
Abstract
Abstract
Algal turf scrubbers (ATSs) have been used for water remediation in many applications, generating benefits such as water quality improvements and biomass production. Their economical operation is hampered, however, by uncontrolled mixed microbial communities, typical of open cultivation systems that result in lower biomass quality. Design of substratum topography is one approach to control the algal community composition by constraining colonization dynamics. This study seeks to elucidate the role of substratum topography on biomass productivity of benthic algae in an ATS environment. A set of tiles with custom-designed textures of repeated hemispheres across a range of diameters were 3D printed and cast on ceramic clay. An algal turf community dominated by Mougeotia taxa was cultivated on the tiles in a laboratory-scale ATS and harvested for biomass measurements after 21 days. Results show higher biomass on the topographies formed of 500 and 2,000 μm diameter hemispheres, with 74.3% more attached biomass than the 1,000 μm hemisphere and smooth topographies. With the surface area, material roughness at the microscale, and surface chemistry discarded as factors by experimental design, surface topography, especially interstitial space distribution, at the millimeter scale is shown to be a factor in the kinetics of algal cell colonization and growth on surfaces. This study is among the first that specifically focuses on the application of substratum design using additive manufacturing to improving the ATS process. These results can have a significant impact on design of next-generation high-productivity attached growth algae systems.
Introduction
A
Full-scale applications are typically open floway reactor systems colonized by the mixed algal communities indigenous to the local water source, and the types and abundance of the algal taxa that colonize, attach, and persist will determine, in part, the overall yield of the particular biotic process or product of interest. Limitations on economic use of low-quality ATS biomass can be mitigated by increased productivity from use of three-dimensional (3D) substrates as preliminarily demonstrated in a previous study (Adey et al., 2013). Overall system performance and economic viability of the ATS may, therefore, be improved by controlling the algal colonization and growth processes.
One approach to improving ATS performance is through intentional 3D design of the substratum surfaces on which algae colonize (Kardel et al., 2015; Carrano et al., 2016; Kardel, 2016). In flow environments, the topographical characteristics of the substratum can be a controlling factor in the development of a colonizing turf (Stevenson, 1996) through interactions with the flow turbulence at the fluid boundary layer, thereby affecting the transport phenomena and kinetics of algal cell colonization and growth (Norton and Fetter, 1981; Burkholder and Wetzel, 1989; Whitehead and Verran, 2006). The characteristics of the flow at the boundary layer also affect advection and dispersion of nutrients to colonizing cells, which can strongly influence growth kinetics (Whitehead and Verran, 2006). Therefore, it is conjectured that manipulation of the substratum topography characteristics can be used to control colonization of algae, and thereby determine the structural and functional characteristics of the algal turf.
There is substantial support for this conjecture, based upon numerous studies involving colonization dynamics of various microorganism types. In situ studies in streams and laboratory investigations have shown that the ecological characteristics of a colonizing benthic algal community are influenced by the physical characteristics of the substratum (Burkholder, 1996). In general, the direct relationship between surface roughness and retention of microbial cells is well recognized because of potential problems in biofouling, infection, and surface hygiene, or cleanability, and common methods exist for measuring surface roughness and visualizing topography (Whitehead and Verran, 2006). Substratum surface topography has been shown to affect the retention of colonizing diatom cells, presumably from induced local boundary layer effects (Miller et al., 1987). This effect is observed, however, for a range of topographic dimensions relative to cell size; surface topographic features with dimensions of the order of cell size have higher cell retention rates than those with features significantly larger or smaller (Whitehead and Verran, 2006).
For any colonizing microbial taxa, there should be a surface feature scale that is optimal for settlement and attachment of colonizing cells. For example, in a study on settling and attachment dynamics of propagules of the macroalga Sargassum muticum, it was observed that attachment directly increased with surface roughness feature size from smooth to an optimum scale of 800 μm, which was on the same order as the size of the propagules (Norton and Fetter, 1981). In studies on periphyton colonization on various substrata materials, it was observed that substrata with heterogeneous surface microrelief, such as the microcrevices found in rocks, offer depressions where colonizing cells can settle without being dislodged by abrasion or shear (Miller et al., 1987; Burkholder and Wetzel, 1989). In addition, various studies show that different surface topographic scales resulted in selective colonization of algal species, presumably through effects on the near-surface shear in the laminar flow sublayer (Harlin and Lindbergh, 1977; Norton and Fetter, 1981; Stevenson, 1983; Reed et al., 1988). Recent work in applying these concepts to engineered systems for algal cultivation have focused on engineered materials and substrate textures that influence biofilm formation from planktonic species (Cao et al., 2009; Cui et al., 2013; Ozkan and Berberoglu, 2013; Gross et al., 2016). Few of these studies, however, have focused on obligate benthic and attached filamentous species of periphytic origin in flowing waters. Recent experiments in natural freshwater flow systems showed definitive and significant effects of surface topography on colonization presence of at least two filamentous algal genera, Microspora and Stigeoclonium, in which each displayed an optimum topographic feature size scale for maximum colonization (Blersch et al., 2017). Results from these studies suggest that taxon selection and control are partially influenced by surface properties in the development of microbial biofilm precursors of algal turfs (Irving and Allen, 2011).
