
Abstract
Abstract
The aim of this study was to evaluate application of a crude enzymatic extract from the mixed culture of Trametes maxima (T. maxima) and Paecilomyces carneus (P. carneus) in removal of phenanthrene (PHE) from water. Since PHE belong to the polycyclic aromatic hydrocarbons (PAHs), it has been used as a model compound for bioremediation treatments. PAHs are highly stable molecules, and are resistant to conventional wastewater treatment. Production of crude extract from a mixed culture of T. maxima and P. carneus does not require special conditions yet the use of 20 g/L of glycerol as carbon source enhances laccase (Lcc) and manganese-dependent peroxidase (MnP) activities. Application of crude extracts becomes feasible because it does not require further purification. The crude enzymes entrapped in alginate beads in a fixed-bed packed column reached 94.3% ± 0.7% of PHE removal after 13 h of continuous treatment under nonsterile conditions, while the alginate beads without enzymes reached only 45.3% ± 23% of PHE removal. The difference between the removal of alginate beads with and without enzymes was the oxidative activity on the surface of the alginate beads, which was observed in the increase in the C═O group and in the X-ray spectroscopy analyses of the C 1s and O 1s regions. Beads without enzymes had less C═O groups. Polyoxyethylene sorbitan monooleate and dimethyl sulfoxide also influence enzymatic activity to carry out a level of PHE removal similar to a photocatalytic treatment.
Introduction
A
Among the POPs, polycyclic aromatic hydrocarbons (PAHs) are ubiquitous in the environment, and they are priority pollutants to be eliminated from water due to their teratogenic, mutagenic, and carcinogenic properties. The lack of biodegradability and toxicity of PAHs is influenced by the increase in the number of benzene rings; also, they are not essential nutrients for microbial growth (Shen et al., 2015). The PAHs have a high fusion and boiling temperature and a low vapor tension, all of which are inversely proportional to the number of rings or molecular mass. PAHs have a high stability in nonlinear structures. Despite PAHs' hydrophobic nature, the ones with three to five rings can be found in liquid phases attached to suspended particles (Torretta, 2012). The solubility of aromatic hydrocarbons is related to their molecular weight, decreasing with the number of aromatic rings (Adeniji et al., 2017). Phenanthrene (PHE) can easily be washed off from the soil and it presents a higher activity in water in comparison with PAHs of higher molecular weight (Ligaray et al., 2016). The PHE has been used as a model for degradation studies of the PAHs with carcinogenic properties. The fundamental objectives of PAH remediation are control, containment, and finally their removal or destruction.
White-rot fungi secrete hydrolytic and oxidative enzymes (Elisashvili and Kachlishvili, 2009; Lee et al., 2014). Among such enzymes are cellobiose dehydrogenase, glyoxal oxidase, aryl-alcohol oxidase, oxalate decarboxylase, NAD-dependent formate dehydrogenase, P450 monooxygenase, glutathione reductase, copper radical oxidase, isoamyl alcohol oxidase, peroxiredoxins, benzoquinone reductase, pyranose 2-oxidase, multicopper oxidase, glutathione S-transferase, glucose oxidase, and manganese superoxide dismutase (Manavalan et al., 2014). The major extracellular lignin-degrading enzymes are lignin peroxidase, versatile peroxidase, laccase (Lcc), and manganese-dependent peroxidase (MnP) (Manavalan et al., 2014).
Lignin-modifying enzymes present low specificity for their substrate, which means that they are able to oxidize molecules with a chemical structure similar to their substrates; therefore, they are used as biocatalyst in the synthesis and degradation of complex molecules such as PAHs (Mogharabi and Faramarzi, 2014). In continuous operations, the entrapment of enzymes presents the advantage of being resistant to oxidizing agents such as H2O2, O3, metals, metal oxides, and metal salts (Oller et al., 2011). The oxidizing agents are toxic to microorganisms affecting the efficiency of the system. The MnP and Lcc enzymes have wide industrial applications. The substrates for both enzymes are different; while Lcc (Ionization potential 7.45 eV) catalyzes bond cleavage in low molecular weight phenolic compounds, MnP (Ionization potential 8.19 eV) catalyzes alkyl-aryl cleavage (Manavalan et al., 2014). The redox potential of Lcc and MnP is higher than other enzymes; therefore, the by-products from Lcc and MnP catalytic reactions are the substrates of the other enzymes. To maximize their production, several strategies have been implemented such as heterologous expression (Lee et al., 2016; Wang et al., 2016) and the use of chemical (Kachlishvili et al., 2014) or biological inductors (Li et al., 2011). The use of biological stress inductors consists of the interaction of two or more microorganisms to enhance the enzymatic activity (Netzker et al., 2015).
