
Abstract
Abstract
To investigate the toxicity of ZnO nanoparticles (NPs) on microalgae, Bacillariophyta (Skeletonema costatum and Phaeodactylum tricomutum) and Chlorophyta (Dunaliella salina and Platymonas subcordiforus) were cultivated in f/2 medium under uniform culture conditions with different concentrations of ZnO NPs. Growth inhibition ratio and the 50% effective concentration were used as two toxicity parameters for comparing toxicities on different microalgae. Dose–response curves were built to show different trends in toxicities of ZnO NPs on the four microalgae. The sensitivity rank of the algae to toxicity of ZnO NPs was S. costatum>D. salina>P. tricomutum>P. subcordiforus. Adsorption of ZnO NPs on S. costatum and D. salina was clearly observed in scanning electron microscope images. In contrast, no adsorption of ZnO NPs was found on surfaces of P. tricomutum and P. subcordiforus. Shapes and surface structures of the microalgae resulted in different adsorption phenomena, which had an influence on interactions between NPs and microalgae. This study helps to explain the different toxicities of ZnO NPs on the four microalgae, which could contribute to further explore toxic mechanisms of NPs.
Introduction
W
Studies have reported that ∼25% of sunscreen-incorporating materials, such as ZnO and titanium dioxide (TiO2) NPs, were washed off during bathing and swimming, implying that ∼250 tons of these nanomaterials are potentially discharged into marine environments (Danovaro et al., 2008). Oceans are the final destination for all waste NPs (Moreno-Garrido et al., 2015), and coastal systems are likely the ultimate sink for many nanomaterials, deliberately discharged into the environment (Klaine et al., 2008). There are many species of microalgae in coastal systems, allowing easy access to studies on the ecotoxicology of these NPs.
ZnO NPs were reported early in the literature as emerging environmental contaminants. Toxicities of NPs are affected by the pH, ionic strength, and type of culture media (Aravantinou et al., 2015). However, even under similar experimental conditions, ZnO NPs exhibited significantly different toxic effects on different species of microalgae. For example, for the same treatment of the ZnO NPs, the medium lethal concentration (LC50) for Skeletonema costatum was 2.36 (2.11–2.64) mg/L and for Thalassiosira pseudonana was 4.56 (4.06–5.11) mg/L in an f2-Si medium (Wong et al., 2010). Zn2+ released by ZnO into the medium and ZnO going into algal cells by transmembrane are two of main toxic mechanisms for ZnO NPs, which have been reported by many studies (Franklin et al., 2007; Miao et al., 2010; Zhang et al., 2016). The adsorption of ZnO NPs by algal cells is a potential toxic mechanism, which has been typically ignored (Hartmann et al., 2012). Moreover, there has been a lack of studies reporting the clear observation of the adsorption of ZnO NPs on the surface of algal cells. The adsorption of ZnO NPs on algal cells should receive more attention to further explore the toxic mechanisms of ZnO NPs.
To be consistent with our previous studies, four species of marine algae—Bacillariophyta (S. costatum and Phaeodactylum tricomutum) and Chlorophyta (Dunaliella salina and Platymonas subcordiforus)—were selected as test organisms. It was convenient to compare toxicities between green algae and diatoms, as these four species are easily cultured in the laboratory and sensitive to many toxic substances. As a euryhaline and eurythermal diatom, S. costatum is representative of an organism that is widespread in the ocean. It also has a rough surface and the potential to interact with particles in media. P. tricomutum has a slender shape and the highest cell density of the four species, which was favorable to decrease the adsorption of the NPs. D. salina, without a cell wall, is a model organism for toxicity tests. This organism allowed us to investigate whether the cell wall influenced the adsorption of the NPs. P. subcordiforus was a common local alga representing local characteristic species. In this study, algal growth inhibition tests were carried out to explore toxicities of ZnO NPs on the four microalgae. The growth inhibition ratios (IRs) and 50% effective concentration (EC50) were calculated to compare toxicities of ZnO NPs on different algae. Scanning electron microscope (SEM) images provided clear observations of the adsorption of ZnO NPs on the algae. To the best of our knowledge, this was the first time the adsorption of ZnO NPs on algal surfaces was clearly observed by SEM.
