
Abstract
Abstract
Methanotrophy has become a promising process for biotechnological applications. The initial goal of this study was to investigate methanotrophic denitrification as an alternative to nitrogen removal in waters that have a low organic content. Sequencing batch reactors (SBRs) were inoculated with a blend of marine and mangrove sediments (Sed-SBR), or anaerobic granular sludge from a upflow anaerobic sludge bed reactor treating poultry slaughterhouse wastewater (AnS-SBR) immobilized in polyurethane foams under anoxic conditions. Methane was provided as the sole external electron donor and organic carbon source. Methane supported denitrification to a small extent (<2.0 mg N.g total volatile solids−1 d−1) and supported biomass growth for the AnS-SBR in 10% of the initial volatile suspended solids. DNA sequencing of initial inocula showed high richness and low dominance for Sed inoculum, while the most abundant microorganisms in AnS inoculum were chemoorganotrophs typically found in anaerobic environments. In initial inocula, reads belonging to methanotrophs accounted for <0.5% of the sample. In enriched samples, aerobic methanotrophic organisms (especially those affiliated with the genera Methylomonas) were the only organisms found, which are capable of oxidizing methane, accounting for nearly 5% of the relative abundance, while there was a decay of initially abundant organisms. Various heterotrophic groups were also identified, indicating that methanotrophs allowed establishment of a diverse community, even under adverse environmental conditions. These results suggest aerobic methanotrophs are versatile under anoxic conditions and that it is a widespread phenomenon, since it was observed for both inocula tested.
Introduction
M
Aerobic methanotrophs are incapable of oxidizing carbon–carbon bounds (Hanson and Hanson, 1996) and are equipped with methane monooxygenase (MMO), an enzyme that catalyzes the oxidation of methane to methanol. The MMO is evolutionary related to ammonia monooxygenase (AMO), which catalyses ammonia oxidation in nitrifiers (Bédard and Knowles, 1989). The homology between MMO and AMO is possibly responsible for the versatility in nitrogen metabolism reported for aerobic methanotrophs (Stein, Klotz, 2011). Aerobic methanotrophs are capable of assimilating NH4+, NO3−, and N2 as nitrogen source (Bodelier and Laanbroek, 2004), and reduce NO3− through a dissimilatory pathway (Hoefman et al., 2014).
Despite these possibilities, aerobic methane oxidation coupled to denitrification (AME-D) usually occurs in syntrophy, with aerobic methane oxidizers converting methane into an organic intermediate (such as methanol), which is then used as an electron donor by the denitrifiers (Knowles, 2005). Indeed, aerobic methanotrophs have been reported to coexist with other non-methylotrophic chemoorganotrophs (Oshkin et al., 2015). Although this coexistence is usually related to carbon cross-feeding, researchers recently described a mechanism that allows certain aerobic methanotrophs to ferment under oxygen-limited conditions, producing acetate, succinate, lactate, and hydrogen (Kalyuhznaya et al., 2013). Hence, aerobic methanotrophs could sustain a community with various metabolic capacities, including denitrification.
On the other hand, denitrifying anaerobic methane oxidation (DAMO) process is related to occur in two pathways. In one of them, aerobic oxidation of methane by intracellular oxygen provided by the reduction of nitrite to N2 (Ettwig et al., 2010). “Candidatus Methylomirabilis oxyfera sp.,” which belongs to the phylum “NC10,” uses this pathway. Although M. oxyfera sp. was the only isolated microorganism belonging to NC10 phylum so far, other species belonging to this same candidate phylum are thought to be capable of the same metabolism (He et al., 2016). Anaerobic methane oxidation coupled to denitrification can be carried out by an anaerobic methanotrophic archaea (“Candidatus Methanoperedens nitroreducens”) (Haroon et al., 2013). These microorganisms are capable of performing full reversal methanogenesis, using nitrate as the terminal electron acceptor (Haroon et al., 2013) and converting it to nitrite.
Methanotrophic denitrification is a prominent alternative because anaerobic reactors generate methane. UASB effluents are usually poor in organic compounds that could be used for conventional denitrification, requiring the addition of an external electron donor or the use of an alternative electron donor, such as methane. The latter dismiss the need of adding organic compounds (such as ethanol), hence diminishing operational costs. It is especially interesting considering that UASB effluents are usually supersaturated with dissolved methane, presenting up to 25 mg CH4 L−1 (Pauss et al., 1990; Hartley and Lant, 2006; Souza et al., 2011).
Several factors limit the applicability of methanotrophic denitrification despite its potential for wastewater treatment. A low specific denitrification activity has been identified as an issue affecting the applicability of methanotrophic denitrification (Cai et al., 2015; Zhu et al., 2016). The typical slow growth rates and long enrichment times have also hindered technological applications of DAMO processes. Issues in establishing the processes have been linked to missing growth factors, possible product inhibition, and biomass washout, which are problems that can retard the enrichment process (Kampman et al., 2014).
Some authors tried to address the slow growth, biomass washout, and low specific denitrification activity by employing greater solid retention times. The primary strategy has been the application of membrane bioreactors (MBRs) (Sun et al., 2013; Kampman et al., 2014). However, MBR implementation can be costly due to the membranes and energy consumption demands. Another disadvantage is related to fouling problems, which increase operational costs (Skouteris et al., 2012; Lin et al., 2013). To overcome such problems and the membrane cost, alternative methods to enhance biomass retention should be studied. The use of polyurethane foam matrices as a suitable support material for biomass immobilization has been reported since the 80s (Huysman et al., 1983). Since then, polyurethane foam matrices have been successfully tested as a means of biomass immobilization in several biological processes and with various reactor configurations, such as sewage treatment in a sequencing batch reactor (SBR) (Sarti et al., 2006), autotrophic denitrification in a vertical fixed bed reactor (Moraes et al., 2012), and formaldehyde degradation in a horizontal fixed bed reactor (Oliveira et al., 2004). However, polyurethane foam matrices have never been tested as a support material for methanotrophic based processes.
