
Abstract
Abstract
This article assesses nitrifying and heterotrophic activity potentials, aiming to ward off future efficiency losses in biological treatment systems. For this purpose, we used biomass developed in two-hybrid fluidized bed reactors (FBRs) treating vinasse from ethanol and sugar beet production. FBRs filled with sepiolite and granular activated carbon (GAC) were used with short cycle aeration (25/15 and 15/15 min) and different hydraulic retention times (4.9 and 7.4 h). At the end of each operational condition, attached and suspended fractions of biomass from both FBRs were collected for activity bioassays. Specific nitrification bioassays suggested that nitrification performance (53–100% for sepiolite FBR and 72–93% for GAC FBR) was associated with the limit of the nitrifiers' potential when the environmental conditions were favorable. On the other hand, low denitrification efficiencies observed in the FBRs (25–44% for sepiolite FBR and 22–65% for GAC FBR) occurred due to low electron availability and the maintenance of residual dissolved oxygen, even during the off-periods, as denitrifiers presented high potential in denitrification bioassays. Assessing biomass potential enabled us to identify at which moment biological capability was close to its limit before loss of efficiency occurred, such as during the reactor start-up or loading reduction/increase. Thus, specific activity bioassays are an important tool for diagnosing biomass potential from simultaneous nitrification denitrification systems to perform aerobic, anaerobic, and anoxic processes.
Introduction
C
A possible alternative to reduce this construction and operating costs is by using compact systems that integrate nitrogen and organic matter removal processes within a single unit by simultaneous nitrification and denitrification (SND) (Münch et al., 1996; Pochana and Keller, 1999; Fdz-Polanco et al., 2000; Zeng et al., 2003; Chiu et al., 2007). SND systems are able to maintain a pH balance that may help develop different microorganism populations (Yoo et al., 1999). Effective total nitrogen (total-N) removal was reached by using different types of inoculum, such as aerobic (Yoo et al., 1999), activated sludge (Canto et al., 2008; Barana et al., 2013), and mixed aerobic–anaerobic sludge (Sen and Dentel, 1998; Santos et al., 2016). Moreover, if intermittent aeration is adopted, a significant reduction in biomass synthesis (Chen et al., 2000) and inhibitory effects, such as those caused by nitrite (Cervantes et al., 2001; Philips et al., 2002), can be achieved. Canto et al. (2008) compared the efficiency of nitrogen and organic matter removal in continuous and intermittent aerated sequencing fed-batch reactors (SBRs) inoculated with continuously aerated activated sludge. The authors observed the best performance in terms of total-N removal (80%) for the intermittent aeration cycle (aeration of 3 h, maintaining alternation between periods with and without aeration). At continuous aeration mode, these authors detected a lack of carbon source for heterotrophic denitrification due to fast consumption by aerobic organic matter oxidizers (Canto et al., 2008).
Ruiz et al. (2006) reported poor sludge sedimentation as one of the disadvantages of intermittent aeration for the SND process. However, using support materials for biomass development (biofilm) can minimize this effect (Metcalf and Eddy, 2003). It has been demonstrated that attached biomass is less affected than suspended biomass by changes in environmental conditions, such as temperature, pH, nutrient concentrations, metabolic products, and toxic substances (Lazarova and Manem, 1995; Bishop, 1997; Metcalf and Eddy, 2003). Moreover, the enhanced resistance to oxygen mass transfer into the biofilm contributes to maintaining an internal anoxic environment, even during the aerated period (Wang et al., 2010; Barana et al., 2013; Santos et al., 2016).
Among the reactor configurations in which immobilized biomass can be used, fluidized-bed reactors (FBRs) are an interesting alternative (Bello et al., 2017). Due to considerable amounts of solid carriers inside FBRs, suspended and attached biomass can concomitantly be retained, resulting in hybrid growth systems (Jianlong et al., 2000; Martín et al., 2007). Segregation among suspended and attached biomass enables spatial distribution between fast and slow growing microorganisms (Yu et al., 2001; Martín et al., 2007). Thus, investigating how nitrification and heterotrophic metabolism behave in SND systems is important to predict their overall nitrogen removal performance. Studies performing nitrifying and denitrifying activity tests are described in the literature (Tarre and Green, 2004; Pala and Bolukbas, 2005; Chiu et al., 2007) directly relating these data with results from continuous reactor operations. However, there are no studies in the literature determining the nitrifying and heterotrophic activities of the suspended and attached biomass that can be established in hybrid SND systems and their association to the understanding of how the loss of efficiency could be predicted.