Considering the recent inquiries into algae–substratum relationship dynamics, and the lack of focus on filamentous turf-forming species, the state-of-the-art for development of the ATS process has not currently addressed the design of targeted substratum topographic characteristics for increased algal productivity performance. The objective of this study is to understand the effect of surface topography, in particular, surface feature height and interstitial space distribution, on total colonization and biomass productivity of an algal turf community in repeatedly harvested, laboratory-scale ATSs. Three dimensional printing was used to produce molds for growth tiles with a variety of surface topography treatments in which both feature height and the number of interstitial spaces were varied and later exposed to cultivation conditions in the laboratory.
Materials and Methods
For the experiments, we designed and fabricated a pattern plate containing four sections with different surface topographies using a polyjet-based 3D printer. This plate was used as a template mold to produce identical growth tiles out of unglazed ceramic clay. The clay tiles were then placed in a laboratory scale ATS reactor seeded with an algal community dominated by attached filamentous algae. After 21 days of cultivation, the attached algal biomass from each section was harvested, oven-dried, and weighed. Statistical analyses were used to assess the effect of surface topography on algal biomass.
Substratum design and fabrication
The pattern plate was designed in Solidworks® (Dassault Systemes Solidworks Corp., Waltham, MA) and fabricated with an acrylate-based resin using an Objet30 polyjet 3D-printer (Stratasys Ltd., Eden Prairie, MN) with a 28 μm layer thickness. A square plate (105 × 105 mm) was designed comprising