
Abstract
Abstract
Effects of moderate-intensity static magnetic field (SMF) on biodecolorization of azo dyes by a marine microbial community (MMC) and halotolerance of the consortium were investigated. Performance of decolorization, chemical oxygen demand (COD) removal, detoxification, and biomass multiplication exposed to SMF of different intensities were monitored and compared. In addition, effects of SMF on microbial community structure were analyzed using a high-throughput sequencing technique. Results showed that compared with those with the MMC without SMF, efficiencies of decolorization, COD removal, and detoxification were higher when exposed to 45.3 mT SMF; however, they were lower when exposed to 95.0 and 249.5 mT SMF. By contrast, biomass multiplication was enhanced by all the three tested SMFs. On the contrary, 45.3 mT SMF also enhanced halotolerance of the MMC. Results of MiSeq sequencing demonstrated that 45.3 mT SMF influenced biodiversity and structure of the MMC, and the relative abundance of some putative genera involved in azo dye decolorization increased, including bacteria such as Shewanella and Vibrio, fungi such as Issatchenkia, and genera affiliated to unclassified Pseudeurotiaceae. This study might provide a potentially useful pathway for improving effectiveness of biological treatment processes and their tolerance to extreme conditions in practical applications.
Introduction
A
Biological processes are attractive for treating wastewaters because they are low cost, effective, and environmentally friendly (Tan et al., 2016). However, biological methods are sometimes unstable and inefficient when treating recalcitrant industrial wastes because of their low biodegradability. It is proven that static magnetic field (SMF) can improve effectiveness of biological treatments through influencing biomass metabolism, enzyme activity, cell membrane permeability, etc. (Kovacs et al., 1997; Liu et al., 2008). In general, SMF is classified as weak (<1 mT), moderate (1 mT–1 T), strong (1–5 T), and ultrastrong (>5 T) (Filipič et al., 2012). Among them, strong SMFs generally inhibit physiological processes of organisms; by contrast, nonstrong (especially moderate) SMFs always show positive effects (Miyakoshi, 2005). Thus, most of the pertinent research focuses on moderate SMFs.
On one hand, moderate-intensity SMF could influence multiplication of microorganisms. Křiklavová et al. (2014) reported that a 370 mT SMF promoted the growth rate of Rhodococcus erythropolis by around 28% through shortening its lag and exponential phases. Liu et al. (2008) also found that a 75 mT SMF could promote the multiplication rate of an anammox community by 50%. Furthermore, although diversity of the anammox community decreased after being exposed to SMF, planctomycetes-related organisms were highly enriched under long-term magnetic-exposed conditions, thus promoting the ammonium oxidizing activity. However, moderate-intensity SMF sometimes displays obscure or even negative effects on some bacteria.
For instance, Potenza et al. (2004) indicated that 300 mT SMF showed no influence on growth of Escherichia coli in nutrient-rich Luria Bertani (LB). Ji et al. (2010) showed that 450 mT SMF inhibited growth of E. coli and the inhibitory effect increased with temperature. Meanwhile, as described by Morrow et al. (2007), 50–500 mT SMF showed variable effects on the growth of Streptococcus pyogenes. Its growth rate was inhibited when exposed to 300 mT SMF; however, its growth rate was promoted by 500 mT SMF. It is suggested that moderate-intensity SMFs can influence growth and survival of bacteria; however, comprehensive magnetic biological effects and the corresponding mechanisms are still not completely understood (Hughes et al., 2005).
On the other hand, moderate-intensity SMFs can also promote biodegradation efficiencies of organic pollutants. Łebkowska et al. (2013) found that removal efficiency of 1600 mg/L formaldehyde by activated sludge was promoted by 20% through being exposed to 7 mT SMF. Křiklavová et al. (2014) reported that phenol removal efficiency by Rhodococcus erythropolis was promoted by short-term repeated exposure to 370 mT SMF. Similar results were also described in many other literature (Tomska and Wolny, 2008; Ji et al., 2010; Wang et al., 2012a; Łebkowska et al., 2013).