A better investigation of nitrification and denitrification potentials of both biomass fractions developed in SND systems can be useful to identify when the biomass potential is close to its limit, warding off the loss of system efficiency. These results, and the reactor monitoring data, can support decision making for reactor start-ups, loading reduction or increase, and others. Therefore, our aim was to estimate the potential of nitrifying and heterotrophic activities (oxygen and nitrate/nitrite as electron acceptors) in the biomass developed in two-hybrid FBRs treating vinasse from ethanol and sugar beet production.
Material and Methods
Experimental setup
Fluidized bed reactors
Two FBRs were operated with a total volume of 1.7 L, an internal diameter of 5.4 cm and a height of 75 cm, resulting in a working volume of 1.3 L (Fig. 1). One FBR was partially filled with sepiolite (TOLSA S.A, 30/60) and the other with granular activated carbon (GAC) (Cullar-D, 20/40 US Mesh). The sepiolite and GAC solid media had particle sizes of 0.32 and 0.47 cm, densities of 2.18 and 1.45 kg/L, and porosities of 0.7 and 0.65, respectively (Martín et al., 2007). Nitrifying sludge from the municipal WWTP located in Valladolid (Spain) was used as inoculum. The recirculation flow was adjusted to achieve, at least, 30% (v/v) of bed expansion in both reactors. Oxygen was intermittently supplied by air pumps and different on/off periods were tested (Table 1).

Schematic of experimental apparatus used for continuous operation. FBRs, fluidized bed reactors.
COD, chemical oxygen demand; GAC, granular activated carbon; HRT, hydraulic retention time.
Monitoring the FBRs
Operational conditions and feeding
Before starting this study, sepiolite and GAC FBRs were operated over 15 months, dispensing a previous adaptation phase. Therefore, the operating procedure of this study was divided into three stages: after a long operation (Condition 1), after reinoculation (Condition 2), and submitted to different aeration cycles (Condition 3). Overall nitrogen and organic matter performances were evaluated by examining FBR effluent samples twice a week, by reaching a total of 15–25 sample points in each operational phase. As a high recirculation flow was adopted to allow the desired bed expansion in both FBRs, these systems were operated as completely mixed units. This fact ensured effluent characteristics within a small variation range, regardless of the aeration period.
The FBRs were maintained at 33°C ± 2°C and continuously fed with vinasse from ethanol and sugar beet production. The wastewater was supplemented with ammonium chloride (Table 1). Sodium bicarbonate was added to ensure the theoretical alkalinity demand required by nitrification (7.14 mg CaCO3 per mg of TKN oxidized). Hydraulic retention time (HRT) was initially set to 4.8 ± 0.1 h and 7.4 ± 0.1 h for sepiolite and GAC reactors, respectively. However, to investigate the SND performance with the same nitrogen loading rate as in the GAC system, the HRT in Condition 3 for sepiolite FBR was increased to 7.4 h. Initial total biomass concentrations (considering the attached and suspended fractions) were 12.6 and 12.4 gVSS/L for sepiolite and GAC systems, respectively.
Nevertheless, after 47 days of operation (Condition 1), reactors were reinoculated (by adding sludge and support material). This was necessary due to not being able to control the pumping system. The biomass concentration was recovered in the reactor with sepiolite after its reinoculation, but was lost again in the GAC reactor. However, the system's recovery was observed. The calculation procedure adopted in this study can be found in the Supplementary Data. Table 1 shows the operational conditions used for both support materials.