four equal sections (52 × 52 mm each) containing hemispherical surface features of increasing scale. One of the sections was designed as ideally smooth (area average roughness Sa = 1.19 μm, as measured with a profilometer), whereas the remaining three sections had a pattern of square-packed adjacent hemispheres of average area peak-to-valley distance (Rz) of 500 μm, 1,000 μm, and 2,000 μm. Figure 1 shows the computer model with the corresponding 3D-printed polymer pattern plate and clay growth tile.

Plan view of computer model, 3D-printed pattern plate, and clay growth tile.
To assess the dimensional accuracy of the fabrication, the surface of the plates was scanned with an axial chromatic noncontact profilometer (Nanovea ST-400, Irvine, CA) with 20 nm resolution in the vertical direction. The dimensional error between the computer model and the 3D-printed plate was <1% for all the sections. Figure 2 shows the 3D areal scan view and a 2D line scan of the surface topography of the 500 μm section of the pattern plate.
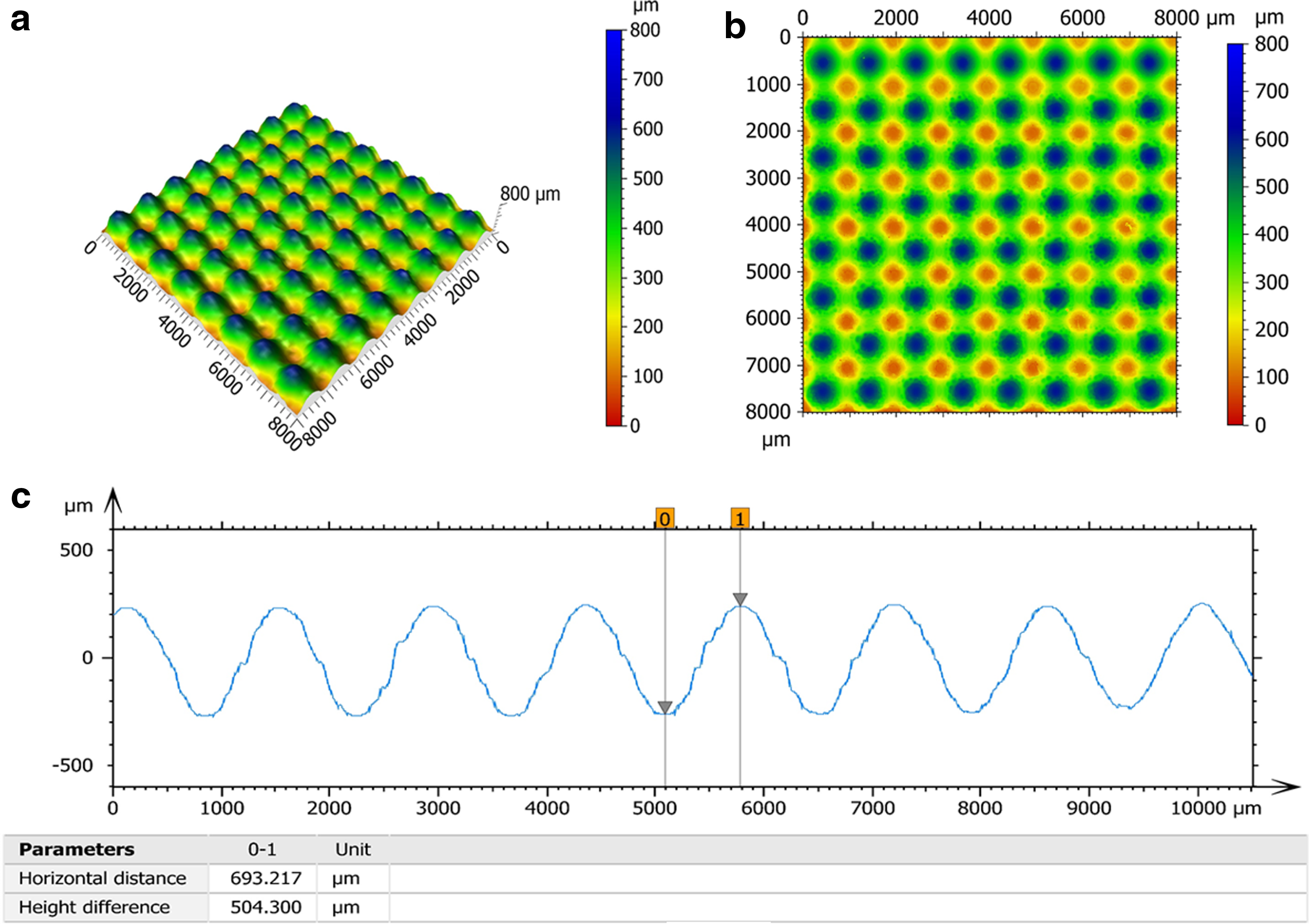
Topographical maps of 500 μm section:
The 3D-printed pattern plate was then pressed in clay material to produce a growth surface with high fidelity of the designed surface topography. The clay formulation was typical ceramic clay with constrained porosity and containing the following materials: tile No. 6 clay (40%), Hawthorne fire clay (25%), OM4 ball clay (10%), Custer feldspar (15%), silica (7%), and talc (3%). The molds were made by slurry hand pressing and, from those, 24 tiles were cast and fired in the kilns. The clay firing cycle lasted 12 h per batch with a subsequent cooling cycle of 15 h duration, resulting in hardened unglazed surfaces of various roughness textures (Fig. 1). A total of 24 plates with identical clay formulation and topography pattern were manufactured.