Materials and Methods
Chemicals
Nanopowders of ZnO (CAS:1314-13-2, 99.8% metals basis, 50 ± 10 nm) were purchased from Aladdin Industrial Corporation, China. ZnO NPs did not disperse well in the f/2 medium, and even after sonicating, they still aggregated. Dry powders (A) and nanopowders of ZnO dispersed in f/2 medium (B) were observed under a SEM, as shown in Fig. 1.
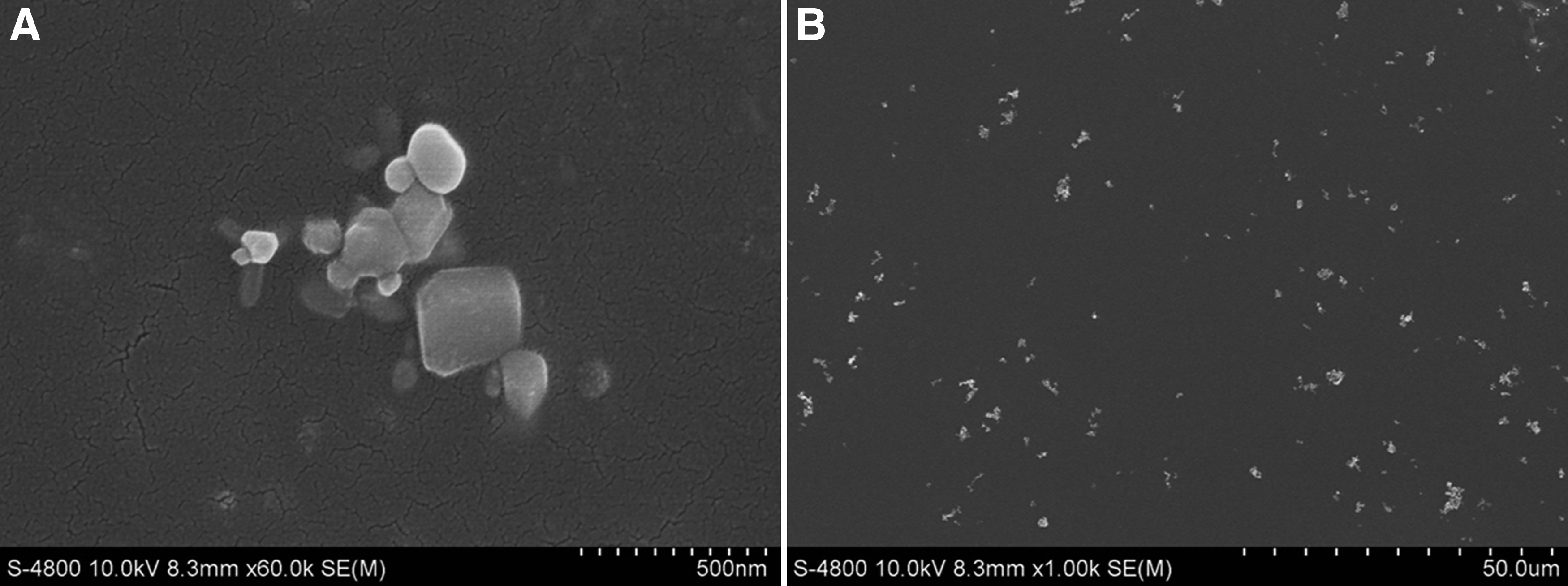
SEM image of dry zinc oxide powders
Organisms
Four algal strains, S. costatum (Bacillariophyta), P. tricomutum (Bacillariophyta), D. salina (Chlorophyta) and P. subcordiforus (Chlorophyta) were obtained from the Algal Center of Key Laboratory of Marine Chemistry Theory and Technology, Ocean University of China. All microalgae were cultivated in an f/2 medium (Guillard and Ryther, 1962) made with sterile seawater (filtered by a 0.45 μm membrane) from Qingdao, China. The microalgae were cultivated in a 5-L Erlenmeyer flask at 20°C ± 1°C under cool, continuous white fluorescent lights (4,000 lux) with a 12-h light–12-h dark cycle, and shaken twice per day to prevent the sedimentation of the algae.
