig. 1) showed that the ZnO NPs had a mixed hexagonal and almost spherical morphology. The geometric mean diameter of the samples were 5.3 ± 2.08 nm, 14.8 ± 2.33 nm, 25 ± 1.22 nm, and 82.6 ± 5.64 nm. Do note that for the ease of numbers, in the later figures, the size is represented as 5, 15, 25, and 80 nm, respectively. The 5 nm particles showed a higher degree of agglomeration than the other particles size due to multiple weak interactions and higher surface energy. Therefore, all the samples were sonicated before each toxicity experiment to ensure a near monodisperse distribution in the solvent. The surface chemical composition of the synthesized ZnO NPs was determined by XPS analyses. The dominant peak of Zn 2p3 at 1020.5 eV and oxygen at 529.2 eV is associated with the binding of Zn2+ in the ZnO (Supplementary Fig. S1). The Bruker Diffrac.Eva program was used to evaluate and process XRD scan data. The ZnO NPs showed peaks that match the characteristic zincite crystal peaks (Supplementary Fig. S2).

Morphological characterization of ZnO NPs by TEM.
Effect of ZnO nanoparticle size on bacterial bioluminescence
Effects of the size of ZnO on the bioluminescence activity was investigated at the dose ranges of 0.1, 0.2, 0.5, and 1.0 mg/L for 5 nm and 0.2, 0.5, 1.0, and 1.5 mg/L for the 15, 25, and 80 nm ZnO particles. In addition to mass concentration, as shown in our previous studies, the surface area concentration maybe an important factor (Jiang et al., 2008, 2009). However, in this work, we used the mass concentration to establish an exposure dose range as the size was fixed for each test. The dose range for each size was determined based on preliminary tests. The control produced a mean bioluminescence in the range, 311 to 358 RLU after 1 and 1.5 h of incubation, depending on each set condition. The inhibition of bioluminescence activity was observed with tested sizes of ZnO under the exposed concentrations. A high inhibition rate of bioluminescence was observed at the initial stage of the incubation period. As shown in Fig. 2, after 0.5 h incubation, the relative bioluminescence activity was lowered to the following ranges, depending on the exposed concentration: 13–41% and 0.4–68% for 5 and 80 nm ZnO, respectively. Almost complete inhibition was observed with 1.0 mg/L of 5 nm ZnO (<27 ± 10.2 RLU) and 1.5 mg/L of 15, 25, and 80 nm ZnO (<25 ± 13.1 RLU) after 1.5 h of exposure. To compare the effects of the different sizes of ZnO on the bioluminescence activity, the mean bioluminescence intensities after 1 and 1.5 h of exposure were used to calculate the EC50 values for each size of the ZnO particles. In general, the effect patterns were dependent on the particle sizes and concentration of ZnO. Of the EC50 values (95% confidence level range), the ranking of the toxicity levels in terms of the inhibition of bioluminescence activity was as follows: 0.31 (0.26–0.38) mg/L, 0.37 (0.31–0.44) mg/L, 0.38 (0.31–0.47) mg/L, and 0.40 (0.34–0.46) mg/L for 5, 15, 25, and 80 nm ZnO, respectively. Slightly high inhibition of the bioluminescence activity was normally observed at smaller sizes of ZnO, but no significant differences were observed among the sets with different sizes of ZnO (p > 0.05).