Cultivation experiments
The experiment consisted of three experimental runs (replicates) with eight growth tiles each placed in a laboratory scale ATS reactor. The tiles were fixed to a polypropylene screen in two rows with four tiles each using marine-grade silicone sealant. The tiles in each row were each rotated 90° so that each row contained all four tile orientations facing the incoming water flow (Fig. 3). The randomization of tile rotation continued throughout all three replications.
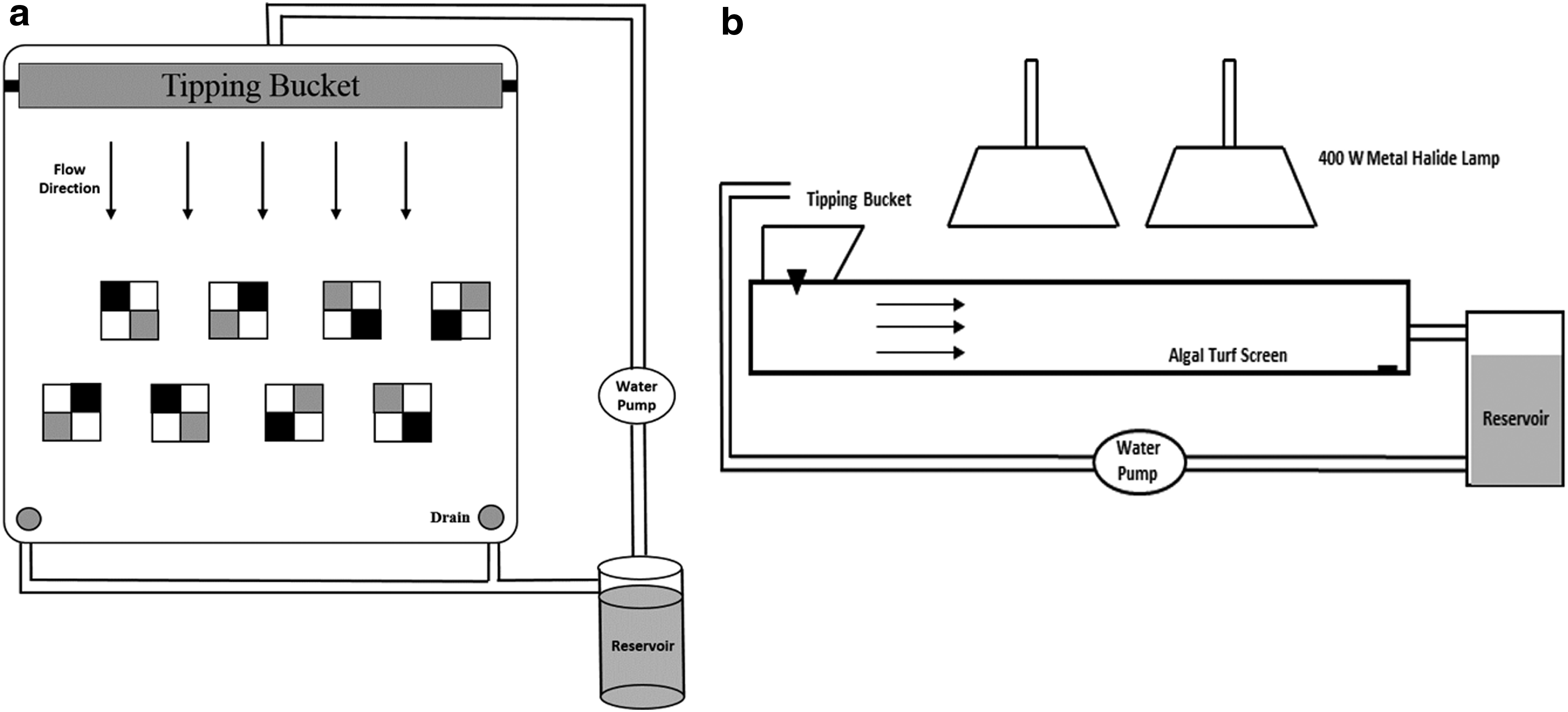
Schematic of algal turf scrubber reactor and tile layout:
The ATS used in this study was a shallow trough with a 1 m2 growth area of 3 mm mesh polypropylene screen substratum (XV-1672; Industrial Netting, Minneapolis, MN) on which algae would attach. Water flowed constantly in the trough, with periodic wave action induced by a tipping wave-surge bucket at an average depth of 2 cm. The water was recirculated from a 120-L collection reservoir through a submersible pump at a flow rate of 45 L/min with a wave surge frequency of 4 min−1. The light was provided continuously by two 400 W metal halide grow lamps (Virtual Sun, La Verne, CA) located directly above the centerline of the cultivator. The relative location and vertical distance of the lamps to the tile surface were adjusted to ensure adequate and homogeneous illumination throughout the growth surface area. An illumination map, measured with a quantum flux meter (MQ-200; Apogee Instruments, Logan, UT) at the substratum surface, shows a relatively uniform light intensity within the experimental region, with an areal average of 245 μmol/(m2·s) (Fig. 4).
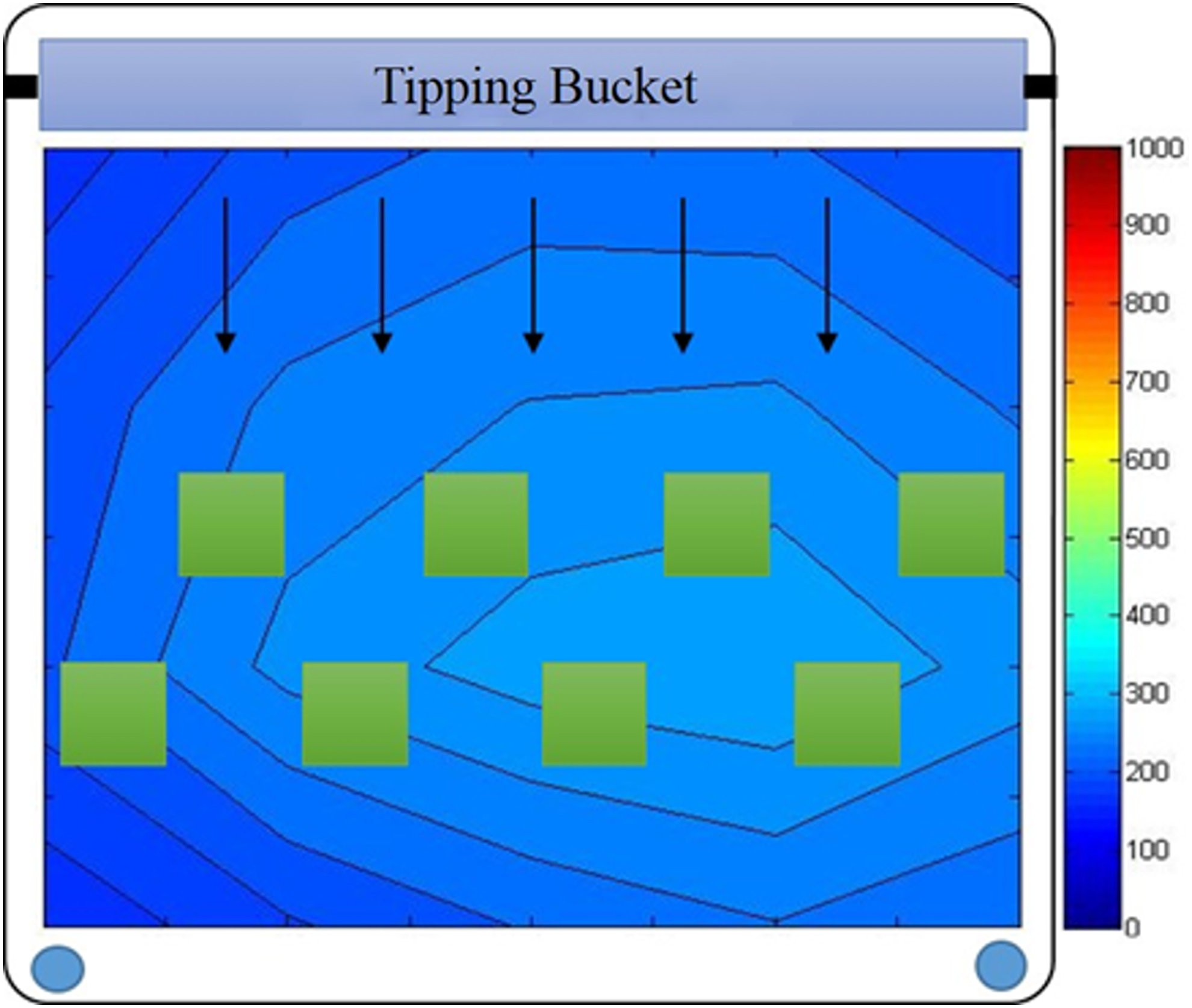
Illumination map for ATS (in μmol/[m2·s]) and relative placement of growth tiles.