Physical-chemical and biomass analyses
The monitoring parameters used during FBRs operation were as follows: biological oxygen demand (BOD); chemical oxygen demand (COD); volatile suspended solids (VSS); ammonium nitrogen (NH4+-N), nitrite nitrogen (NO2−-N), nitrate nitrogen (NO3−-N), and dissolved oxygen (DO) concentration. For VSS measurements at the end of each operational condition, biomass was detached from support materials by sonication, as performed by Martín et al. (2007). We determined the anions (nitrite and nitrate) by high-performance liquid chromatography. The DO concentration was periodically measured inside both reactors using a multi-probe system (YSI 556). All other analyses were performed according to Standard Methods for the Examination of Water and Wastewater (APHA, 2012).
Study of nitrifying and heterotrophic activities
At the end of each experimental condition, we conducted specific activity bioassays of nitrification and heterotrophic metabolisms (oxygen and nitrate/nitrite as electron acceptors) to evaluate the biomass potential to perform aerobic, anaerobic, and anoxic processes. Biomass samples (attached and suspended fractions) were collected by periodically withdrawing mixed liquor together with bioparticles from inside the two reactors. All activity assays were carried out at 25°C and the reaction liquids were supplemented with a micronutrient solution, as described by Santos et al. (2016).
We ran biomass nitrifying activity tests in 20-mL respirometry flasks. An oximeter (ORION 870; ThermoFisher) was used for DO monitoring. An initial DO concentration near to saturation (>8 mgO2/L) was adopted before starting the test. The potential use of oxygen as an electron acceptor for acetate oxidation (heterotrophic activity) was concomitantly explored at the attached and suspended biomass from both systems (Table 2). NH4+-N and NO2−-N pathways were evaluated based on the rate of oxygen consumption, with NH4+-N and NO2−-N sources, respectively.
In these tests we evaluated the potential of using nitrate/nitrite as electron acceptors in 100-mL sealed flasks. Heterotrophic denitrification by NO3−-N and NO2−-N pathways was estimated by the evolution of N2 production. Acetate was used as the electron donor. As recommended by Villaverde et al. (2000) and Chudoba et al. (1992), the initial F/M ratio was <3, aiming to avoid biomass growth during the bioassays. To maintain anaerobic conditions and also minimize the interference of nitrogen gas (N2), before beginning each anoxic test, the flask was bubbled with helium for 15 min. The N2 generation during the denitrification tests was measured by gas chromatography on a Hewlett Packard 6,890 Series II equipped with a thermal conductivity detector (TCD) and a Varian CP3800 equipped with a TCD. Helium was used as the carrier gas.
Table 2 depicts the initial substrate and biomass concentrations for the nitrification (NH4+-N and NO2−-N pathways) and heterotrophic metabolisms (oxygen and NO3-N/NO2-N as electron acceptors) bioassays. Activity bioassays were performed when an apparent or dynamic stationary state was established in each operational condition. Each test was carried out in duplicate, with a difference (in terms of the rate of oxygen consumption, for nitrifying bioassays and N2 generation for denitrifying bioassays) between the duplicates below 15%. Biomass samples (for duplicates) were collected with, at least, 1 week of interval, since its removal could negatively affect the reactors' performance.
Results and Discussion
Nitrogen and carbon removal
After a stability period for COD removal (15 months), the reactors were continuously operated for another 130 days by using vinasse supplemented with a nitrogen source. Table 3 summarizes the data obtained during the monitoring of each FBR. As nitrite was not detected in the effluent samples for both support materials, the total-N was determined by the sum of NH4+-N and NO3−-N effluent forms.
DO, dissolved oxygen.
COD removal efficiencies obtained for both systems were similar in all the experimental phases (Table 3). To further analyze the nitrogen conversion by the SND process, the applied and removed nitrogen rates were also calculated (Table 4). The highest total-N removals were achieved in Condition 1 for both FBRs (Table 4). For sepiolite FBR, the total-N and organic loading removed were 0.13 gN/(L·day) and 1.91 gCOD/(L·day), respectively. In the case of GAC FBR, the total-N and organic loading removed were 0.11 gN/(L·day) and 1.26 gCOD/(L·day), respectively. Zeng et al. (2003), who also operated an SND system subjected to intermittent aeration (3 h aerated and 1 h nonaerated) reached the oxidation of 0.04 gN/(L·day) without intermediate compound accumulation when treating synthetic wastewater (sodium acetate as a carbon source and affluent COD equal to 400 mg/L) in an SBR. DO concentration of 0.5 mgO2/L was maintained during the aerobic phases (Zeng et al., 2003).