Application of free enzymes is limited by environmental conditions. The enzymatic cocktails have found their main application in the treatment of lignocellulosic biomass. The mixture of crude enzymes from Cladosporium cladosporioides Ch2-2 with commercial xylanase enhanced the saccharification of milled Jerusalem artichoke stalks, and it showed a higher selective delignification than the white-rot fungi (Ji et al., 2014). In the case of municipal wastewaters in which heavy metals, organic compounds, and proteolytic enzymes are present, the enzymes are subjected to inactivation (Le et al., 2016). The use of free Lcc cocktail has been successfully applied by Garcia-Morales et al. (2015) in the removal of the endocrine disruptors (bisphenol A, triclosan, 4-nonylphenol, and 17-α-ethinyl estradiol) from the groundwater. However, the enzymatic activity was affected by the presence of ions in the groundwater (Garcia-Morales et al., 2015). To degrade PAHs, the enzymes must have a high activity and keep their operational stability. Immobilization of Lcc in alginate beads with Fe2O3 nanoparticles allowed the reusability and removal of the Remazol Brilliant Blue R dye and triclosan in wastewater (Le et al., 2016).
Enzyme immobilization allows the reuse and recovery of enzymes, as well as their application in continuous processes. In the gel entrapment, the enzymes are surrounded by a polymeric matrix. The procedure does not require special conditions, and it has a minimum of structural alteration of the enzyme bringing resistance to pH effects, organic solvents, and microbial attacks (Rao et al., 2014). The term “alginate” refers to a family of unbranched polysaccharides of 1,4′-linked β-
In this research, we proposed that the removal of PHE for a water matrix is feasible in a continuous treatment by a crude enzymatic extract without further purification. To test PHE removal, we use a crude enzymatic extract from the mixed culture of Trametes maxima (T. maxima) and Paecilomyces carneus (P. carneus). The strains were collected in Mexico; therefore, they do not need special adaptation conditions. In the mixed culture, P. carneus was used as a biological stress inductor to enhance enzyme production. Among the extracellular crude enzymes, Lcc and MnP enzymes were followed due to their low substrate specificity. All the crude enzymes precipitated were entrapped on alginate beads. The activity of the entrapped crude enzymatic extract on alginate beads in the removal of PHE was tested under nonsterile conditions in the presence of dimethyl sulfoxide (DMSO) and polyoxyethylene sorbitan monooleate (Tween 80). DMSO and Tween 80 were evaluated as solubilizing agents and facilitators of enzymatic activity in the PHE removal. The enzymatic activity on the surface of the alginate beads was followed by X-ray photoelectron spectroscopy (XPS). The results were used in further studies, coupling the current treatment to a TiO2-C-Ag/UVA/photocatalytic system.
Materials and Methods
Chemicals
Potato dextrose agar and yeast extract were from BD Bioxin (Becton, Dickinson and Company, Cuatitlán Izcalli, México) and BD Bacto (Becton, Dickinson and Company), respectively. The glucose and glycerol were reagent grade from J.T. Baker (Phillipsburg, NJ), as well as the salts used in the culture medium, buffers, and the crosslinking solution used on alginate beads. The sodium salt of alginic acid from brown algae and the glutaraldehyde (50% wt. in H2O) were from Sigma-Aldrich (Toluca, Mexico). The cOmplete™, Mini Protease Inhibitor Cocktail was from Roche (Sigma-Aldrich). The PHE (90%), DMSO (99.5%), and Tween 80 (average mol. wt. 1,310) were from Sigma-Aldrich. Dichloromethane (DCM), acetonitrile (ACN), acetone (ACT), and methanol (MeOH) were used without further purification, and they were high-performance liquid chromatography grade from Honeywell (Wabash, IN).