The source of the inoculum is another important feature of bioprocesses (De Vrieze et al., 2015). Various inocula have already been tested for the enrichment of methanotrophs. AME-D was achieved using activated sludge as inoculum (Modin et al., 2008) and in soils and sediments with an oxic–anoxic interface (Bodelier and Laanbroek, 2004). Successful enrichment of a culture containing the phylum NC10 was accomplished using ditch sediment (Raghoebarsing et al., 2006; Ettwig et al., 2009; Kampman et al., 2012), anaerobic reactor sludge (Luesken et al., 2011), or freshwater sediment and rice paddy soil (He et al., 2015a). Rice paddy soil is rich in methane and has been shown to be a good inoculum for the enrichment of methanotrophic denitrifiers. Anaerobic methanotrophic archaea (DAMO archaea) have been obtained from a mixed inoculum that contained anaerobic digester sludge, freshwater lake sediment, and return activated sludge from a sewage treatment plant (Hu et al., 2009; Haroon et al., 2013). Thus, the literature shows that methanotrophic bacteria can be found in many different environments, especially in methane-rich ones, such as anaerobic reactors and mangrove areas.
Bioreactor configuration also plays a role in enriching slow growers. Sequence batch reactors are considered an adequate tool to enhance biomass retention (Strous et al., 1998). This configuration has been previously used to enable methane oxidation coupled to denitrification process (Ettwig et al., 2009; Hu et al., 2011; Luesken et al., 2011; Haroon et al., 2013).
In this study, SBRs were used to evaluate the methanotrophic denitrification, immobilizing the biomass with polyurethane foam matrices. Two different inoculum types were used: one from an anaerobic environment (a UASB sludge used to treat poultry slaughterhouse wastewater) and the other from a methane-rich environment (a blend of marine and mangrove sediments).
Materials and Methods
SBRs and experimental procedure
SBRs were operated for 186 days and two inocula were tested. The inocula were marine sediment mixed with mangrove sediment (Sed inoculum) or anaerobic granular sludge collected from a UASB reactor used to treat poultry slaughterhouse wastewater (AnS inoculum). Each inoculum was tested in triplicate (the reactors were designated as Sed-SBR and AnS-SBR).
Methane was provided as the sole electron donor and nitrate and nitrite were supplied in concentrations compatible to those found in nitrified effluent from sewage treatment plant. The SBRs were borosilicate glass flasks with a volume of 1,129 ± 12 mL. The reaction volume was ∼700 mL, and the headspace volume was 430 mL.
Two control reactors were also maintained, one for each inoculum type (Sed control and AnS control reactors). The purpose of the control reactors was to measure the endogenous consumption of oxidized forms of nitrogen. Therefore, nitrate and nitrite were added, but no electron donor was provided.
Batch cycles took 3 days each (72 h), and the liquid medium was completely changed at the end of each cycle after being purged with the aid of a siphon and filling with freshly prepared medium. The liquid medium change procedures took around 15 min (0.25 h). The reactors were then flushed with argon for 15 min (0.25 h), followed by methane for 3 min (0.05 h). Sed-control and AnS-control were only flushed with argon for 15 min. Gas flow was kept at 2.5 L min−1 with the aid of a hydraulic gas flow meter (RWR) equipped with a valve. All the SBRs, including methane-free controls, were kept under an initial relative pressure of 230 ± 25 mBar. Stainless steel porous stones, similar to those used in liquid chromatography, were used for gas flushing. During the cycle operation, the reactors were maintained at a controlled temperature (30 ± 2°C) in a dark chamber and agitated at 130 rpm (Nova Etica, model 430).
The initial methane concentration was measured by gas chromatography (GC) (Perna et al., 2013), and dissolved methane was calculated according to the Henry's Law. We used the Henry's Law coefficient for 30°C (Sander, 1999). Hence, initial methane concentration was 20.6 ± 7.7 mg CH4 L−1, which is the range of methane supersaturated UASB effluent (Souza et al., 2011).
An abiotic control, designated as “Blank,” was also maintained throughout the experiment, to check for gas leaking through the rubber stopper. A borosilicate glass flask was fulfilled with tap water maintaining the same headspace volume adopted for the SBR. The Blank control was flushed with argon and methane following the same procedures previously described. The pressure was monitored with a pressure measurement device from Desin Instruments (TPR18 model).
Inocula characterization and medium composition
Biomass was fixed on polyurethane foam matrix cubes with a 12 mm edge, wrapped by a PVC spiral structure with 12 mm in diameter and in height. One hundred units of support material (a total mass of 39.065 ± 1.566 g) were used in each reactor. Granular sludge was ground in a blender before inoculation. For immobilization procedures, the polyurethane foam cubes were immersed in the blend of marine and mangrove sediments or in the ground anaerobic sludge for 12 h (Zaiat et al., 1994).