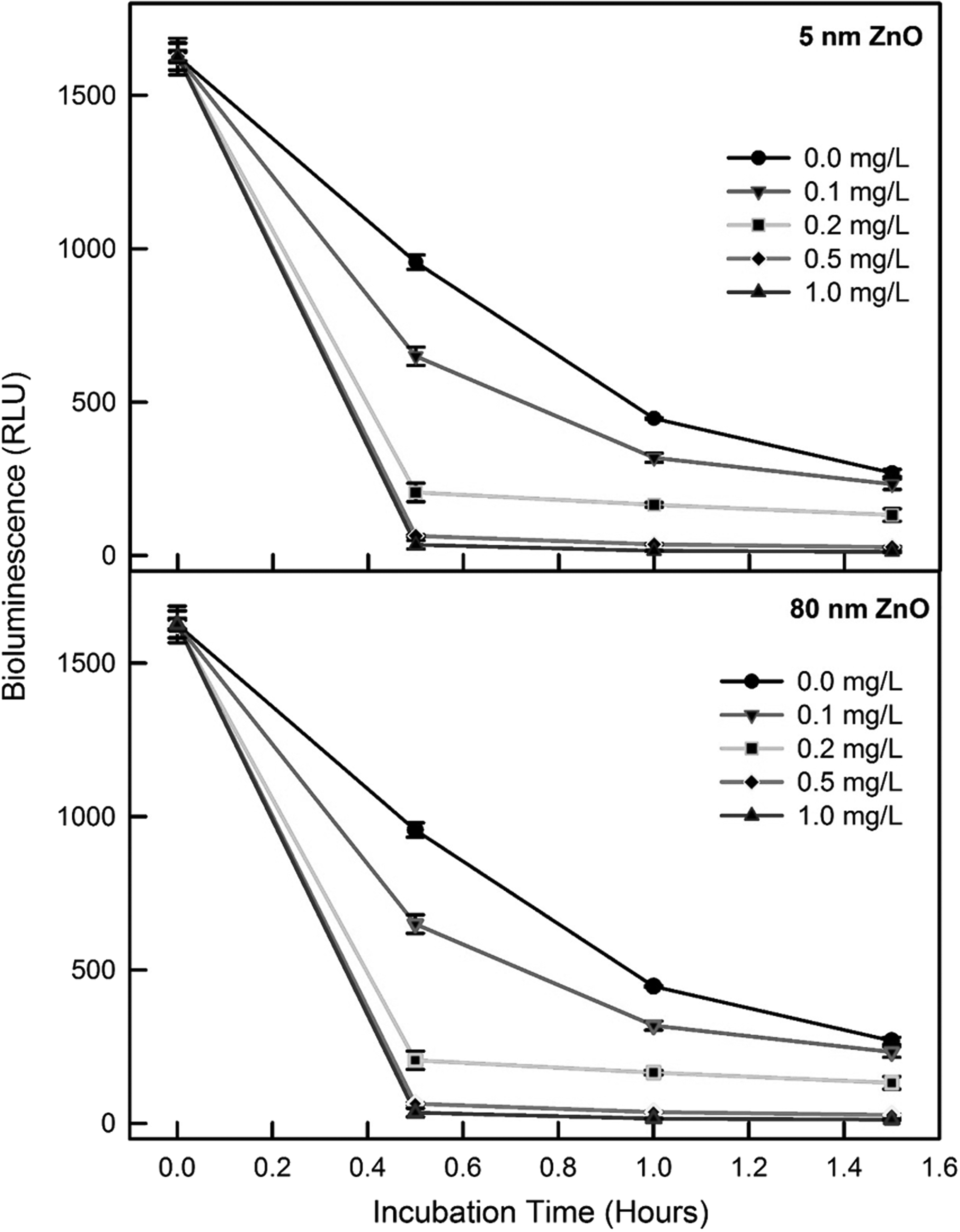
Representative profiles of bioluminescence activity in the presence of 5 nm and 80 nm ZnO NPs. RLU, relative light unit.
Effect of different sizes of ZnO NPs on seed germination
Effects of different sizes of ZnO on seed germination were evaluated using seeds of Lactuca. The dose range of ZnO was determined based on a preliminary range test, 0, 5, 10, 15, and 30 mg/L for all sizes. For the control, approximately 51 (average 17 ± 1.5 per batch of 20 seeds) out of 60 seeds germinated (longer than 2 cm growth) during a 3-day incubation period. In general, less germination occurred with increasing concentrations of ZnO for all the tested sizes (Fig. 3). For example, almost complete inhibition of Lactuca germination occurred with 30 mg/L, whereas the germination ranges of the 5 mg/L ZnO were 72–84% relative to the germination observed in the control group. Inhibitions of seed germination were observed 35 ± 3%, 84 ± 4%, 60 ± 7%, and 76 ± 0% of the control at 10 mg/L of 5, 15, 25, and 80 nm ZnO, respectively. No general patterns were observed according to the ZnO particle sizes. Figure 3A and B, show the EC50 values for seed germination with different sizes of ZnO treatments. The EC50 values (95% confidence level) of the Lactuca seeds were calculated to be 9.98 (8.99–11.08) mg/L, 6.56 (6.08–7.09) mg/L, 8.72 (7.93–9.6) mg/L, and 7.07 (6.52–7.66) mg/L for 5, 15, 25, and 80 nm sizes, respectively. With respect to calculated EC50 values, the 5 nm ZnO showed the lowest inhibitory effect on seed germination compared to the other sizes of ZnO. On the other hand, no statistically significant differences were observed between the results of each size.