Production of crude extract by mixed culture
Strains of T. maxima (GenBank HF947516) and P. carneus (GenBank HF947521) were donated by the Laboratorio de Micromicetos of the Instituto de Ecología, A.C. Xalapa, Veracruz, México. The strains were kept in potato dextrose agar at 4°C until inoculation in a liquid culture. The medium used to induce the enzymes was modified from Revankar and Lele (2007). The medium was composed of glucose (20 g/L), yeast extract (2.5 g/L), H2PO4 (1.0 g/L), (NH4)SO4 (0.05 g/L), MgSO4 (0.5 g/L), CaCl2 (0.01 g/L), FeSO4 (0.01 g/L), MnSO4 (0.001 g/L), ZnSO4 (0.001 g/L), and CuSO4 (0.002 g/L). Four 5-mm diameter disks with full mycelium growth of T. maxima were inoculated into 500 mL Erlenmeyer flasks with 240 mL of liquid medium. The mixed culture was obtained by inoculating P. carneus after 48 h of culturing T. maxima. To follow the enzymatic activity, 4 mL samples were taken at different times. The samples were centrifuged at 5,070 g for 20 min, before the analysis of reducing sugars (DNS technique), proteins (Bradford reagent), Lcc (Wolfenden and Willson, 1982), and MnP (Minussi et al., 2001). Lcc and MnP activity was measured at room temperature in a UV-Vis Spectrometer (Model UV-1800; Shimadzu). In the case of Lcc activity, the reaction mixture contained 600 μL of the supernatant and 300 μL of 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) in 0.1 M sodium acetate pH 4.5. The ABTS oxidation reaction was followed at 420 nm (ɛ = 36000 M−1cm−1) for 3 min. The MnP activity was determined using 700 μL of the supernatant, 50 μL of 0.2% Phenol red, 50 μL of 25 mM sodium lactate pH 4.5, 50 μL of 2 mM MnSO4, 50 μL of 0.1% bovine albumin, and 50 μL of 2 mM sodium succinate pH 4.5. The absorbance of the MnP reaction mixture was followed for 5 min at 610 nm (ɛ = 4460 M−1cm−1) after adding 50 μL of 100 mM H2O2. The MnP activity reaction was stopped using 40 μL of 2 N NaOH.
In experiments in which glycerol was evaluated as carbon source, a 22 factorial design was established to determine the conditions needed to favor enzyme production, testing inoculum ratios (1:1 and 1:2 of T. maxima/P. carneus) and concentration of glycerol (15 and 20 g/L). All the experiments were done in duplicate, and samples were taken for Lcc and MnP maximum activity at 96 and 144 h, respectively. The STATGRAPHICS Centurion XVI and Design-Expert V 6. 0. 8. were used for statistical analysis of the data. All the data obtained in our experiments are presented as mean ± standard deviation (SD) unless another value is stated in the text. In all the graphics, the sample size value is defined as “n,” and the errors bars represent the SD.
Crude extract entrapment on alginate beads
Crude enzymatic extract was obtained after 15 days of incubation of the mixed culture in the conditions established by the results of factorial design. The crude enzyme precipitation and entrapment methodology is described in the flowchart of Fig. 1. The biomass was separated by means of fine pore filter paper, followed by filtration on syringe filter of 0.22 μm and the addition of protease inhibitor. The protein precipitation was performed on 10 mL of the supernatant solution in a bath of dry ice using ACT which was previously cooled in liquid nitrogen. The precipitation was carried out in a proportion of 83.3% of the ACT. The mixture was shaken for a period of 1 h under sterile conditions. Afterward, it was centrifuged at 4°C for 20 min, and the solvent was removed by decantation. The pellet was dried at ambient temperature during a 30-min period and reconstituted in a 50 mM acetate buffer of pH 4.5 (Harris-Valle et al., 2007). The entrapment of the reconstituted pellet was made in alginate. Three percent (w/v) of the solution (sodium salt of alginic acid) was prepared in a 50 mM acetate buffer at pH 4.5 and mixed with a 2-mL aliquot of the protein precipitate according to Palmieri et al. (1994). The mixture was added dropwise into a solution of 0.15 M CaCl2 in a 10 mM acetate buffer pH 5. The beads were kept at 4°C in the CaCl2 solution for 24 h. Then, the beads were washed with a 0.05 M Tris buffer pH 7 prior crosslinking with a 1% glutaraldehyde solution for 90 min (Bhushan et al., 2015). Finally, the beads were washed with a 50 mM acetate buffer pH 4.5 and freeze-dried after being frozen with liquid nitrogen. The protein, as well as Lcc and MnP enzymatic activities, was measured after freeze-drying in beads surface as described by Palmieri et al. (1994).