Total solids (TS) and total volatile solids (TVS) content of both inocula were determined according to APHA (2005) (method 2540). The Sed inoculum contained 0.537 ± 0.030 g TS.g inoculum−1 and 0.015 ± 0.001 g TVS.g inoculum−1. The AnS inoculum contained 0.054 ± 0.002 g TS.g inoculum−1 and 0.036 ± 0.004 g TVS.g inoculum−1. By the end of experiment, the TS and TVS were analyzed. To analyze solids content in the end of experiment, solids were removed from the polyurethane foam with the aid of a glass rod and distilled water.
Composition of mineral medium was adapted from van de Graaf et al. (1996) and included 34 mg.L−1 NaNO2; 42 mg.L−1 NaNO3; 27.2 mg.L−1 KH2PO4; 300 mg.L−1 MgSO4.7H2O; 180 mg.L−1 CaCl2.2H2O; 1 mL.L−1 trace elements solution I; and 1 mL−1 trace elements solution II. The trace element solution I contained 5 g.L−1 EDTA and 5 g.L−1 FeSO4. The trace element solution II contained 15 g.L−1 EDTA, 0.43 g.L−1 ZnSO4.7H2O; 0.24 g.L−1 CoCl2.6H2O; 0.99 g.L−1 MnCl2.4H2O; 0.25 g.L−1 CuSO4.5H2O; 0.22 g.L−1 NaMoO4.2H2O, 0.19 g.L−1 NiCl2.6H2O; 0.09 g.L−1 Na2SeO3.5H2O; and 0.014 g.L−1 H3BO4.
Temporal profiles
After 115 days of enrichment, the polyurethane foam matrices from one of the triplicates were divided into four smaller batches to carry temporal profiles assay. Four conditions were applied to each biomass types: (1) nitrite and methane supplied; (2) nitrite supplied, but no electron donor provided; (3) nitrate and methane supplied; and (4) nitrate supplied, but no electron donor provided. Hence, four batch reactors evaluated the temporal profiles for the biomass from Sed-SBR, (Sed-TP, tested under the aforementioned conditions I–IV); and the other four reactors evaluated the profiles for the biomass from AnS-SBR (AnS-TP) (Fig. 1).

Experimental setup for temporal profiles assay. SBR, sequencing batch reactor.
Temporal profile assays were conducted in batch reactors, which were borosilicate glass flasks with an effective volume of 607 ± 7 mL. Twenty-five support material units from the enriched SBRs were used, which accounted for 10.54 ± 0.06 g of the support material (0.045 ± 0.01 g TVS.g support material−1) for reactors that used the biomass from the Sed-SBR. For the batch reactors using enriched biomass from AnS-SBR, the support material mass was equivalent to 10.26 ± 0.12 g (0.083 ± 0.013 g TVS.g support material−1). In all the four conditions, the nutritional medium was the same described in Inocula Characterization and Medium Composition section. For conditions I and III, methane was added, while no electron donor was provided for conditions II and IV. The assays lasted until nitrate or nitrite concentrations were below the detection limit of the analytical method or the concentration stabilized. The sampling interval changed throughout the experiment: 1 mL samples were collected hourly during the first 6 h, and the sampling was performed every 6 h from 6 to 36 h. After 36 h, samples were collected every 12 h until the end of the experiment.
Analytical methods
SBRs were monitored at the beginning and end of each operation cycle. The monitoring analysis included nitrite and nitrate concentrations, determined using an ionic chromatograph Dionex ICS-5000 equipped with an ion suppressor and conductivity detector. The column was maintained at 35°C, and isocrat elution was used. The eluent was composed of a solution of sodium carbonate (4.5 mM) and sodium bicarbonate (0.5 mM).
Samples were collected at various times during the batch cycles to assess organic compounds that could eventually be formed as intermediates of methane oxidation. The samples were analyzed by gas and high performance liquid chromatography (GC and HPLC, respectively). The former followed the methods described by Adorno et al. (2014) using a flame ionization detector and was used to detect methanol, ethanol, and various volatile organic acids. The HPLC method was described by Penteado et al. (2013). Two detectors were operated in series—a UV detector and a refraction index detector. The HPLC method was used to determine methanol, ethanol, n-butanol, organic acids, glucose, and fructose.
Methane consumption could not be measured because methane was provided in excess and the conversion remained within the confidence interval of the method.
Nonparametric statistical tests
Nonparametric tests were applied to analyze results. Mann–Whitney test was used to compare two independent samples, while Wilcoxon test was applied for two paired samples. The Kruskal–Wallis test was used to analyze three independent samples. In all tests, the adopted confidence level was 95%.
Biological analysis by 16S rRNA genes using IonTorrent
Samples were taken from the inocula (Sed-inoculum and AnS-inoculum) and the biomass in the reactors after 165 days of operation (Sed-SBR and AnS-SBR). Samples were successively washed thrice in phosphate-buffered solution (PBS 1X) and subsequently centrifuged at 6,000 rpm for 10 min. Total DNA was extracted with the phenol:chloroform protocol (Griffiths et al., 2000). The amount of DNA was checked with an ND-2000 spectrophotometer (Nanodrop, Inc., Wilmington, DE).
Sequencing with the Ion Torrent platform was performed in chip 318 V2 400 bp. The primers used for amplifying the V4 region of the 16S rRNA were 577F (5′-AYTGGGYDTAAAGNG-3′) and 924R (5′-CCGTCAATTCMTTTRAGT-3′) (Cole et al., 2005).