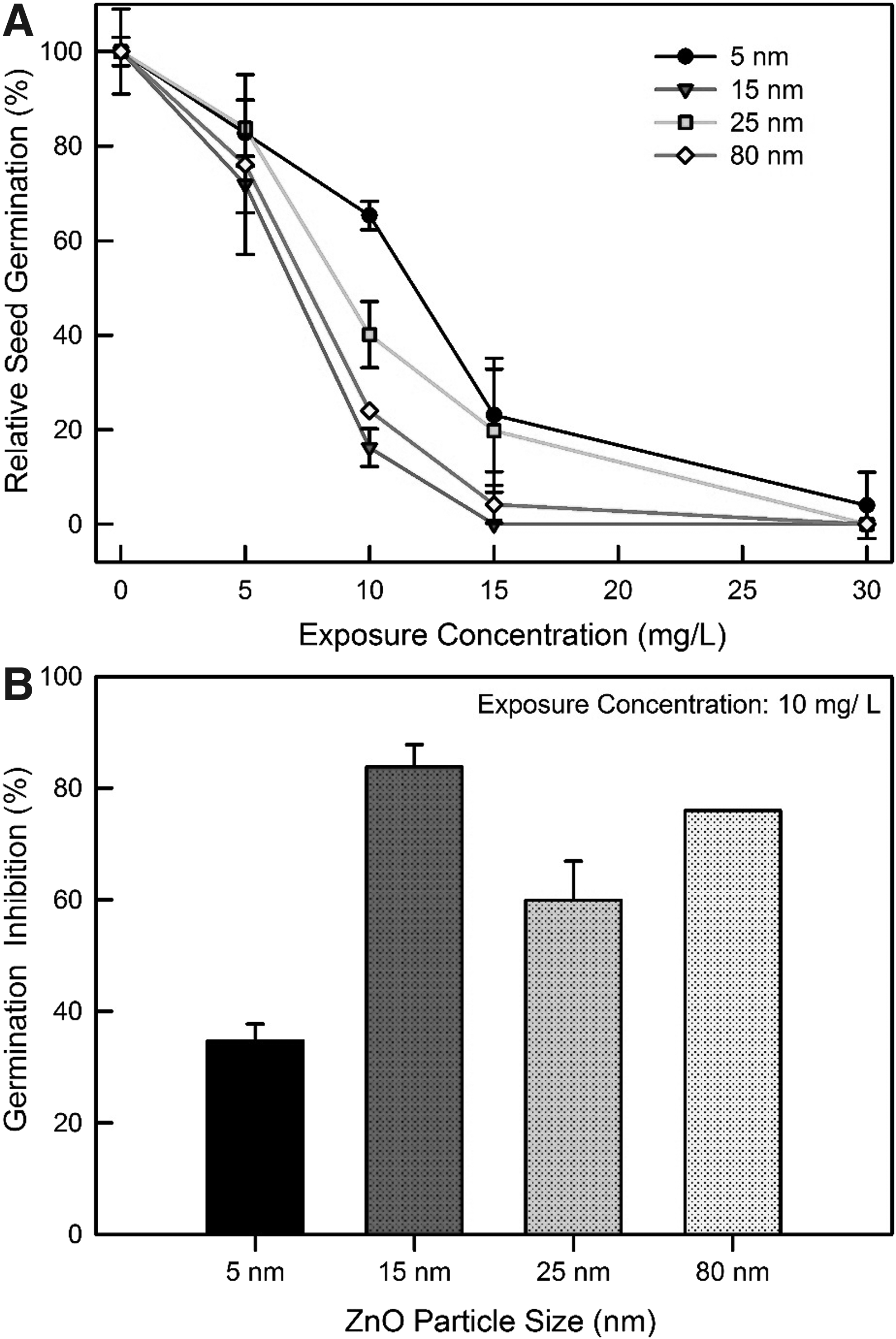
Effects of ZnO NPs size on seed germination after 3 days incubation period:
Effect of different sizes of ZnO NPs on algal growth
The effects of the different sizes of ZnO on algal growth were investigated at the dose range of 0.1–5.0 mg/L for all sizes of ZnO, which was determined based on preliminary tests. Two endpoints, the chlorophyll content and cell counts, were monitored at the end of 3 days incubation. The chlorophyll content and cell counts of the control were in the range, 24,600–33,540 mg/m3 and 3.1–5.0 × 107 counts/mL, respectively, depending on each set condition. Clearly, the effect of ZnO on algal growth was dependent on the particle size and ZnO concentration. For instance, at 0.5 mg/L of 5 nm ZnO, the chlorophyll content and cell count were 59.8% and 56.5% of the control, respectively, whereas 95.8% and 88.6% of the control were observed at 0.5 mg/L of 80 nm ZnO, respectively (Fig. 4). Strong inhibition was observed with 5.0 mg/L of all the ZnO sizes tested, showing 18.4–25.2% and 21.3–38.9% of the control for the chlorophyll content and cell count, respectively (Fig. 4). Of the EC50 values (95% confidence level range), the ranking of the inhibition levels in terms of chlorophyll content and cell count was as follows: 1.14 (0.65–1.98) mg/L and 1.08 (0.56–2.06) mg/L for 5 nm, >1.20 (0.93–1.56) mg/L and 1.35 (0.96–1.96) mg/L for 15 nm, >1.33 (1.03–1.73) mg/L and 1.95 (1.30–2.93) mg/L