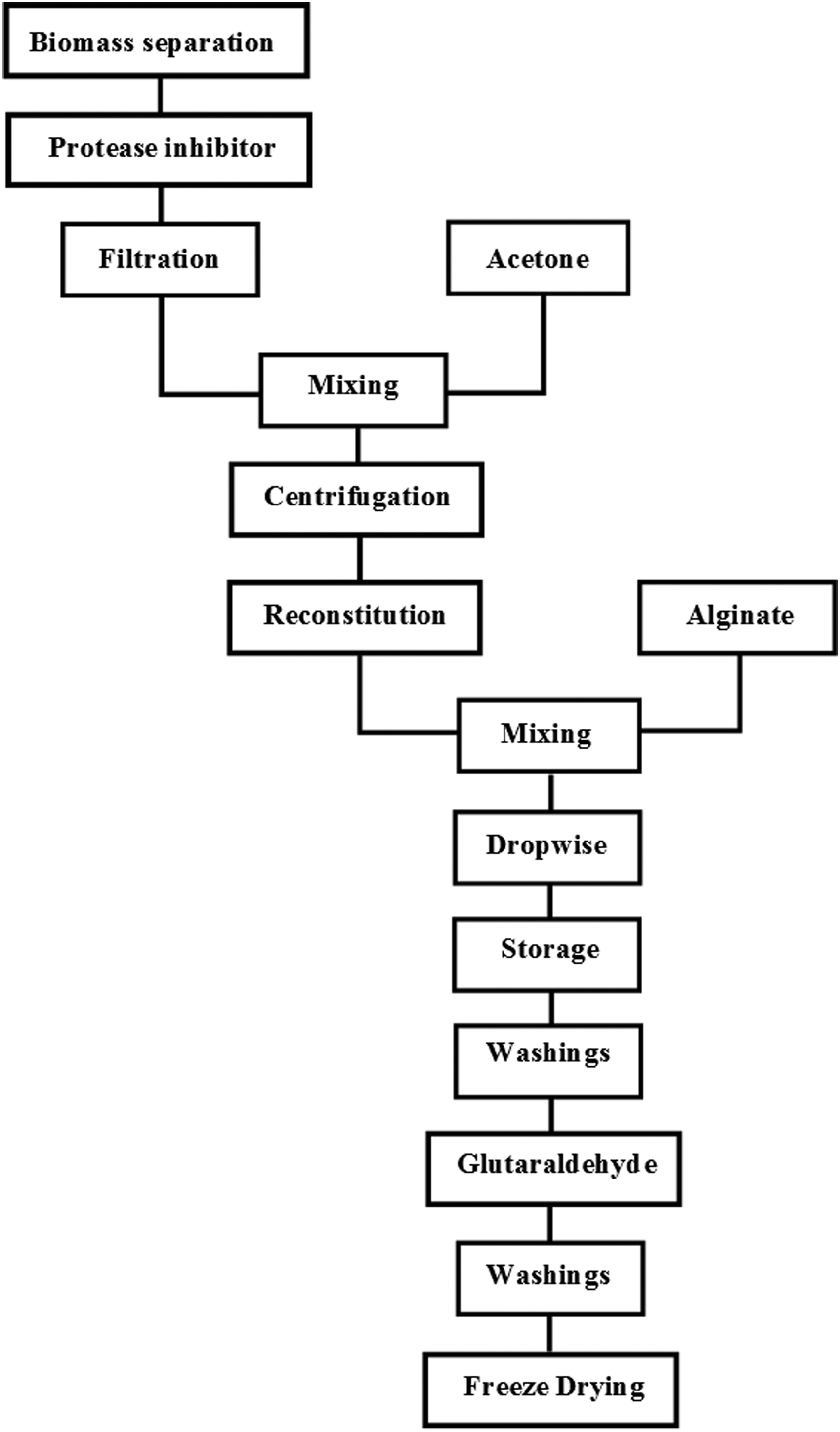
Flowchart of enzyme precipitation and entrapment.
Alginate bead characterization
Material's structure was determined by means of scanning electronic microscopy ([SEM] JEOL JSM-5900LV, 200 kV, at resolution 2.2\AA with an EDS detector). The water content in the alginate beads was calculated by measuring the difference between the wet and dry weights of the materials. The length of the beads was measured using a Vernier caliper. The surface composition was identified using XPS (Al Kα = 1486.6 eV; K-Alpha Thermo Fisher Scientific). The OriginPro 8.1 SR3 v8.1.34.90 software was used for the XPS analysis. Regarding XPS analysis, there is scarce information of the changes in the surface of the removal materials after pollutant treatments. According to McArthur et al. (2014), XPS technique has been mainly used to elucidate the chemical functional groups on the surface of biomaterials.