Sequences were processed using Quantitative Insights Into Microbial Ecology (QIIME) (Caporaso et al., 2010). Chimeras were filtered using the UCHIME algorithm (Edgar et al., 2011). Reads with lengths <200 bases or with expected error >0.5 were removed. Operational taxonomic units (OTUs) were determined using the UCLUST method at a 97% similarity level (Edgar, 2010). The OTU tables were normalized using a QIIME random subsampling (Caporaso et al., 2010). The singletons were removed for taxonomic classification, which was aligned to the Greengenes database v 13.8 with a confidence threshold of 50% (De Santis et al., 2006).
Sequences were submitted to the European Nucleotide Archive (www.ebi.ac.uk) under the accession numbers ERS1201616 to ERS1201619 (project accession number PRJEB14367).
Results and Discussion
Performance of SBRs for denitrification and cell maintenance
The results suggest that methane oxidation supported nitrate and nitrite consumption. The total nitrogen removal efficiency achieved was 49.9 ± 27.0% (Sed-SBR) and 86.1 ± 11.5% (AnS-SBR). Meanwhile, the efficiency removal was 10.4 ± 10.4% in the Sed control and 18.3 ± 18.3% in the AnS control. A Mann–Whitney test (95% confidence level) revealed that the difference between nitrogen consumption data from the SBR containing methane and the respective methane-free control was statistically significant, indicating that methane oxidation supported biological nitrogen consumption (W: 226.5, p: 1.05 × 10−6 to Sed and W: 927, p: 4.0 × 10−6 to AnS). A Kruskal–Wallis test showed that the SBRs operated in triplicate with the same inoculum under the same conditions showed no statistically significant differences in nitrogen consumption (χ2: −216; p: 1 for Sed-SBR and χ2: −192; p: 1 for AnS-SBR). This indicates the methane oxidation-based denitrification was consistent over the triplicates. Moreover, neither Sed-SBR nor AnS-SBR had any preference between the tested nitrogen forms (NO2− and NO3−), according to the Wilcoxon test (V: 298, p: 0.588 and V: 171, p: 0.473, respectively). These data suggest that, regarding total nitrogen removal efficiency, methane oxidation coupled to denitrification in immobilized biomass in polyurethane foams might be feasible for wastewater treatment. However, more studies are needed to optimize the process, since the denitrification rates were kept low throughout the experiment.
Average nitrogen removal rate (i.e., the sum of NO2
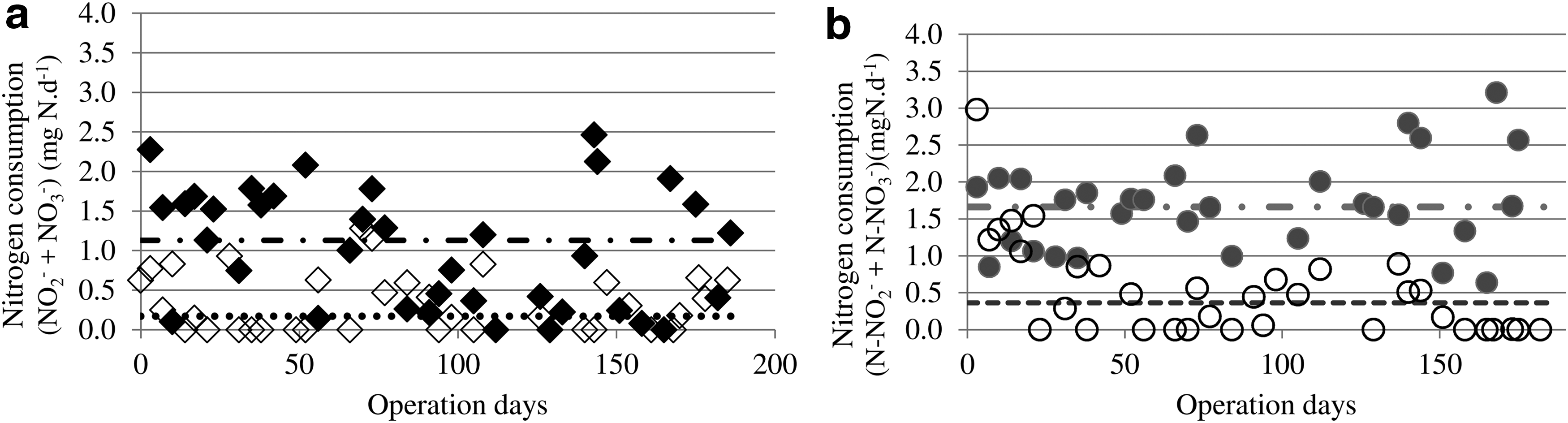
Nitrogen consumption over 186 days of SBR operation.
Similarity of denitrification activity found in this study and those conducted with suspended biomass (Gavazza dos Santos et al., 2004; He et al., 2015b) could indicate that polyurethane foams are not suitable media for immobilization of methanotrophic organisms. Another hypothesis is that biomass washout is not the only factor hindering the application of methanotrophic denitrification processes. Most likely, there are other factors that influence the establishment of the process, such as specific growth factors.