Preparation of ZnO NPs suspension
A stock solution of ZnO NPs was prepared by dispersing dry nanopowders of ZnO into milli-Q water to a final concentration of 100 mg/L. With shaking and sonication, the well-dispersed stock solution could be stored for a week at room temperature. The stock solution was diluted with milli-Q water, and then dispersed with an ultrasonic bath for 30 min at 50 W before the toxicity assay.
Algal growth inhibition test
All tests in the study complied with the Organization for Economic Cooperation and Development (OECD) Guidelines 201. All glassware were acid soaked, cleaned with milli-Q water, and then autoclaved before test. The 100-mL precultured algal cells (precultured 5–7 days until log phase growth prevailed) were transferred into a 250-mL Erlenmeyer flask, and then, the suspension of ZnO NPs at various concentrations was added into the test flask in triplicate. The final concentrations of ZnO NPs in all algal growth inhibition tests were 0, 0.5, 1, 3, 5, and 10 mg/L to allow for easy intercomparison of the toxic effects on different species. The test flasks were randomly placed in the growth incubator for 4 days in accordance with precultured conditions. Cell densities were counted every 24 h, and 4-day growth curves were plotted. Cell densities were measured by a hemocytometer under an inverted microscope with 400 × magnification. Cell densities of every sample were counted three times, and results were expressed as an average ± standard deviation.
Sample preparation for SEM
Adsorption of ZnO NPs on microalgae was observed with a SEM (Hitachi S-4800 cold field emission). After a 24 h-exposure under a 10 mg/L treatment of ZnO NPs, the algal cells were collected by centrifugation (3,000 rpm, 10 min), and cells were fixed by 2.5% glutaraldehyde at 4°C overnight. The samples were then washed three times with a 0.1 mol phosphate-buffered solution (PBS, pH 7.4) followed by centrifugation (3,000 rpm, 10 min). Samples were successively dehydrated through a series of alcohol solutions (30%, 50%, 80%, 90%, and 100%) for 20 min. After dehydration, samples were fixed with tert-butyl alcohol and then freeze dried for the final observation.
Data analysis
One-way analysis of variance, with a comparison to Duncan's multiple-range test (using the software SPSS 19), was applied to test for significant differences among different treatments (the significance level was always set at p < 0.05). Data were expressed as an average ± standard deviation. The growth IR was calculated as: IR(%) = (1 − T/C) × 100%, where T and C were cell densities in the experimental group and control group, respectively. The EC50 value was calculated by the software SPSS 19 with a 95% confidence interval.
Results and Discussion
Inhibition of algal growth
To account for different initial cell densities and growth habits of different species, IR was used instead of cell densities to visualize toxicity comparisons. Algal growth was affected in various degrees by the ZnO NPs that were added into the medium.
IR of S. costatum increased rapidly with an increasing concentration of ZnO NPs added in the medium, as shown in Fig. 2A. As time increased, the IR of S. costatum also increased, reaching 67.3% under 10 mg/L concentration of ZnO NPs over 96 h. The IR of S. costatum was significantly different (p < 0.05) than that of the control group, even under the minimum concentration (0.5 mg/L) of ZnO NPs over only 24 h, which suggested that S. costatum was sensitive to ZnO NPs. As shown in Fig. 2B, the IR of P. tricomutum also increased with increasing concentrations of ZnO NPs, in a similar trend as S. costatum. However, with time, the IR of P. tricomutum increased over 48 h, but then decreased up to 72 and 96 h. The peak IR of P. tricomutum was 56.8%, occurring at 48 h under 10 mg/L concentration of ZnO NPs. The IR of P. tricomutum under 0.5 mg/L concentration of ZnO NPs at any time showed no significant difference (p > 0.05) from the control group. The IR under 1 mg/L concentration of ZnO NPs over 96 h showed a significant difference (p < 0.05) from the control group. P. tricomutum was not as sensitive to ZnO NPs compared with S. costatum. D. salina showed similar results as S. costatum, as shown in Fig. 2C, but the increment of IR over time and the concentration of ZnO NPs were both less than those for S. costatum. The peak IR of D. salina was 61.3%, occurring at 96 h under 10 mg/L concentration of ZnO NPs. The toxicity of ZnO NPs on P. subcordiforus was quite different from toxicities on all other species. Its maximum IR was only 24.2%, occurring at 96 h under 10 mg/L concentration of ZnO NPs. Except for 10 mg/L concentration of ZnO NPs, other concentrations of ZnO NPs showed the IR of P. subcordiforus with little difference (p > 0.05) to the control group, as shown in Fig. 2D. P. subcordiforus had the highest tolerance to toxicities of ZnO NPs among the four species.