Removal of PHE by fixed-bed packed column
The system consisted of a glass column, a mixing tank, and an oxygen concentrator as is shown in Fig. 2a and b. The lyophilized alginate beads were packed on a height of 40 mm in a glass column (25 mm of internal diameter and 100 mm in height) to have a length to particle diameter ratio of 21. The packing was made of two layers of 30 mm glass beads separating from 0.43 ± 0.04 g of the alginate beads placed in the middle by a fiberglass mesh. The system was fed with 1 L multiple-pass mixture of 30 mg/L of PHE, 0.01 mg/L of DMSO, and 10 mg/L of Tween 80 dissolved in deionized water using a peristaltic pump with the feed recycle speed of 14.7 mL/min with a contact time of 1.3 min. After 2 h of homogenization (the PHE/DMSO/Tween 80 mixture was circulated through the system), 5 min pulses of 0.125 mL/min of O2 from an oxygen concentrator (MARK 5 Nuvo lite 115 V unit; Nidek Medical Products, Inc.) were administered to the packed column. All the experiments were triplicated, and samples were taken every hour.

Packed column with feed recycled system:
Samples were extracted by means of solid phase extraction cartridges (ENVI-18 Supelco bed wt. 500 mg of 6 mL). The cartridges were conditioned with DCM, ACN, MeOH, and deionized water (10 mL of each one), and the elution was made with 10 mL of DCM. The 0.22 μm filter extract was analyzed by ultra-performance liquid chromatography coupled to mass spectrometry (UPLC-MS technique). The UPLC-MS analysis was carried out by the gradient elution program (the analysis began with 40:60 H2O/ACN, which was followed by a gradient from 10:90 H2O/ACN for 1 min, 40:60 H2O/ACN for 1.1 min and 40:60 H2O/ACN for 2.5 min) with an injection volume of 3 μL and flow rate of 0.5 mL/min at 20°C using a UPLC BEH C18 column (1.7 μm, 2.1 mm × 50 mm) in a Waters UPLC-MS Class H system.
Results and Discussion
Production of crude extract by mixed culture
The glucose consumption in Fig. 3a was complete within 240 h for the mixed cultures in comparison with T. maxima monoculture. There was no statistically significant difference between mono- and mixed culture enzyme production using glucose as the carbon source (least significant difference 95% test, α = 0.05, analysis of variance distribution F, p = 0.04). The higher activities were 7.9 × 104 ± 3.6 × 103 U/g for Lcc and 2.8 × 104 ± 1.2 × 103 U/g for MnP at 120 h in T. maxima monoculture. In the mixed culture, the 1:1 ratio had maximum activities of 7.2 × 104 ± 1.2 × 103 U/g for Lcc at 120 h and 2.4 × 104 ± 1.2 × 103 U/g for MnP at 144 h, whereas the enzymatic activities of the 1:2 ratio were 8.4 × 104 ± 2.6 × 103 U/g for Lcc and 2.4 × 104 ± 1.0 × 103 U/g for MnP at 120 h, as is shown in Fig. 3b and c. The ascomycete P. carneus does not secrete the enzymes Lcc and MnP because the enzymatic tests showed no activity of these enzymes. However, the fungi might compete for carbon source with T. maxima increasing Lcc release as a defense mechanism.

Mixed culture in 20 g/L of glucose:
It has been suggested that carbon depletion leads to higher yields of Lcc expression, which is mainly produced by the secondary metabolism phase. However, the coculture of Ganoderma lucidum and yeast proves that glucose depletion was not correlated with the enhancement of Lcc activity by G. lucidum. The glucose consumption of the yeast predominated producing glycerol as a by-product which instead influenced Lcc activity in G. lucidum (Li et al., 2011). Glycerol has proved to be an efficient carbon source in the production of Lcc and MnP in Cerrena unicolor, in which enzymatic activity was regulated by the carbon sources, being higher with glycerol rather than glucose (Kachlishvili et al., 2014).