Methane not only supported denitrification but also allowed cell maintenance in the SBRs inoculated with anaerobic sludge (AnS-SBR). At the beginning of the experiment, the total volatile solids content in AnS-SBR was 2.844 ± 0.129 g TVS (0.073 ± 0.002 g TVS.g support material
In addition, the nitrogen consumption in AnS-SBR was statistically higher than in Sed-SBR (Fig. 3a). The Mann–Whitney test indicated that the difference was statistically significant (W: 838, p: 0.0012). However, this difference could be related to the volatile solids content. The specific nitrogen consumption (Fig. 3b) was 0.623 ± 0.373 mg N.g TVS

Boxplot of
Temporal profiles
The temporal profiles of nitrate and nitrite consumption were similar for both inocula (Fig. 4). Nitrite and nitrate reduction were observed to a lesser extent under methane-free conditions (conditions II and IV), despite the fact that the same enriched biomass was used for all the four conditions tested. This observation hints on favor of previous remarks—that denitrification only occurs in the presence of methane.
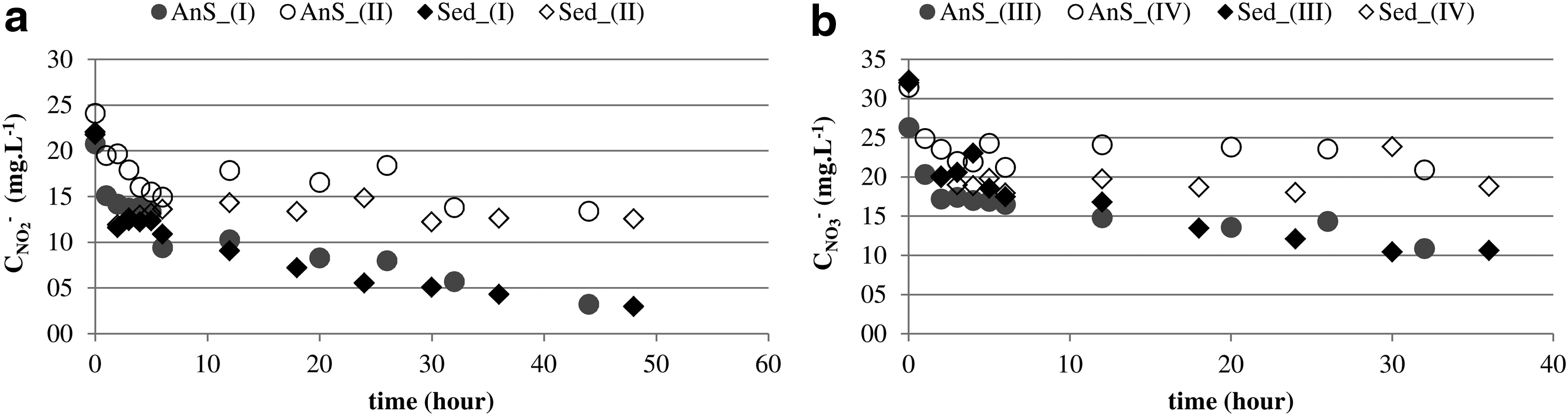
Temporal profiles for marine and mangrove sediments (Sed-TP) and anaerobic sludge (AnS-TP) after 115 days of enrichment providing methane as sole electron donor and carbon source.
Temporal profiles showed that the maximum denitrification activity was 8.9 mg NO2
Temporal profile suggests that the apparent denitrification kinetic in this configuration resembles a first order kinetics. Thus, higher initial nitrogen concentrations could enhance the denitrification activity. Indeed, domestic sewage pretreated in UASB reactors contains low nitrogen concentrations, which could have impaired the establishment of methanotrophic-denitrifier organisms. Nevertheless, the obtained temporal profiles indicate that higher denitrification rates can be achieved by manipulating operational conditions.
Bacterial community
Composition of bacterial community was assessed by 16S rRNA massive sequencing by IonTorrent®. Data after normalization procedures are presented in Table 1. The Sed inoculum showed the highest richness, which the higher Chao1 and Shannon hints. This was expected, since the Sed inoculum was obtained from soil in a sulfate-rich environment. It was also expected that microorganisms related to the sulfate cycle appear among the most abundant (Fig. 5). For AnS inoculum, the most abundant OTUs are chemoorganotrophs capable of oxidizing organic compounds under anaerobic conditions.