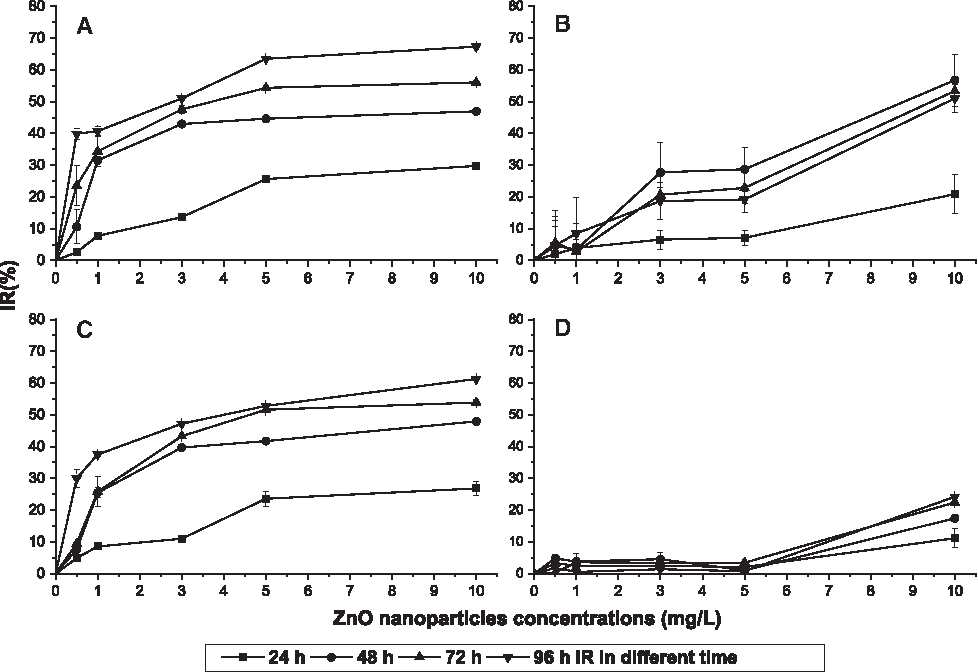
Growth inhibition ratios of algae under different concentrations of ZnO nanoparticles at 24, 48, 72, and 96 h.
In summary, S. costatum was sensitive to the exposure to ZnO NPs, whether under a low concentration of ZnO NPs or during early exposure times, and it had the highest IR of four species. S. costatum was the most vulnerable species in this study.
EC50 of ZnO NPs on algal species
As a practical toxicity parameter, EC50 is widely used in toxicity tests for comparison purposes. The probit analysis program in the software SPSS 19 was used to calculate the values of EC50 as shown in Table 1.
Probit analysis was used to calculate the values of EC50 by the software SPSS 19.
Data were not given by the statistical software.
The EC50 of S. costatum and D. salina showed a similar trend. Both of these values decreased with time and exhibited minimum value at 96 h. The lowest EC50 of S. costatum was lower than that of D. salina, which indicated that S. costatum was more vulnerable than D. salina. The EC50 of P. tricomutum increased with time, and its overall value was larger than that of S. costatum or D. salina. Due to the low IR of P. subcordiforus, the statistical software could not provide its value of EC50. In accordance with the results of algal growth inhibition tests, S. costatum had the lowest EC50, and therefore, it was the most vulnerable species compared with the other algae. In previous studies, D. salina had been used to evaluate the toxic effects of ZnO NPs on algae, and the reported value range of EC50 agreed with results in this study (Manzo et al., 2013; Aravantinou et al., 2015). Peng et al. (2011) studied toxicities of four types of ZnO NPs on three species of algae and found that P. tricornutum was the least sensitive to ZnO NPs of all the species. In this study, the sensitivity rank of the algae to the toxic effects of ZnO NPs was S. costatum>D. salina>P. tricomutum>P. subcordiforus.