The ATS was inoculated initially with a mixed algal community dominated by filamentous green algae obtained from periphyton sampling from local streams in Auburn, Alabama. The resulting algal turf was cultivated to maturity through multiple weeks of regular and repeated harvest and regrowth before experimentation. The reservoir was maintained at consistent nutrient concentrations through daily partial replacement of growth media from commercial F/2 media (ProLine; Pentair Co., Apopka, FL), modified for freshwater use at the typical loading concentration suggested by the manufacturer. Water parameters of pH and conductivity were monitored daily with a pH/EC probe (HI-98130; Hanna Instruments, Woonsocket, RI) and averaged 7.66 ± 0.16 and 0.59 ± 0.08 mS/cm, respectively, throughout the term of the experiments. Temperature was maintained at ambient laboratory temperatures and monitored daily at 23.6°C ± 0.5°C. Both dissolved P and N concentrations were monitored regularly using Insta-Test strips (LaMotte Co., Chestertown, MD), yielding 258 ± 63 ppb PO4-P and 2.42 ± 0.99 ppm NO3-N, respectively.
Sample analysis
In each of the three replications, the tiles were removed after 21 days, and each section from each tile was harvested in a multistep method that included vacuuming, mechanical scraping, and washing with low-pressure distilled water. Immediately after harvesting, the biomass recovered from each section was stored separately in vials. Dry mass analysis on biomass was performed by transferring the biomass from each vial to an aluminum weigh pan of known mass to oven dry at 105°C (Rice et al. 2012). Biomass data were normalized to mean daily productivity by dividing by the days of growth. The dry biomass data were then analyzed with parametric tests to understand the impact of surface texture feature size on biomass. The experiment was designed as a two-stage nested design wherein both section topography and replication are considered fixed factors (Fig. 5).
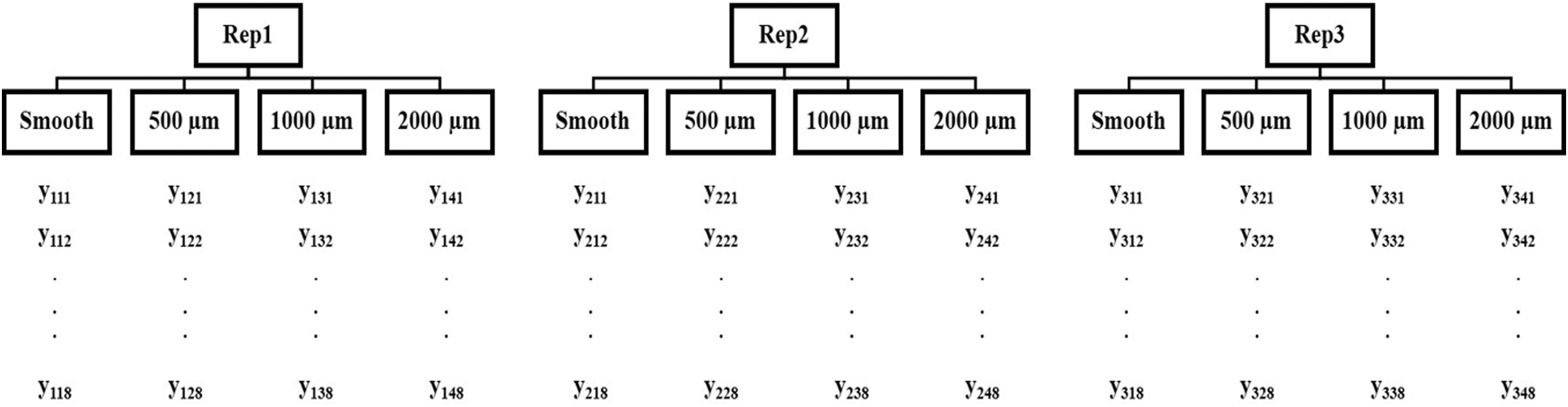
Two-stage nested design with topography feature size as nested factor under replications. (Montgomery, 2008)
Algal taxa identification was done to the lowest taxonomical level possible through compound microscope equipped with digital imaging (Motic Corp., Richmond, BC). Throughout the experiment, weekly grab samples were taken using tweezers from the growth substratum using tweezers for identification. The algae observed were keyed using standard identification keys (Prescott, 1984), confirming that the dominant taxa in all subsamples was Mougeotia scalaris Hassall for the term of the experiment.