Table 3 shows a global biomass loss of 33.3% and 64.5% at the end of the operation of sepiolite and GAC systems, respectively. We obtained these values by comparing the initial VSS concentration of each FBR (determined after inoculation) with VSS measurements at the end of Condition 3.
Temporal profiles of total-N loading removed (data not shown) indicated that the total-N removal efficiencies slumped at the final days of each operational condition. This fact is associated with the progressive loss of biomass observed in both FBRs (Table 3). Attached fractions were more affected (Table 3), which is directly associated with the loss of support materials. Despite the progressive loss of biomass observed in both FBRs (Table 3), better maintenance of biomass adhered to sepiolite can be explained by its lower particle size and specific gravity.
Nitrification and nitrifying rates in FBRs
Table 5 summarizes ranges of specific nitrifying rates calculated for immobilized and suspended biomass from two reactors at the end of each experimental phase. Generally, the attached biomass fraction presented higher nitrifying rates for NO2−-N oxidation than for the NH4-N oxidation (Table 5). On the other hand, the opposite was observed for the suspended fractions, in which higher conversion rates were observed for NH4+-N oxidation (Table 5). These findings corroborate with results from Carvalho et al. (2006), who carried out a microbial quantification of ammonium oxidizing bacteria (AOB) and nitrite oxidizing bacteria (NOB) by fluorescence in situ hybridization in flocs and granules from an SND system. The authors observed that the biggest fraction of NOB appeared to grow in the more attached form (granules), whereas AOB appeared to be more abundant in the flocs. Therefore, the FBR configuration and the microorganism arrangement into the biofilm clearly influenced the oxygen mass transfer (Sevillano et al., 2008), microorganism selection at the attached and suspended fractions (Carvalho et al., 2006; Martín et al., 2007) and, therefore, the nitrification efficiencies obtained for each condition. The coexistence of immobilized and suspended biomass is correlated to the segregation between fast and slow growing microorganisms. While the attached fraction favors an effective retention of slow growing nitrifiers (NOB) the suspended fraction is formed by fast growing nitrifiers (Carvalho et al., 2006).
Nitrifying activities were measured in terms of mgO2/(gSSV·h). In the case of NH4-N and NO2-N pathways, the obtained activities were presented in terms of mg NH4-N/(gVSS·h), by considering an oxygen demand of 3.7 mgO2/gNH4-N oxidized and 1.14 mgO2/gNO2-N oxidized (Metcalf and Eddy, 2003).
FBR, fluidized bed reactor.
Concerning sepiolite FBR, we observed an increase in nitrification efficiency in Conditions 2 and 3 (Table 3), resulting in higher specific NH4-N loading nitrified values (Table 4). In Condition 2, this fact was associated to biomass reinoculation, achieving a specific NH4-N loading nitrified of 0.59 gN/(gVSS·L·h). Surprisingly, maximum nitrification efficiency was achieved in sepiolite FBR during Condition 3 despite the biomass loss (Table 3). This result could be attributed to the increase in the nitrifying activity rate from the range of 1.6–2.0 to 2.4–2.9 mgN/(gVSS·h) (Table 5).
The GAC reactor presented the highest global nitrification efficiencies, which is in accordance with the specific NH4+-N loading nitrified observed during the reactor's operation (Table 4). Despite the decrease in the microorganism mass in the GAC reactor in Condition 2, nitrification efficiency was enhanced, which was corroborated by the increase in specific NH4+-N loading nitrified from 0.57 to 1.21 gN/(gVSS·h) (Table 4). In Condition 3, the decrease of the nitrification efficiency in the system can be associated with the loss of biomass that occurred mainly for the adhered fraction. However, this reduction was not reflected in the specific NH4-N loading nitrified value observed in Condition 3, as it was maintained (Table 4). Table 5 illustrates that the suspended fraction of biomass increased its participation significantly in the global nitrification rate (25% of global rate). Therefore, the nitrifying activity bioassays markedly showed the potential of suspended biomass to perform an aerobic process. Concerning practical decision making, it can be suggested that the discard of suspended biomass should be controlled through operational periods, as excessive suspended biomass can cause resistance to oxygen mass transfer into liquid medium (Fu et al., 2010). Moreover, if not controlled, suspended biomass can compete with a denitrifying community to oxidize the bioavailable organic matter present in influent (Metcalf and Eddy, 2003).