for 25 nm, and >2.15 (1.74–2.66) mg/L and 2.90 (1.90–4.43) mg/L for 80 nm (Fig. 4). The EC50 value of the 5 nm ZnO NPs was lower (that corresponds to high toxicity) than the larger sizes, showing 0.86, 0.59, and 0.53 times of that of the control for 15, 25, and 80 nm ZnO, respectively. In terms of the EC50 values, significant differences were observed for both endpoints between that of the 5 and 80 nm particle sizes (p < 0.05).
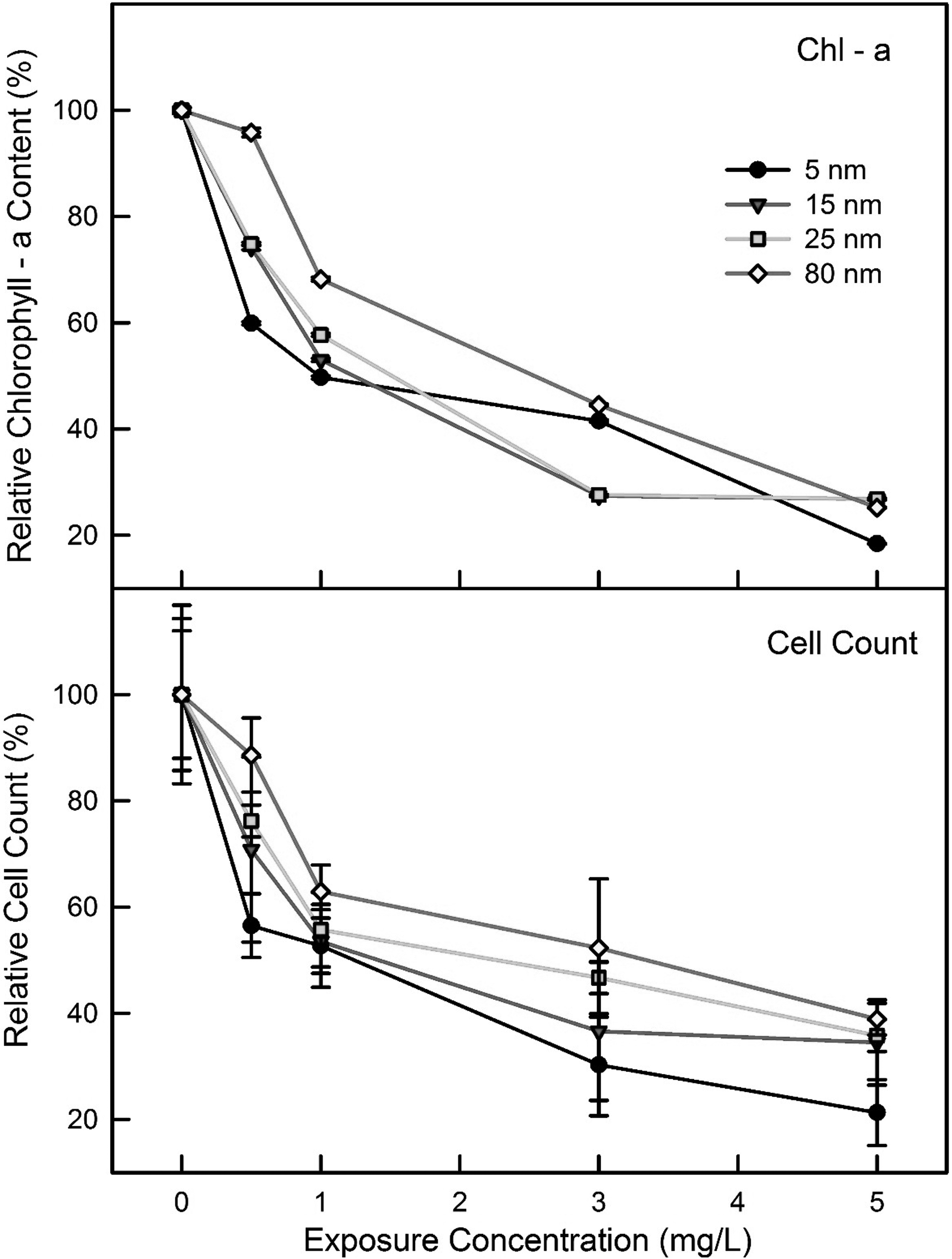
Effects of ZnO NPs size on chlorophyll content and cell count of Chlorella vulgaris.
Effect of different sizes of ZnO NPs on gene mutation
The mutagenicity potentials of the four different sizes of ZnO NPs that showed inhibitory effects against the bioluminescence activity, seed germination, and algal growth were evaluated using the Ames test by employing the mutant strain, S. typhimurium TA98. The effects of the ZnOs were tested at concentrations 100 and 300 mg/L based on the results of a preliminary test. In this investigation, the spontaneous reversion (negative control) rates for TA98 ranged from 50 to 61 colonies. High reversion activity (positive control) was observed with up to 10 μg/L of 2-nitrofluorene using strain TA98, showing approximately more than 300 CFUs (colony forming units). Various ranges of CFU (from 50 to 120 CFUs) were observed at 100 and 300 mg/L of ZnO. The mutation ratios (MRs) of the tested sizes of ZnO ranged from 1.0 to 2.4 under the two concentrations tested (Fig. 5).

Comparison of the effects of particle sizes of ZnO on mutation ratios (sample/negative control) of the TA98 strain.
The aim of this research was to determine the ecological toxicity of different particle sizes of ZnO based on the activity of commonly used bioassays because of their simplicity, controllability, and reproducibility. We tested NPs impact on four bioassays, namely (1) bioluminescence production of recombinant E. coli strain RB1436, (2) germination activity of seed Lactuca, (3) chlorophyll contents and cell count of C. vulgaris algal growth, and (4) genetic revertant mutations using a mutant strain of S. typhimurium (Ames test). The particle size corresponds to the surface area to volume size ratio that affects the NPs interactions with organisms. Overall, the present result of the size-based toxic study of the ZnO NPs indicated that with decreasing particle size, the toxicity increased for seed germination and algal growth, decreased for bacterial gene mutation, while remained relatively unchanged for bacterial bioluminescence, depending on the tested bioassays. No single bioassay showed uniform toxicity or sensitivity among the ZnO sizes tested. The observation suggests that any knowledge of the NP toxicity using only one kind of bioassay or size has limited benefits, therefore, it is important to correlate multiple bioassays to understand the environmental interaction and impact of engineered NPs.