Statistical analyses were performed on the response variable of mean biomass productivity per topographic section, expressed in units of mg/(cm·2·d). These data were first analyzed for homoscedasticity and other underlying analysis of variance (ANOVA) assumptions, including a test for equal variance and analysis of residual plots. After confirmation, parametric statistical tests were employed, including an ANOVA on the two-stage nested design using Minitab software (Minitab, Inc., State College, PA) followed by a Tukey test for multiple pairwise comparisons. These analyses were used to indicate algal preferences toward surface topography for attachment and growth of biofilms.
Results and Discussion
The largest average dry biomass productivity per section in each replication was found in the 500 μm topographic section, with the least amount of biomass always found in the smoothest one (Fig. 6). ANOVA (Table 1) demonstrates that the effect of topography on biomass productivity is significant (F = 13.175, p = 0.001) at the significance level of 0.01. It also confirms that there are no significant differences between the replicates of this experiment (F = 2.374, p = 0.149).
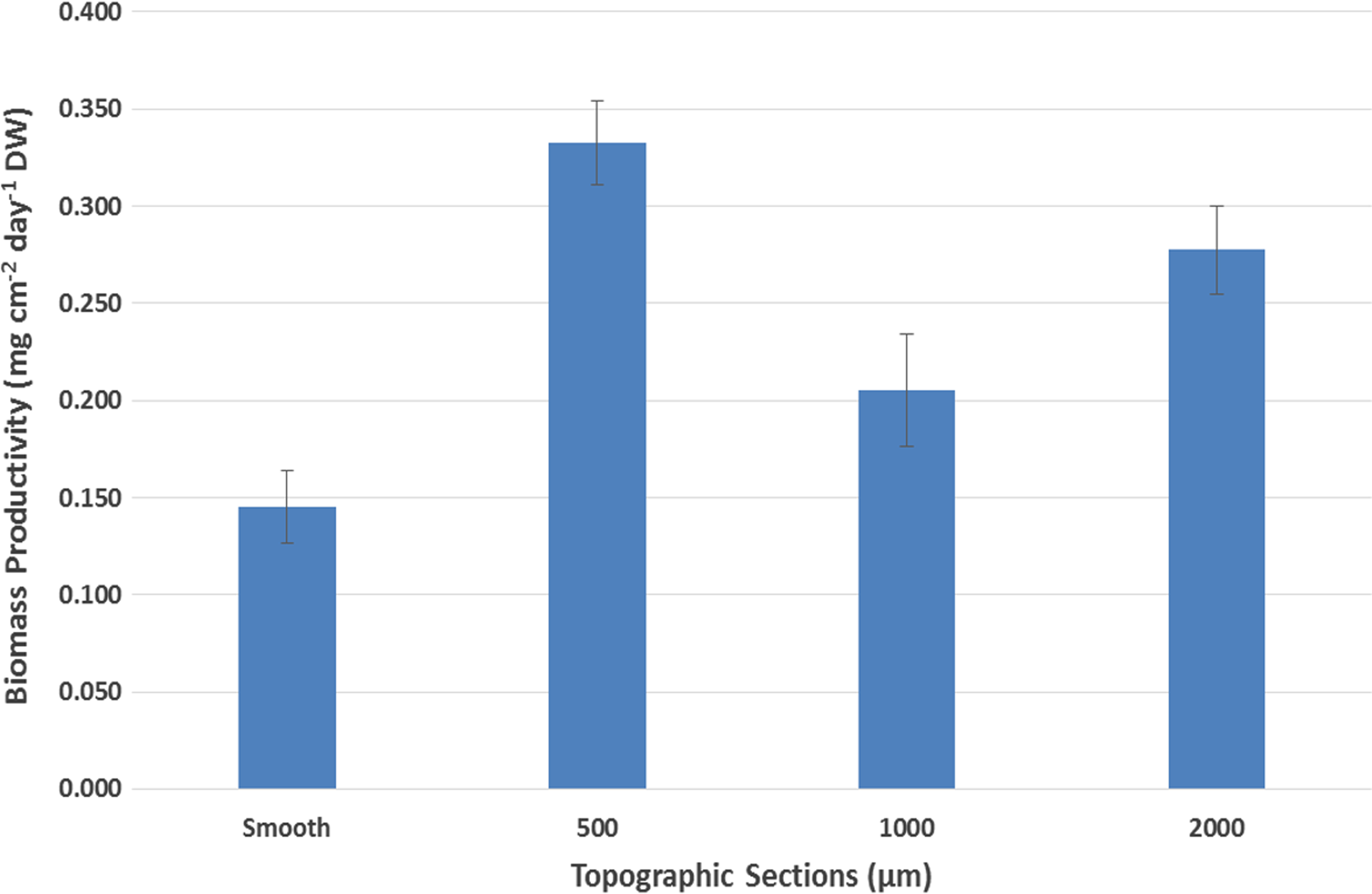
Average biomass dry weight productivity per section (n = 24).