Denitrification and heterotrophic metabolism rates
This study found that even with elevated nitrification efficiencies in the FBRs, the total-N removal efficiencies were partial, resulting in residual NO3-N in effluent samples (Table 3). This may have occurred when the anoxic periods were insufficient to ensure complete NO3-N reduction (Barana et al., 2013), or there was a lack of bioavailable electron donors for heterotrophic denitrification (Barana et al., 2013; Santos et al., 2016) and/or the anoxic zone was not developed (Zeng et al., 2003; Satoh et al., 2004). In accordance with these hypotheses, we observed significant heterotrophic activity rates, using oxygen as an electron acceptor, by activity bioassays for both FRBs throughout the whole experimental period (Table 5). Oxidation of organic matter proceeds preferentially with oxygen as an electron acceptor, where NO3− and other inorganic compounds are used as alternative electron acceptors only in anaerobic/anoxic conditions (Thauer et al., 1977; Strohm et al., 2007). Heterotrophic activity rates were similar for the attached and suspended biomass for both tested supports. In Condition 3, in which the F/M ratios in FBRs were the highest (0.16 and 0.30 gCOD/(gVSS·day) for sepiolite and GAC reactors, respectively), similar COD removal efficiencies were achieved for both FBR. This suggests biomass capacity for the biodegradation of the influent organic matter, regardless of the carrier material characteristics (Table 3). Therefore, the high COD removal efficiencies (Table 3) observed at each operational condition were confirmed in the heterotrophic activity bioassays.
Table 3 shows the DO concentrations during aerated and nonaerated phases. In Condition 1, in which the highest total-N removal efficiencies were achieved for both reactors (Table 3), we observed an efficient maintenance of low DO concentrations in the liquid medium during the aerobic period (1.8 ± 0.1 mgO2/L). Consequently, low residual DO concentration was achieved in the consequent nonaerated phase (0.13 ± 0.1 mgO2/L) (Table 3). These DO dynamics were not maintained in the subsequent conditions (high residual DO concentration during nonaerated phases). Moreover, the decrease in the on/off-periods for the GAC reactor in Condition 3 from 25/15 to 15/15 min (Table 3), which aimed at reducing the oxygen supply and the consequent limitation of organic matter oxidation metabolism, did not show positive effects for total-N removal efficiency. By contrast, residual DO during the nonaerated phase was kept near to 1 mgO2/L. As a result, the total-N removal efficiency in the GAC reactor dropped from 44% (Condition 2) to 22% (Condition 3). Zeng et al. (2003) operated an SBR by alternating anaerobic–aerobic phases. A decrease in SND efficiency from 98% to 62% and 51% was observed when the DO concentration (during aerobic phase) rose from 0.5 to 1.25 and 2.5 mgO2/L, respectively. Moreover, the total-N loading removed dropped from 0.10 to 0.04 gN/(L·day) when external carbon sources were not added at the beginning of the anoxic period.
Barana et al. (2013) evaluated the COD and nitrogen removal from an anaerobic reactor effluent in a structured-bed reactor subjected to recirculation and intermittent aeration (SBRRIA). The SBRRIA was continuously fed with an HRT over 24 h and the duration of the aerobic periods was gradually reduced. The authors observed nitrification efficiencies of at least 90% during the entire operation. However, only after a longer nonaerated period (1 h aerated and 2 h nonaerated) was the total-N removal enhanced, achieving an efficiency of 62%. Besides the longer nonaerated phase, the observed low denitrification efficiency was attributed to the absence of readily biodegradable organic matter (Barana et al., 2013).