In the bioluminescence test, an observable inhibition of bioluminescence activity was observed in the range, 23–43% of the control at 0.2 mg/L ZnO for all sizes. Of the highest concentrations tested (1.5 mg/L), almost complete inhibition of the bioluminescence activity was observed for all sizes. With respect to the EC50s, the toxicity ranking of the different sizes of ZnOs was as follows (EC50s mg/L): 5 nm (0.31) > 15 nm (0.37) > 25 nm (0.38) > 80 nm (0.40). Although slightly high toxicity (corresponds to low EC50 value) appeared as the particle size decreased in terms of EC50, no significant differences were observed between the sizes (p > 0.1167). Heinlaan et al. (2008) reported EC50 values of 1.9 mg/L for ZnO NP in terms of the inhibition of the bioluminescence activity of V. fischeri, which was approximately four to five times higher (less inhibition) than this investigation. Regarding the bacterial toxicity, ZnO has been shown to induce reactive oxygen species (ROS), which can cause membrane damage and antibacterial effects in E. coli, induced by lipids, carbohydrates, proteins, and DNA damage (Kohen and Nyska, 2002; Zhang et al., 2007). Many researchers have reported the size-dependent toxicity of NPs (Carlson et al., 2008; Choi and Hu, 2008). Generally, smaller particles, leaving a higher number of atoms exposed on the surface available for redox, photochemical, and biochemical reactions in addition to physicochemical interactions with cells, adhere readily to the biological components and generate more notable adverse effects (Marambio-Jones and Hoek, 2010; Xiang et al., 2015). Morones et al. (2005) also reported that the bactericidal properties of the NPs are size dependent, because only NPs that present a direct interaction with the bacteria preferentially have a diameter of ∼1–10 nm. Heinlaan et al. (2008) reported that metal ions from metal oxide NPs may be responsible for the observed antibacterial effects because bacteria are largely protected against NP entry (no transport mechanisms for supramolecular and colloidal particles).
In the present study, Lactuca seeds were chosen to investigate the seed germination in the presence of different size of ZnO nanoparticles. The effects of chemicals on seed germination are generally dependent on their ability to reach the embryonic tissues across the physiological barriers, which are predominantly the seed coat (Akinci and Akinci, 2010). This ability is dependent directly on a seed's coating structure, which varies among plant species and changes according to the physical and chemical properties of the pollutants (Seregin and Kozhevnikova, 2005). Starch in seeds is degraded by a range of enzymes, such as amylase for germination. Therefore, the inhibition of specific enzymatic reactions by NPs could explain the toxicity of NPs. Conflicting results on the inhibitory effects of elevated concentrations of NPs on germination of seeds from different plant species have been reported. Lin and Xing (2007) observed the significant inhibition of radish, rape, and ryegrass seed germination by ZnO NPs with EC50 values ranging from 20 to 50 mg/L, and they reported no inhibitory effects by the dissolved Zn ions from the NPs on seed germination or growth. In the present investigation, the effects of ZnO NPs on Lactuca seed germination (EC50) was approximately four to five times lower (corresponds to high toxicity) than that reported by Lin and Xing (2007), showing in the range of 6.56–9.98 mg/L, depending on the particle sizes. The present work showed that the effects of ZnO on seed germination activity varied according to the ZnO sizes, even though no consistent patterns of toxicity were observed for all particle sizes.