In the present work, despite glucose having no difference in enzymatic activity, the use of glycerol as carbon source showed a difference between mono- and mixed cultures despite having a lower Lcc activity in comparison with glucose as observed in Fig. 4a–d. The T. maxima monoculture had a maximum Lcc activity of 3.5 × 104 ± 1.4 × 104 U/g at 96 h and MnP activity of 1.8 × 104 ± 1.0 × 103 U/g at 144 h with 20 g/L of glycerol, whereas in 15 g/L of the carbon source, Lcc and MnP activities were 2.6 × 104 ± 4.2 × 103 at 120 h and 2.4 × 104 ± 2.4 × 103 U/g at 192 h, respectively. In the mixed culture of T. maxima and P. carneus with 15 g/L of glycerol and 1:1 inoculum ratio, the Lcc and MnP activities were 4.9 × 104 ± 2.2 × 102 U/g of Lcc at 96 h and 2.4 × 104 ± 1.6 × 103 U/g of MnP at 144 h. These activities were lower in comparison with the 1:2 ratio for which the values were 6.3 × 104 ± 5.7 × 103 U/g of Lcc at 96 h and 2.8 × 104 ± 1.0 × 103 U/g of MnP at 144 h (Fig. 4a, b). The T. maxima mixed cultures in 20 g/L with a 1:1 inoculum ratio had maximum Lcc and MnP activities of 6.6 × 104 ± 6.1 × 103 U/g at 96 h and 2.4 × 103 ± 2.2 × 102 U/g at 144 h, respectively, while at a 1:2 ratio, the maximum Lcc and MnP activities were 4.6 × 104 ± 6.0 × 103 U/g at 96 h and 1.7 × 104 ± 2.3 × 103 U/g at 144 h, respectively (Fig. 4c, d).

Mixed culture in glycerol:
In the mixed culture of T. maxima and P. carneus, a higher concentration of glycerol as carbon source contributed to increasing the Lcc activity, as well as doubling the ratio of P. carneus in a low carbon source concentration. The latter was determined using the factorial design 22 (Fig. 5a). The 1:2 inoculum ratio and 15 g/L of glycerol enhanced the activity of MnP as observed in Fig. 5b. Equations (3.1) and (3.2) represent the linear regression models obtained for Lcc and MnP activities, respectively.

Factorial Design 22:
The latter indicates that glycerol as a carbon source for the mixed culture allows the regulation of Lcc and MnP secretion in comparison with glucose, in which there was no difference between mono- and mixed cultures. The increase in enzymatic activity as a positive effect of the use of mixed fermentations has been previously described by Netzker et al. (2015). The carbon depletion has been considered a factor that regulates Lcc production in mixed cultures (Hailei et al., 2009). In mixed cultures, the microorganisms compete for access to carbon sources by activating a second metabolism. In the study by Wei et al. (2010) the interaction between Panus rudis and G. W5 strain resulted in an increase of Lcc activity regardless of the carbon source. Yet in the work of Li et al. (2011) glucose addition was detrimental for Lcc production in the coculture of G. lucidum and Candida sp. HSFO7A.
Crude extract entrapment on alginate beads
Based on results obtained from the factorial design 22, 20 g/L of glycerol and 1:1 inoculum ratio of the tested conditions were chosen to obtain the crude extract. The enzymatic activity in Fig. 6 (where the error bars represent the SD) was followed for 408 h, reaching the highest Lcc and MnP activities of 3.4 × 105 ± 1.7 × 104 U/g at 336 h and 1.7 × 104 ± 825 U/g at 288 h, respectively. The precipitate had a protein content of 5.8 ± 1.7 g/L, Lcc activity of 4.9 × 104 ± 1.9 × 104 U/g, and MnP activity of 1.6 × 103 ± 3.3 × 102 U/g before alginate entrapment. The ACT precipitation turned out in purification factors of 0.68, 1.3, and 3.4 for protein, Lcc, and MnP assays, respectively.
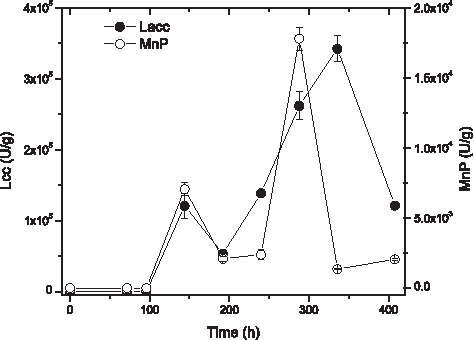
Enzymatic activity of mixed culture in 20 g/L of glycerol: ○ (mean ± SD) MnP and • (mean ± SD) Lcc; n = 2.