The organic source used as an electron donor for denitrification can also influence the denitrifying activity rate, as observed by Henze (1991), in which denitrification was more efficient with methanol than ethanol. Pala and Bolukbas (2005) observed that acetate was a more effective electron donor than glucose. In the present study, despite the remaining COD effluent, BOD analyses, performed sporadically, indicated biodegradability values close to zero in the final effluent from GAC and sepiolite reactors (data not shown). This may be one of the causes of the reduced total-N removal.
Although biofilm thickness monitoring was not carried out, the total-N loading removed could also be influenced by the inefficient development of the aerobic/anoxic biofilm (Zeng et al., 2003; Satoh et al., 2004). Satoh et al. (2004) monitored the DO profile observing different stages of biofilm development in a membrane aerated biofilm reactor. The authors noted that the anoxic zone development of the biofilm coincided with the rise in total-N removal rates. Liu and Tay (2001) evaluated the effect of shear stress in biofilm developed in a rotating disc reactor. They observed that hydrodynamic conditions had a significant influence on the density and thickness of the biofilm. As high superficial velocities (obtained by recirculation flow) were used in FBR systems to suspend the bioparticles (Sevillano et al., 2008; Tisa et al., 2014; Bello et al., 2017), it seems possible that fluidization negatively affected the development of an effective anoxic zone in the attached biomass.
Table 6 summarizes ranges of specific denitrifying rates for immobilized and suspended biomass from two FBRs at the end of each experimental phase. Observed ranges of denitrifying activity rates (Table 6) were much higher than the specific Total-N loading removed achieved in the FBRs (Table 4). Figures 2 and 3 depict N2 profiles measured during anoxic acetate oxidation by using NO3 and NO2 as electron acceptors, respectively. For each bioassay, each data point represents an average between results from duplicate batch tests. To better clarify the tendency of N2 production, Origin software was used to extrapolate a best-fit continuous curve.
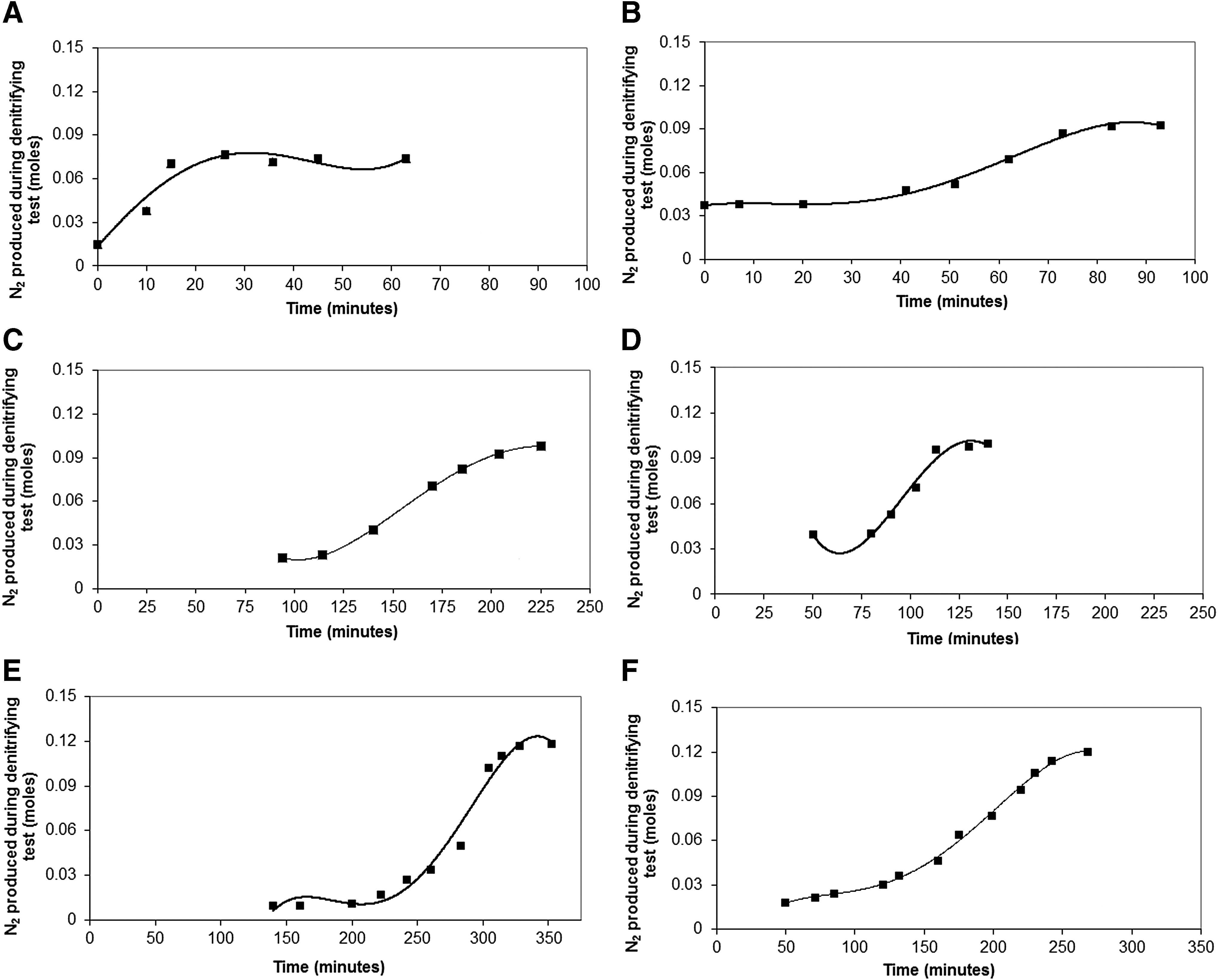
N2 production during denitrifying activity tests with attached biomass using NO3 as nitrogen source.

N2 production during denitrifying activity tests with attached biomass using NO2 as nitrogen source.