Through microscopy and taxa identification, it was determined that the ATS presented a community of attached benthic filamentous algae dominated by M. scalaris. Surface topography was found to be a significant factor, with results showing the largest biomass productivity in the sections with feature sizes of 500 μm (0.333 g/[cm2·day]) and 2,000 μm (0.278 g/[cm2·day]), respectively, contrasting to 1,000 μm (0.205 g/[cm2·day]) and smooth area (0.145 g/[cm2·day]). To further discriminate the results, a Tukey multiple comparison test with 95% confidence interval was employed for all possible combinations of section topography. The results of the Tukey test statistically confirm that the topographies with feature sizes of 500 and 2,000 μm are preferred by M. scalaris. The test, however, cannot differentiate between the biomass productivity between these two topographies, so they are assumed as equally preferred. The smooth and 1,000 μm topographies present lower amounts and are grouped in the same category by the Tukey test.
Several explanations for the observed preferences toward specific surface topography may exist, and these include the following: differences in available surface area, localized microscale roughness of the material, effects of surface chemistry, and the effects of interstitial space distribution for refugia. With respect to the effect of localized microscale roughness, because all surfaces were made of the same material (ceramic clay), both microscale roughness and chemical characteristics were the same across all tiles and sections. With respect to the available surface area, it might be expected that sections with topography in the third dimension inherently present higher effective area per unit of nominal plan view area due to increased fractal geometric scaling, and that biomass productivity per unit area would directly relate to effective surface area. Indeed, smooth sections had an effective surface area 78% lower than topographic sections (2,704 mm2 compared with 4,827 mm2), and they consistently presented the lowest amount of biomass productivity. However, effective surface area alone does not explain differences in biomass productivity between the topographic sections (i.e., 500, 1,000, and 2,000 μm), since all three topographies present identical values of effective surface area by design, a function of the adjacent square-packed hemisphere pattern in which the effective surface area is constant regardless of hemisphere diameter (Fig. 7). Thus, in this experiment, topographic surface feature size does not artificially subsidize the effective area available for colonization.
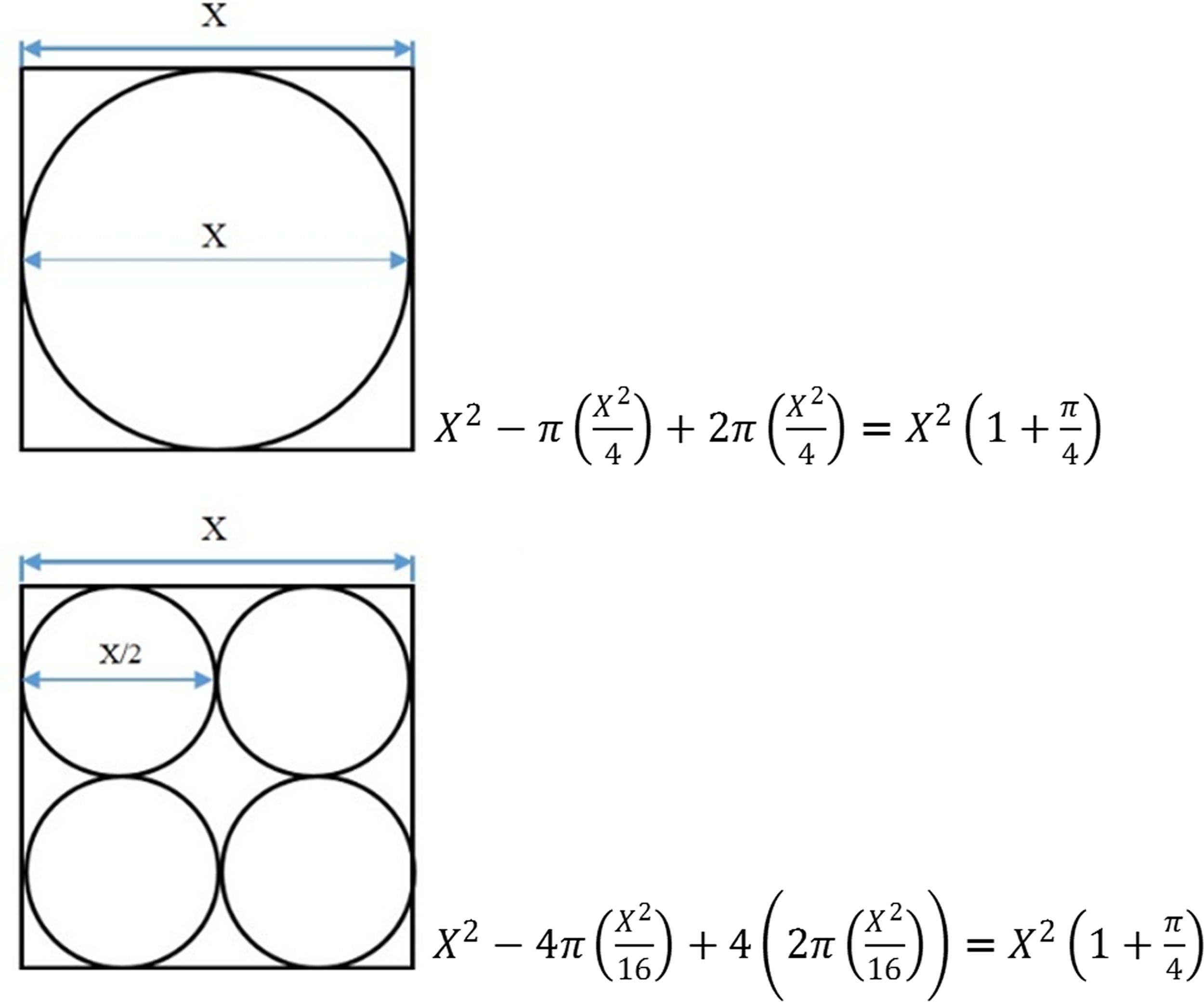
Calculation of surface area for two textures with adjacent hemispherical features.
These results suggest that there is an effect of the height and number of the topographic features upon interstitial spaces and refugia that is a factor for cell colonization and growth. Certainly, the possibility of an interaction between colonizing cell size and flow conditions induced by surface topography clearly exists. With an increasing number of topographic features per unit area, there are more sites where velocity boundary layer conditions are amenable to the settlement and attachment of a colonizing algal cell. With results showing increased productivity at both 500 μm and 2,000 μm sections, the relationship appears nonlinear, likely a result of the specific cell and surface geometries involved.