Beads of the entrapped crude enzymes observed in the SEM micrograph of Fig. 7a and b showed pore diameters higher than 100 μm. The size distribution histogram in Fig. 7 of the oval-shaped aerogels presented a mean length of 1.9 ± 0.9 mm in their major axis after freeze-drying. The surface of the synthesized alginate beads with entrapped crude enzymes showed values of 0.058 ± 0.003 g/L protein content.

Micrograph SEM at 20 kV of hydrogel:
Removal of PHE by fixed-bed packed column
The packed column with alginate beads without enzymes reached 45.3% ± 23% of PHE removal (total concentration remaining of 10.8 ± 4.7 mg/L of PHE in the water). The beads with enzyme reached 94.3% ± 0.7% (total concentration remaining: 0.83 ± 0.11 mg/L) of PHE removal after 13 h of treatment under nonsterile conditions, as observed in Fig. 8 where the error bars represent the SD. The initial enzymatic activity of alginate beads normalized to total Bradford protein was of 581 ± 73 U/g for Lcc and 126 ± 52 U/g for MnP. The PHE removal by crude enzymes packed in the column was comparable to a TiO2-C-Ag/UVA/photocatalytic treatment which was operated in the same conditions as the packed column (90.9% ± 5.6% of PHE removal), as shown in Fig. 8. The results of the single treatments with irradiation of low energy are described in González-Ramírez et al. (2017).
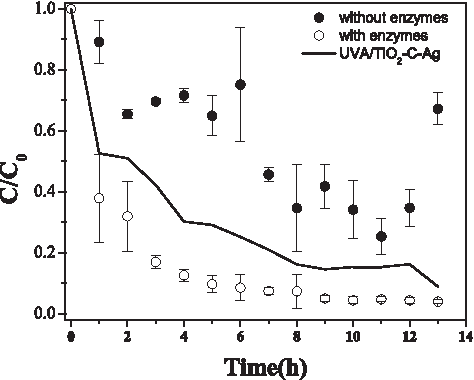
Removal of PHE by alginate beads where: ○ (mean ± SD) with enzymes, • (mean ± SD) without enzymes, and (—) UVA/TiO2-C-Ag treatment; n = 3. PHE, phenanthrene.
PHE removal by the enzymatic crude extract in the alginate beads was also comparable to that of a microbial consortium isolated from contaminated sludge in Shengli oilfields, which was capable of degrading 40 mg/L of PHE at 89% under saline conditions (Shen et al., 2015). The latter indicates that the use of crude enzymes entrapped in alginate is an eco-friendly alternative capable of removing PHE in the presence of dissolved organic matter without needing special conditions or further purification.
The enzymatic cocktails present synergistic effects in which their immobilization allows the development of continuous treatment processes (Asgher et al., 2014). Such enzymatic synergy was identified in this research, making use of XPS surface analysis. The oxidative difference in alginate beads' active surface in PHE removal confirmed by XPS surface analysis is shown in Fig. 9. The XPS analysis was made by the static method with a Gaussian–Lorentzian fitting together with a Shirley-type baseline adjustment and a charge correction to adventitious carbon at 284.8 eV. The adventitious carbon correction is necessary to correct the thin layer of carbonaceous material deposition from the air on the surface of the materials (Greczynski and Hultman, 2017). The data obtained were identified using the NIST database version 4.1 for XPS (NIST, 2012).

XPS C 1s and O 1s regions surface analysis of alginate beads with and without enzymes at 13 and 0 h of treatment. XPS, X-ray photoelectron spectroscopy.
In the XPS spectra (Fig. 9), the differences between the beads with and without crude enzymes were associated with the oxidized species of C and O. The O 1s region presented an increase of C—O and C═O groups in comparison with the beginning of treatment. Fei et al. (2016) described the incorporation of graphene oxide in which there was an addition of C═C and C═O groups. In the present work, the PHE deposited on the surface of alginate beads with enzymes resulted in an abundant integration of C═O species in the hydrogel after PHE treatment. The latter XPS results gave evidence of oxidation of the PHE/DMSO/Tween 80 mixture on the alginate beads by the surface enzymatic activity. However, except for Lcc and MnP, the rest of the enzymes responsible for the PHE removal remain unknown due to the use of a crude extract.