In contrast to the bioluminescence bioassay, there were trends indicating that smaller particles were less toxic than larger particles on seed germination, showing EC50 (95% confidence level) values of 9.98 (8.99–11.08) mg/L, 6.56 (6.08–7.09) mg/L, 8.72 (7.93–9.6) mg/L, and 7.07 (6.52–7.66) mg/L for 5, 15, 25, and 80 nm ZnO, respectively. The toxicity (EC50) for the 80 nm ZnO NPs was 0.71 times lower than that of the 5 nm ZnO NPs, indicating more toxic effects by larger particle sizes on seed germination. Statistically significant differences, based on a Student's t-test, were also observed between 5 and 80 nm (p-values of 0.0112). Thuesombat et al. (2014) reported that increasing the size of Ag NPs over the 20–150 nm diameter range increased the inhibition effect upon seed germination and seedling growth, which were consistent with these investigations. Information on the NP uptake by plants and their mechanisms of phytotoxicity is largely unknown. The potential mechanisms of NP toxicity could include the dissolution of ions, NP accumulation, increased ROS production upon contact, and subsequent lipid oxidation of the cellular membranes (Stampoulis et al., 2009). Interestingly, Lee et al. (2008) reported that Cu ions released have little impact on plants and that their dose-dependent toxicity is specifically due to NP accumulation within cells. Further studies should examine the toxicity mechanisms of NPs on a range of response endpoints of plants (e.g., root and shoot length, biomass, and enzyme activity) with respect to the size distribution relative to phytotoxicity, uptake, translocation, and rhizosphere, and root surfaces because no assessment using just one seed species and endpoint can fully elucidate the general toxicity of NPs to the plants (Raliya and Tarafdar, 2013; Raliya et al., 2015a, 2015b; Raliya et al., 2016).
Various species of algae have been used as standard toxicity tests to assess the environmental contaminants (Crane et al., 2008). In this investigation, the effects of different sizes of ZnO were assessed based on the chlorophyll contents and cell counts of C. vulgaris. With respect to the average EC50s of the two endpoints, the toxicity ranking of the different sizes of ZnO of algal growth was as follows (EC50s mg/L): 5 nm (1.11) < 15 nm (1.28) < 25 nm (1.64) < 80 nm (2.53). The toxicity of the 5 nm ZnO NPs was ∼2.3 times higher than that of the 80 nm ZnO NPs, indicating more toxic effects by the smaller particle sizes on algal growth (p-values of 0.05). Overall, in terms of the EC50s, the effect of ZnOs on the chlorophyll content revealed a slight sensitivity compared to cell count, showing 0.8 times of the EC50 of the cell count. On the other hand, no statistical significant differences were observed between the results of the two endpoints, indicating that one or both observations could be used for the assessment. In the case of algae, the cell walls are the primary site for the interaction and a barrier for the entrance of NPs, where the mechanisms for entrance are unclear (Navarro et al., 2008). The diameter of pores across the cell wall generally ranges from 5 to 20 nm; thus, only NPs with a size smaller than that of the largest pore are expected to pass through the cell wall (Moore, 2006). As soon as the NPs enter the cell, they may bind with different types of organelles and interfere with the metabolic processes, possibly producing ROS (Navarro et al., 2008). Previous studies reported that the toxicity of ZnO NPs may be partially attributed to the solubility and dissolved Zn of the ZnOs (Franklin et al., 2007; Aruoja et al., 2009). The ionic dissolution of NPs depends on the particle size (Wong et al., 2013; Lopes et al., 2014). In addition, NPs aggregate in the media and are entrapped and wrapped around the algal cells, which may contribute to growth inhibition (Ji et al., 2011). The toxicity of the ZnO NPs toward algae Scenedesmus obliquus was strongly dependent on the initial particle sizes, the exposure concentration, and the irradiation conditions (Bhuvaneshwari et al., 2015). Smaller sized NPs with a greater surface area could effectively interact, easily stick to/cross the membrane and become internalized into the algal cells. A previous study also noted that the smaller sized ZnO have a more adverse effect on marine algae than the larger particles (Khare et al., 2011; Peng et al., 2011).