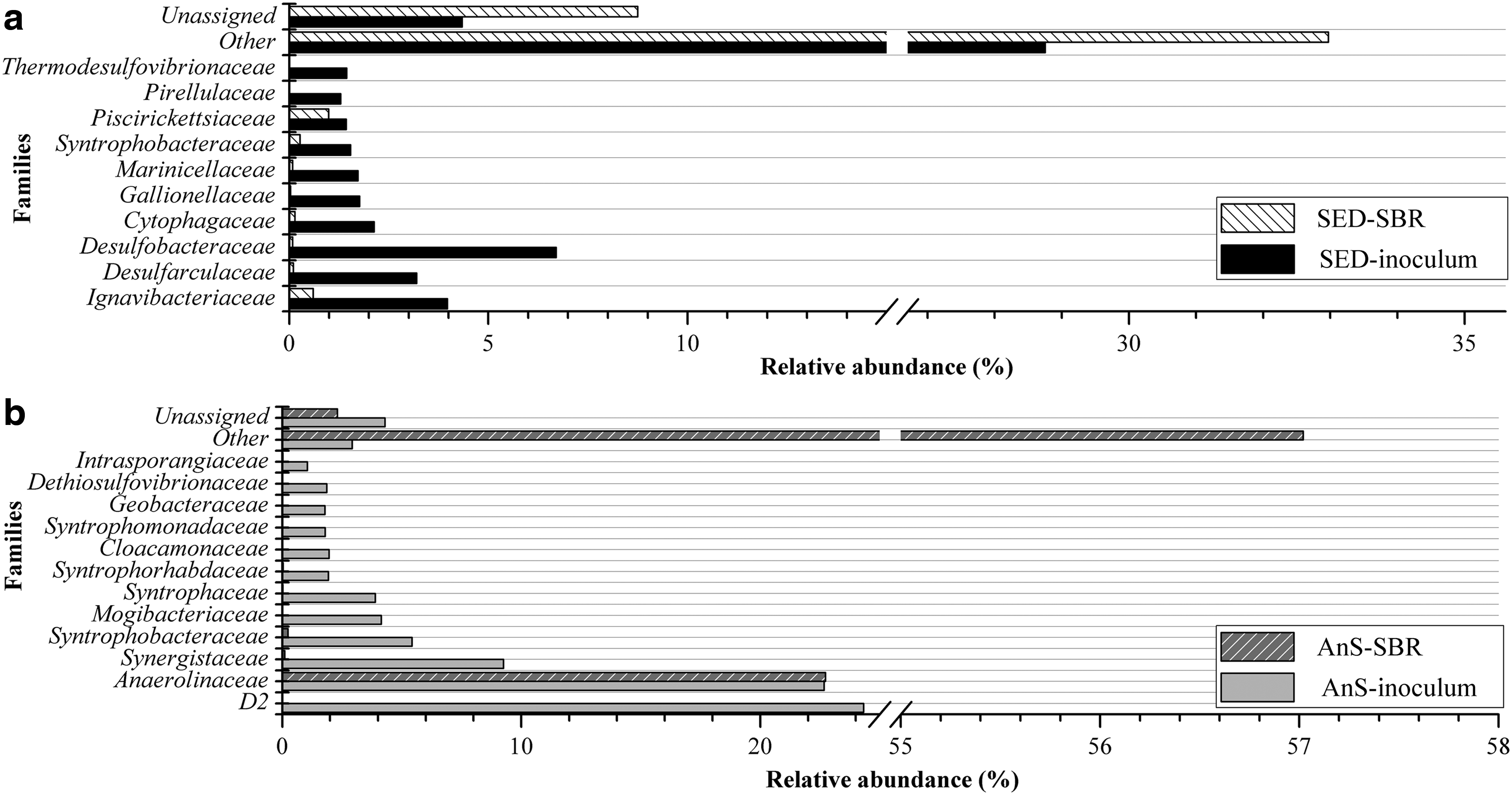
Most abundant families in
OTUs, operational taxonomic units.
Sequencing data show that aerobic methanotrophs were favored in both enriched inocula (Sed-SBR and AnS-SBR) and supported a heterotrophic community (Figs. 5 and 6). OTUs related to aerobic methanotrophs had a relative abundance of 0.09% in the Sed inoculum and were responsible for 6.85% in the enriched sample (Sed-SBR). Methanotrophic-related OTUs were not identified in the AnS inoculum, but accounted for 6.99% in the enriched sample (AnS-SBR). Thus, aerobic methanotrophs were negligible in both inocula and flourished due to the SBR operation procedures.
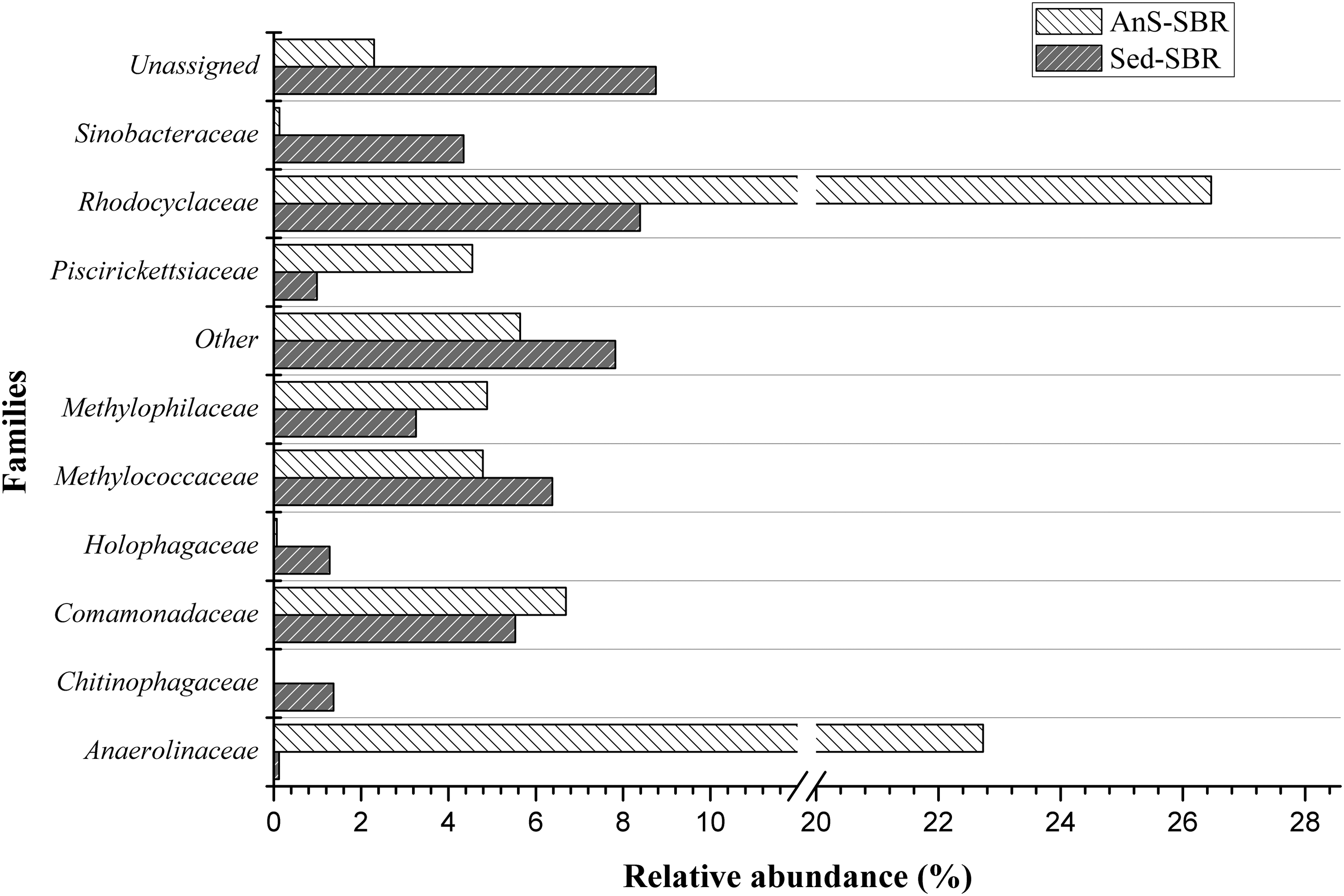
Most abundant families in enriched samples Sed-SBR and AnS-SBR. OTUs that could not be related to any known taxonomic classification are referred as Unassigned. OTUs, the relative abundance of which was <1.0% for both samples, are referred as “Others.” OTUs that were not identified to the family level are not shown.