In addition, the use of Tween 80 might benefit the PHE removal process due to the surfactant content of unsaturated fatty acid esters. The oxidative degradation of PHE by Phanerochaete chrysosporium has been related to the simultaneous action of its extracellular ligninolytic enzymes, specifically, lipid peroxidation by MnP and unsaturated fatty acid esters of Tween 80 in the presence of O2 (Sutherland et al., 1991; Hammel et al., 1992; Moen and Hammel, 1994). The presence of O2 in lipid peroxidation by MnP has been described by Watanabe et al. (2000) as necessary to supply H2O2 by deprotonation of
In contrast to MnP, Lcc requires the addition of oxidizable low-molecular weight compounds and binding oxygen to reach the oxidation of nonsubstrate compounds such as PHE (Solomon et al., 2001; Cantarella et al., 2003; Han et al., 2004). The oxygen-centered radicals' product of lipid peroxidation generates co-oxidation of aromatic compounds; in the case of PHE, electron abstraction by lipid oxyradicals is the main route associated with oxidation (Moen and Hammel, 1994). In our system, Tween 80 in combination with the addition of O2 pulses into the system might activate the enzymes on the surface of the alginate beads generating oxyradical reactions which could be responsible for PHE removal. In the mineralization of PHE, Sack et al. (1997) found the presence of thiol groups from glutathione necessary to activate radical generation from MnP crude enzyme. The presence of organic or metal cofactors in combination with oxygen (present in the ground state having low reactivity with organic molecules) allows the formation of substrate radicals that can transfer electrons to oxygen contributing to oxyradical production (Bugg, 2014; Soi and Steiner, 2016). Moreover, the presence of DMSO as a cosolvent of Lcc enhances the solubility of substrates (Mogharabi and Faramarzi, 2014).
On the basis of the above, the PHE/DMSO/Tween 80 mixture might increase the contact between PHE and the crude enzymes. In the photocatalytic treatments in which DMSO or Tween 80 was not used, PHE precipitated and no removal was observed. In the research of Eibes et al. (2006), the presence of ACT enhanced the solubility of PAHs improving the degradation of PAH by MnP. In the absence of ACT, the removal of the PAHs was associated with the loss of solubility and reduction of the ACT by H2O2. In the case of the PHE removal by alginate beads without enzymes, PHE deposition was associated with an increase of carbonyl groups. The absence of sulfur groups related to DMSO on the beads' surface could be the product of reduction of DMSO by oxyradicals (especially OH) to by-products which are water miscible (Scaduto, 1995). The decrease of DMSO lowered PHE solubility and increased PHE removal by deposition on alginate beads.
Conclusions
In T. maxima and P. carneus mixed culture, the source of carbon had the highest effect on Lcc and MnP secretion rather than carbon depletion. Twenty gram per liter of glycerol and 1:1 inoculum ratio were the conditions which significantly enhanced Lcc (6.6 × 104 ± 6.1 × 103 U/g) and MnP (2.4 × 103 ± 2.2 × 102 U/g) secretion in the mixed culture. The enzymatic crude extract entrapped in the alginate beads removed 94.3% ± 0.7% of PHE after 13 h of treatment. The results indicate that the entrapped crude enzyme treatment had a superior PHE removal than single TiO2-C-Ag/UVA/photocatalytic treatment.
Despite the low purification factors in the precipitation, the crude enzymatic extract entrapped in alginate beads showed an oxidative active surface in PHE removal, which was confirmed by XPS surface analysis of the beads. The XPS technique enabled the identification of functional groups on the surfaces of the alginate beads after PHE treatment. Oxyradical reactions produced on the surface of alginate beads with enzymes could be responsible for an important increase of C═O groups. However, the mechanism by which PHE removal was achieved requires further research. The addition of Tween 80 and DMSO in PHE aqueous solution benefited the enzymatic activity of the crude extract from the mixed culture of T. maxima and P. carneus without needing special conditions or further purification. Such results indicate that the use of crude enzymes entrapped on alginate is an eco-friendly alternative capable of removing PHE in the presence of Tween 80 and DMSO. In the application of crude enzymatic extracts for wastewater treatments, the presence of anthropogenic surfactants and organic solvents might favor PAH removal. The future development of such materials might be an important alternative in the treatment of real wastewaters as the PHE removal was comparable with the one achieved in a photocatalytic treatment.
Footnotes
Acknowledgments
The authors thank the National Institute for Nuclear Research (ININ), CINVESTAV-IPN, and CONACYT (Grant No. 355106) for the support provided through the development of this research. The study was supported by Consejo Nacional de Ciencia y Tecnología (Grant No. 355106).
Availability of Data and Materials
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
Author Disclosure Statement
None of the authors in this article have a financial or personal relationship with other people or organizations that could inappropriately influence or bias the content of the article.