Considering the profile of N2 production during the denitrifying activity assays in Condition 3, longer lag phases were observed for sepiolite and GAC supports (Figs. 2C–F and 3C–F). This shows that the denitrification potential was negatively influenced by the applied operational conditions, requiring an extended period (lag phase) to recover its denitrifying activity during the bioassays. According to Table 3, even for a nitrification efficiency of 100% observed in Condition 3 for sepiolite FBR, denitrification efficiency (in terms of total-N removal efficiency) was the lowest of the period (25% ± 17%), resulting in a total-N loading removed of 0.03 gN/(L··day). However, in this operational phase, the denitrifying activity rates were significant (Table 6). This also supports our main premise that proposed bioassays align with routine monitoring methods and could provide useful supplemental data for operational decision making.
The difference among denitrification rates observed in specific activity tests and the reactor operation is consistent with those of previous studies. Timmermans and Van Haute (1983) and Christensson et al. (1994) obtained denitrifying activity rates of 1.3 and 3.3 gNOx-N/(gVSS·day), respectively. These values are much better than some systems operating in an SND condition, in which the denitrifying activity rates varied from 0.037 to 0.64 gNOx-N/(gVSS·day) (Pala and Bolukbas, 2005; Chiu et al., 2007).
Conclusions
Considering an adequate DO supply, the nitrification performance in continuous FBR was associated with the limit of the potential of the nitrifying microrganisms, which could be influenced by biomass loss, reinoculation, or different NH4-N loading rates. The total-N removal efficiencies achieved at the lab-scale systems were worse than the potentialities of the denitrification processes, explored by the specific activity bioassays, as imposed on-off aeration periods and operational conditions were not adequate. Even with high nitrification efficiencies, the denitrification rates were partial, characterized by residual nitrate in the effluent. Therefore, we can conclude that the inefficient SND performance observed in this study was not correlated to the characteristics of support materials.
However, the specific activity bioassays were successful in identifying the biomass potential from FBR systems to perform aerobic, anaerobic, and anoxic processes. These results, associated with the reactor monitoring data, can support decision making at different moments of operating wastewater treatment systems, warding off loss efficiency.
Footnotes
Acknowledgments
We gratefully acknowledge the financial support from the Spanish CICYT (project PPQ2003-09688-C02-01) and the National Council for Scientific and Technological Development (CNPq/Brazil).
Author Disclosure Statement
The authors declare that they do not have any conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.