Adsorption of ZnO NPs on microalgae
Although the experiment conditions were the same for all four species of microalgae, the species had different sensitivities to ZnO NPs based on different shapes and characteristics. The adsorption of ZnO NPs on microalgae was studied to investigate the interactions between ZnO NPs and microalgae. D. salina adsorbed ZnO NPs on its surface (Fig. 3C, D). Some of the ZnO NPs gathered to small aggregates, and these aggregates still adsorbed onto the surface of D. salina. When the concentration of ZnO NPs was high enough, the NPs could cover parts of the algal surface and limit the transfer of energy and substances between cells and the environment (Tang et al., 2014). Furthermore, many damages were observed on the surface of D. salina (Fig. 3B), which were not observed on the control (Fig. 3A) or on the other three microalgae. The cell wall usually protects algae from environmental stress, thereby enhances the adaptability of algae. As D. salina does not have a cell wall, it is vulnerable to physical damages caused by aggregation or physical processes (Schwab et al., 2011). Harmful substances, such as ZnO NPs, can easily enter the cell without a cell wall and influence the growth of the algae.
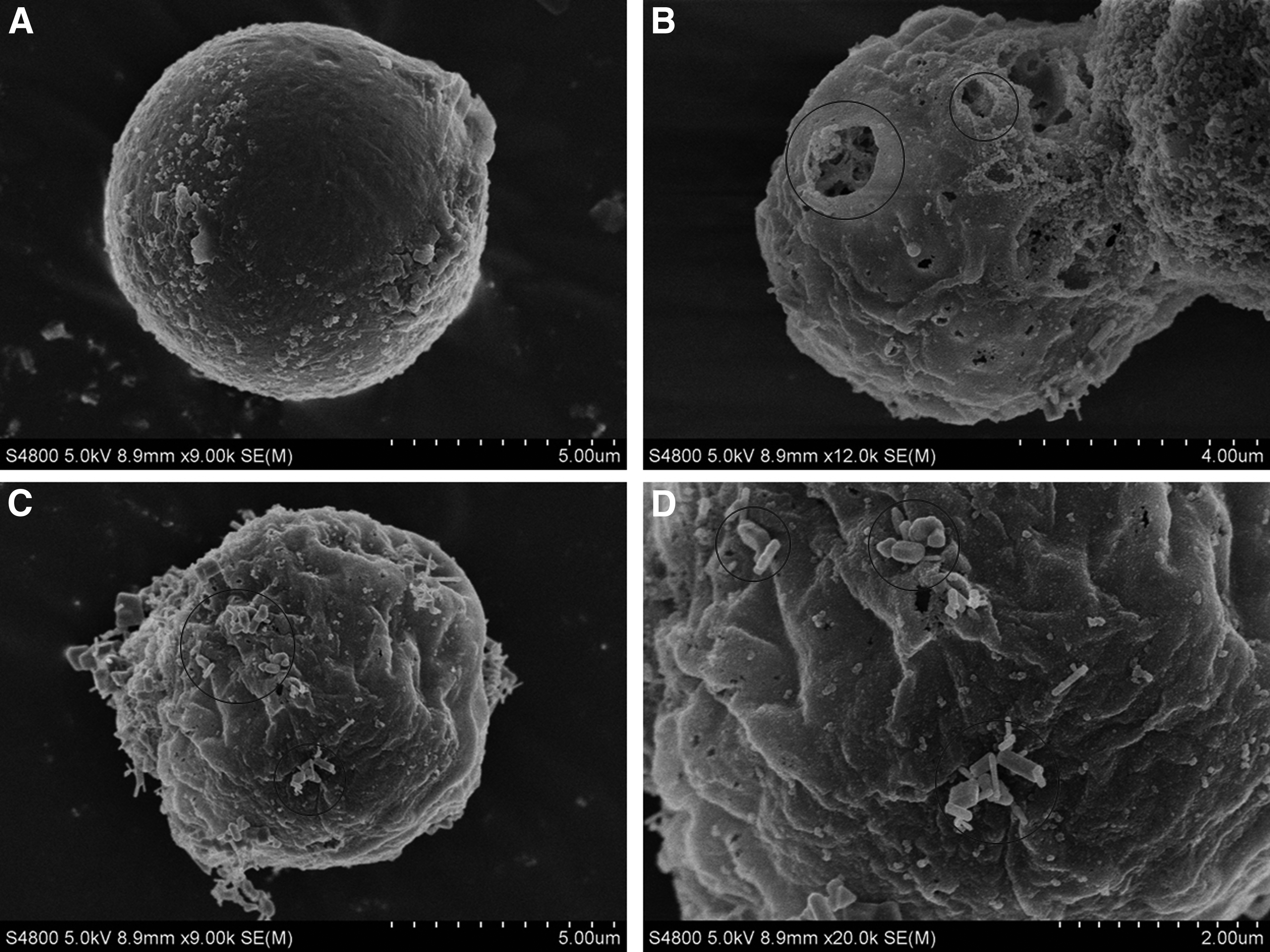
SEM images of adsorption of ZnO NPs on D. salina with a treatment of 10 mg/L of ZnO NPs after 24 h exposure.