A further insight into this hypothesis can be obtained by observing the early colonization on the tiles (Fig. 8). Qualitative inspection suggests that the topography with the 2,000 μm features develops the largest amount of biomass earlier in the process, likely explained by the increased protection offered by the larger features to the turbulent flow conditions of the ATS. It may be that the larger features provide localized flow boundary conditions that are quiescent, allowing for greater cell settlement and attachment than other topographies.

Early colonization for 6 days on growth tiles.
The other topography that was preferred by M. scalaris was the 500 μm feature topography, which presents the largest amount of biomass at the end of the experimental run. A likely explanation is that it contains the largest number of interstitial spaces out of the four sections. The 500 μm section packs 2,704 hemispheres, which is 4 × greater than the 1,000 μm section (676 hemispheres) and 16 × greater than the 2,000 μm section (169 hemispheres). This greater number of hemispherical features, although providing an identical effective surface area, creates a greater number of sheltered low points (valleys) per unit area, where cells can withstand shear forces from overlying flow. These sites act as refugia to protect mature algal filaments once grown, and, when subjected to flow-induced sloughing, permit residual biomass to remain attached, thereby preventing irrecoverable loss of biomass and allowing rapid filament regeneration. Overall, the results argue for the balance between topographic feature height and distribution of interstitial space as strong determinants of algal biomass accrual on substratum, which has implications for the optimization of these parameters in the design of artificial substratum for cultivation processes.
Conclusions
In this study, the effect of surface topography on biomass productivity of an algal turf dominated by M. scalaris was studied. The findings support a nonlinear preference toward topographies with hemisphere features of 500 and 2,000 μm sizes. These dual preferences can be explained by the sheltered quiescent zones that arise between the larger 2,000 μm features that promote early cell settlement, and by the abundance of interstitial spaces available in the 500 μm section that provides residual biomass for quick filament regeneration. This suggests an approach for designing cultivation substratum topographies that balances feature height with the distribution of interstitial refugia space. Future work will focus on understanding the spatial, dimensional, shape, and frequency components of the surface features and their impact on both early attachment and on mature turfs.
Footnotes
Acknowledgments
The authors gratefully acknowledge Dr. Gary Wagoner of Auburn University for support in production of ceramic materials, and John Rains of Auburn University. Partial support for this research was provided by an equipment grant from the Auburn University Office of the Vice President of Research. In-kind support was provided by the Auburn University College of Agriculture, the Auburn University Samuel Ginn College of Engineering, and the Alabama Agricultural Experiment Station.
Author Disclosure Statement
No conflicts of interest exist.