Among the aerobic methanotrophs identified in Sed-SBR and AnS-SBR, those affiliated with the genus Methylomonas (which belongs to the family Methylococcaceae) were more abundant and its establishment is in accord with the SBR monitoring data. The genus Methylomonas was more abundant than other methanotrophs and accounted for 5.70% in Sed-SBR and 4.73% in AnS-SBR. All species of Methylomonas described thus far can use NO3− as a nitrogen source for assimilation (Brenner et al., 2005) and have a relatively high nitrogen requirement (Bodelier and Laanbroek, 2004). The observed nitrogen consumption could be due to assimilatory functions, which is supported by the solids content data in the reactors.
Organisms belonging to the genus Methylomonas have been reported to be strictly aerobic. However, they should have played an important role under the anoxic conditions maintained in the SBRs. It should be emphasized that the SBRs were flushed with argon for 15 min, whenever the flasks were opened to change the medium. The only period of time when the microorganisms could have been exposed to oxygen was during the change of the medium. This procedure took an average of 15 min (0.25 h), which corresponds to 0.46% of the total time of the batch cycle (72 h). Authors who worked with methane oxidation under microoxic conditions usually continuously provide oxygen to prevent anoxia (Liu et al., 2014), which implies aerobic methanotrophs quickly consume oxygen whenever it is present. Furthermore, the Blank control results show that there was no gas leaking throughout the batch cycle, since its relative pressure was kept at 222 ± 10 mBar. Thus, the presence of Methylomonas under the conditions of this study indicates the possibility of a versatile metabolic pathway or syntrophic relationship that could support it.
Aerobic methanotrophic bacteria were already reported under anoxic conditions, but were thought to be barely active (Roslev and King, 1995; Schubert et al., 2006; Cuba et al., 2011). However, recent studies have suggested that methanotrophic organisms are able to ferment under oxygen-limiting conditions and excrete products to other non-methanotrophs (Kalyuhznaya et al., 2013). This indicates that aerobic methanotrophs are more versatile than previously thought and the role of aerobic methanotrophs under anoxic and oxygen-limiting conditions remains not fully understood. The results obtained in our study seem to support this idea.
Aerobic methanotrophs were the only organisms identified, which can oxidize methane. DAMO bacteria were not identified, despite the fact SBR operation procedures were expected to favor them better than aerobic methanotrophs. DAMO organisms were also not detected in inocula samples. As previously discussed, cell maintenance was observed for both inocula, but biomass went extinguished in methane-free controls. Nevertheless, aerobic methanotrophs were not the most abundant in enriched samples Sed-SBR and AnS-SBR (Fig. 6). Interestingly, among the most abundant families, there are mostly chemoorganotrophs. Since methane was the only external carbon source provided, other heterotrophic organisms depended on methanotroph metabolism to establish.
These findings indicate that methane as the sole external electron donor and carbon source can support other heterotrophic bacteria, implying the establishment of a syntrophic relationship. Other researchers have already reported syntrophic relationships between methanotrophic organisms and non-methylotrophic chemoorganotrophs. Kalyuhznaya et al. (1999) have already claimed that methanotrophs provide available carbon sources for other members of the microbial community. Oshkin et al. (2015) argue that community dynamics are more important in methane cycling than a single organism or even the size of the methanotrophic community. A recent understanding also claims that structural organization of microbial communities is more important than identifying the most abundant species, since these are not necessarily the most metabolically active (De Vrieze and Verstraete, 2016). The findings of this study are consistent with the arguments presented by Oshkin et al. (2015) and De Vrieze and Verstraete (2016). Thus, a deep understanding of these syntrophic relationships could contribute to a full comprehension of the technological potential of methanotrophic based technologies.