Adsorption of ZnO NPs on the surface of S. costatum was also observed (Fig. 4A, B). There were many caveolae on the rough surface of S. costatum, and the ZnO NPs could easily lock into these small caveolae. With the ZnO NPs adsorbed on caveolae, zinc could penetrate the cell wall by either a transmembrane process or the release of zinc ions into cells (Manzo et al., 2013; Zhang et al., 2016). By either route, zinc entering the cell has a negative influence on the growth of the microalgae (Zhang et al., 2016). Silicic thorns were observed on the surface of S. costatum (Fig. 4A). ZnO NPs that adsorbed onto the thorns have the potential to limit the movement of the algae, thereby, affecting its growth (Bhattacharya et al., 2010). If the concentration of NPs is high enough, the NPs could encapsulate the whole microalga and completely inhibit its growth (Li et al., 2015). In this study, the desalination and dehydration processes in sample preparation could wash off some of ZnO NPs with the PBS and alcohol. The observed amounts of NPs adsorbed onto S. costatum in Fig. 4 may be lower than the actual amount in the absence of these sample preparation steps.

SEM images of adsorption of ZnO NPs on S. costatum with a treatment of 10 mg/L of ZnO NPs after 24 h exposure.
There were no ZnO NPs observed on the surface of P. tricomutum and P. subcordiforus (Fig. 5A, B). P. tricomutum showed a high growth rate, supporting its resistance to the toxicity of ZnO NPs. Its long and thin shape could also decrease the interactions between ZnO NPs and the algae (Fig. 5A). The release of exopolymeric substances could also increase the tolerance of P. tricomutum to contaminants (Miao et al., 2010). The smooth surface of P. subcordiforus could make it difficult for ZnO NPs to adsorb (Fig. 5B). The surface charges of P. tricomutum and P. subcordiforus could also affect the adsorption of ZnO NPs on the surface (Bhattacharya et al., 2010). During the experiments, the shape of P. subcordiforus changed slightly because the microalga was not fixed by osmium tetroxide. Osmium tetroxide can dissolve ZnO NPs, so this process was not used in this article.

SEM images of adsorption of ZnO NPs on P. tricomutum and P. subcordiforus with a treatment of 10 mg/L of ZnO NPs after 24 h exposure.
Conclusion
Algal growth inhibition tests were carried out to investigate toxicities of ZnO NPs on four species of marine microalgae. For toxicity comparisons, IR and the toxicity parameter EC50 were calculated, and a dose–response curve was built. In this study, the sensitivity rank of the microalgae to toxic effects of ZnO NPs was S. costatum>D. salina>P. tricomutum>P. subcordiforus. The adsorption of ZnO NPs on S. costatum and D. salina was clearly observed in SEM images. No adsorption of ZnO NPs was found on the surfaces of P. tricomutum and P. subcordiforus. The shapes and surface structures of the microalgae resulted in different adsorption phenomena related to the different toxicities of ZnO NPs on the four microalgae.
Footnotes
Acknowledgments
Four species of algae were provided by the Algal Center of Key Laboratory of Marine Chemistry Theory and Technology, Ocean University of China. This study was supported by the Public Science and Technology Research Funds Projects of Ocean (201505034), and the National Program on Global Change and Air–Sea Interaction (GASI-03-01-02-01).
Author Disclosure Statement
No competing financial interests exist.