In the gene mutation test, spontaneous reversion for TA98 ranged from 30 to 50 colonies, which is slightly less than that reported previously (Mortelmans and Zeiger, 2000; Mathur et al., 2007). In this study, mostly no significant mutagenicity with the strain TA98 was observed under the tested concentrations (ratio >2.0 indicates positive mutagenicity), indicating most of the ZnO sizes under the tested concentration had no mutagenic effects under the tested conditions. At 100 mg/L of 5 and 15 nm ZnO, however, a slightly high MR (>2) was observed, showing 2.4 and 2.03, respectively. Singh et al. (2009) reported that bacterial mutagenicity assays (Ames test) may be unsuitable for measuring genotoxicity induced by the NPs because prokaryotes lack the ability to perform endocytosis and the NPs may be unable to diffuse across the bacterial cell walls; this lack of uptake ability could potentially lead to false negative results. Several instances of conflicting results concerning the genotoxicity of NPs using different assays have been reported. For example, both the comet and micronucleus assays indicated that TiO2 NPs induce DNA damage, whereas the Ames test and chromosome aberration assay reported negative results (Kang et al., 2008). Maenosono et al. (2007) reported that bacterial gene mutation assays using NPs seldom produce positive results due to a lack of penetration of the test material through the bacterial cell wall. Therefore, the actual uptake of the test material into the bacterial cells needs to be confirmed. On the other hand, Kumar et al. (2011) reported that the larger sized ZnO and TiO2 NPs caused more cell DNA damage to bacteria than the smaller ones. In general, the key mechanism responsible for the genotoxic effects of NPs involves oxidative stress, which refers to a redox imbalance within the cells resulting from the increased intracellular ROS and decreased antioxidant levels. In addition, one genotoxicity test cannot cover all the potential forms of genetic damage that may arise by nanoparticle exposure. Therefore, multiple toxicity tests from genetic to cellular level using various organisms are required to understand the toxic or biocompatibility profile of nanomaterials.
In this study, the toxicity rankings were clearly differed depending on the tested methods: bacterial bioluminescence > algal growth > seed germination >> gene mutation. Several mechanisms could be responsible for the different toxicity of NPs in each organism (Jiang et al., 2009; Karakoçak et al., 2016). Although the precise toxic mechanisms of the metal oxide NPs are largely unknown (Griffitt et al., 2008; Menard et al., 2011), however, their toxic effects can be attributed to two different actions: (1) chemical toxicity based on the chemical composition, for example, release of (toxic) ions; and (2) oxidative stress or stimuli induced by the surface, size, and/or shape of particles, resulting in damage to lipids, carbohydrates, proteins, and DNA (Crane et al., 2008; Navarro et al., 2008; Hu et al., 2010).
Conclusion
Toxicity of NPs in the environments has shown large variability in laboratory tests due to the reactions with the constituents of environmental matrices. Therefore, there is a need for standardized methods for assessing the effects of NPs exposure, particularly under field conditions. This work showed that each of the bioassays (bacterial bioluminescence, seed germination, algal growth, and gene mutation) has a specific sensitivity to ZnO NPs, and different effects regarding the particle sizes of ZnO. The results suggest that a combination of various bioassays may constitute a better way for assessing the influence of NPs on the environment rather than just one test species.
With the rise in the applications of NPs in various fields, including agriculture, their potential toxicity on the environment is capturing great attention. The eventual toxicity to a specific organism depends on various characteristics of the NPs such as size, shape, exposure concentration, and composition. Further, nanoparticle-induced toxicity may be systematic or localized at the cellular and tissue level, and it may also cause genotoxicity. In this investigation, we performed controlled studies with different sizes of ZnO NPs and examined their impact on bacterial bioluminescence, seed germination, gene mutation in bacteria, and algal growth. The implication of research finding to both adverse and useful environmental impacts is discussed.
Footnotes
Acknowledgments
This research was supported by the Basic Research Project of the Korea Institute of Geoscience and Mineral Resources (KIGAM) funded by the Ministry of Science, ICT and Future Planning of Korea. Also, this research was partly supported by the National Research Council of Science and Technology (NST) grant by the Korea government (MSIP) (No. CAP-17-05-KIGAM). The authors acknowledge the support of the Nano Research Facility (NRF), Washington University in St. Louis for their help in nanoparticle synthesis and characterization.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.