Among the chemoorganotrophs identified, non-methanotrophic methylotrophic organisms should be mentioned. Members affiliated with the Methylophilaceae family seem to have been favored, especially in the Sed-SBR sample. Reads affiliated with the family Methylophilaceae accounted for 4.26% of Sed-SBR and 1.91% of AnS-SBR, and accounted for <0.02% of inocula samples. Members of the family Methylophilaceae have also been described as strictly aerobic and have been previously found in methanotrophic environments. Methylophilaceae has also been reported to participate in a syntrophic relationship in a denitrifying micro-aerated methanotrophic reactor (Liu et al., 2014). Indeed, Methylomonas and Methylophilaceae have been reported as abundant under oxygen-limited conditions, while methane was provided in excess (Hernandez et al., 2015; Oshkin et al., 2015). Moreover, family Methylophilaceae contains some species that are only able to assimilate methanol under denitrifying conditions (Kalyuhznaya et al., 2009).
As previously cited, non-methylotrophic chemoorganotrophs were also favored This could explain why organic intermediates could not be measured by either the GC or HPLC analysis. Any organic intermediate excreted by methanotrophs was probably consumed by other chemoorganotrophs. A chemoorganotroph usually described in anoxic methanotrophic environments was found among the most abundant for all samples. Members affiliated with the class Anaerolinea, in which is described the family Anaerolinaceae, were identified in the enriched samples with a relative abundance of 16.1% in the enriched Sed-SBR and 24.9% in the AnS-SBR samples. Yamada et al. (2006) described this family as strictly anaerobic chemoorganotrophs incapable of using NO3− (Yamada et al., 2006). Organisms of the class Anaerolineae were found to be abundant in a sample from an anoxic methane oxidation area in the Indian Ocean (Blazejak and Schippers, 2010). It seems that organisms affiliated to the class Anaerolineae are capable of establishing a syntrophic relationship with methanotrophic organisms. Another abundant group identified in Sed-SBR, which was not identified to the family level, was members affiliated with the phylum OP11 (15.3%). This group is commonly identified in anaerobic environments, although its metabolism remains unclear (Wrighton et al., 2012). For both enriched samples, the most abundant organisms are chemoorganotrophs, usually described as anaerobic, which must have survived using products excreted by the methanotrophs.
Although some taxonomic groups usually related to denitrification were identified, the nitrate and nitrite consumption rates were low, as previously discussed. Denitrification is a metabolic pathway that is widely dispersed among prokaryotes. Many members of the families Rhodocyclaceae and Comamonadaceae can denitrify with nitrate or nitrite even under autotrophic conditions (Vilar-Sanz et al., 2013). Members of the family Rhodocyclaceae accounted for 4.64% in the Sed-SBR and 21.07% in the AnS-SBR. Total reads of 3.76% in Sed-SBR and 6.09% in AnS-SBR belonged to the family Comamonadaceae. Considering the low denitrification rates, organisms in these groups were probably maintained by metabolic pathways other than denitrification.
It is also interesting that enriched samples shared more OTUs between them than with their respective inoculum. Figure 4 shows that the most abundant OTUs in both inocula had mostly disappeared in enriched samples, except for the OTUs affiliated with Anaerolinaceae in AnS samples. Moreover, a comparative analysis of the microbial communities had a higher similarity index between the enriched samples. In the UPGMA dendogram, based on the Bray–Curtis similarity index (Fig. 7), enriched samples were clustered together in 93% of runs. The Bray–Curtis index was 25% between the enriched samples, whereas it was ∼16% between the enriched biomasses and their respective inocula. Considering the different inoculum sources, one with a high richness, the increment of the similarity index between the enriched samples indicated that the enrichment procedure could select specific microorganism groups found in the different inoculum sources. It is noteworthy that several of the most abundant groups found in the samples from the AnS-SBR and Sed-SBR had similar capabilities such as those belonging to the families Methylococcaceae, Methylophilaceae, and Anaerolinaceae (Fig. 6).

UPGMA dendrogram based on Bray–Curtis similarity among inocula and enriched samples. Bray–Curtis similarity was calculated after sample normalization procedures. Bootstrap values shown as percentage (1,000 runs).
SBRs operation procedures allowed the establishment of methanotrophic, methylotrophic, and other chemoorganotrophs related to the syntrophic methane metabolism. These findings indicated that organisms affiliated with genus Methylomonas are highly adaptable and are able to establish even under adverse environmental conditions, such as intermittent oxygen-limited and anoxic conditions. Moreover, they perform an important function in establishing syntrophic dynamics, even when they are not the most abundant organisms in the community.
Conclusions
Methane supported nitrite and nitrate reduction, and nitrogen efficiency removal suggests that methanotrophic denitrification can be applied to nitrified effluents from sewage treatment plants. The low denitrification rates indicate that biomass immobilization in polyurethane foams did not enhance the establishment of an exclusive methanotrophic-denitrifier community. Despite that, the analysis of bacterial community showed aerobic methanotrophs grew under mostly anoxic conditions. Methane was provided as the only carbon source and organisms of the family Methylococcaceae were the only organisms identified that are associated to the direct use of methane. Aerobic methanotrophs allowed the establishment of other heterotrophic groups, hence supporting a diverse bacterial community. These findings suggest that aerobic methanotrophs potentially play a key role in syntrophic dynamics even when submitted to adverse environmental condition, and can thus be technologically explored.
Footnotes
Acknowledgments
The authors thank the Fundação de Amparo à Pesquisa no Estado de São Paulo (FAPESP 2013/08257-8 and 2009/15984-0) for financial support.
Author Disclosure Statement
The authors declare that they do not have any conflict of interests.