Although improving effectiveness of microorganisms by adding SMF is attracting more and more attention, it is obvious that most of the corresponding research aims at pure cultures. As is well known, complicated microbial communities are always more adaptable to complex environments than pure microorganisms, thus can keep more stable treatment efficiency for field application. Meanwhile, study of microbial community composition can contribute to understanding of biotreatment processes (Forss et al., 2013). Therefore, systematic research based on microbial communities, especially on dynamics of community structure, is still in demand.
Wastewaters from petroleum, textile, and leather industries always contain high concentrations of salts, which can cause plasmolysis of microorganisms and thus reduce biological activity (Castillo-Carvajal et al., 2014). Application of halophilic or halotolerant microorganisms can overcome these limitations; however, many of them are also inhibited by excessive salinity (Zhuang et al., 2010). Turk et al. (2004) indicated that high salinity would induce alterations in membrane lipid patterns of microorganisms, which might be related to halotolerance. Meanwhile, Niu et al. (2013, 2014) reported that SMF could promote microorganisms to adapt to low temperature through producing more unsaturated fatty acids in plasma membrane. It is speculated that SMF may also stimulate regulation of membrane lipid composition, thus promoting halotolerance of microorganisms. As is known, there has been no report on promoting microbial halotolerance by SMF.
The aim of this study was to estimate the enhancing effect of SMF on marine microbial community (MMC) for treating azo dyes under hypersaline conditions. Effects of SMF of different intensities on decolorization, chemical oxygen demand (COD) removal, and detoxification of azo dyes by MMC, as well as biomass multiplication and tolerance to hypersaline conditions were investigated through sequencing batch tests. In addition, the MiSeq sequencing method was used to analyze and compare structures of microbial communities with and without SMF of optimal intensity, to illustrate magnetic biological effect on the MMC.
Experimental Protocols
Reagents
Six azo dyes including Acid Red B (ARB), Reactive Green KE-4BD (KE-4BD), Acid Orange II (AOII), Reactive Brilliant Red K-2G (K-2G), Acid Scarlet 3R (3R), and Reactive Violet KN-4R (KN-4R) with the characteristic absorption wavelengths of 516, 627, 507, 509, 484, and 545 nm, respectively, were purchased from Dye Synthesize Laboratory, Dalian University of Technology (Dalian, Liaoning Province, China) with the purity of >99%. Other chemical reagents (analytical grade) were purchased from J&K Chemical Ltd. (Shanghai, China). Biochemical reagents were purchased from Sangon Biotech Co., Ltd. (Shanghai, China).
MMC and culture mediums
The MMC (sea mud samples) used in this study were gathered from the beach of Heishijiao Park (38.88°N, 121.57°E), Dalian, China. Before acclimatization and inoculation, the sea mud was pretreated through filtration with a 100-mesh screen. Acclimatization of the MMC and the following decolorization tests were performed in the media containing (g/L): glucose 2.0, (NH4)2SO4 0.5, K2HPO4 0.1, MgSO4 · 7H2O 0.5, NaCl 30.0, and ARB 0.05–0.30 (pH was adjusted to about 6.0). The medium was prepared just before use without sterilization. The sea mud was cultured in shaking flasks at rotation speed of 160 r/min and temperature of 30°C.
Decolorization experiments
Decolorization experiments were performed in 250 mL shaking flasks with 100 mL culture medium containing glucose 2.0, (NH4)2SO4 0.5, K2HPO4 0.1, MgSO4 · 7H2O 0.5, NaCl 30.0, and azo dye 0.30. Three tested SMFs were supplied by circular permanent magnets (100 mm in diameter and 0.5–17.5 mm in thickness), the average intensities of which were 45.3, 95.0, and 249.5 mT. Experiments were performed in the form of sequencing batch tests (12 h for each batch, eight batches for each group of test). At the end of each batch, the sea mud (MMC) was separated from the mixture through centrifugation at 10,000 g for 10 min, and then returned back to the flasks with fresh culture medium for the next batch. Meanwhile, the blank control set inoculated with autoclaved sea mud was used to investigate the manner of decolorization through adsorption.
Decolorization, COD removal, (acute toxicity) detoxification efficiencies, and biomass multiplication were monitored for investigating the effect of SMF on bioprocess with MMC. Furthermore, high-throughput sequencing techniques were used for monitoring difference between the community structure of the MMC with and without SMF.
Assays
Concentration of azo dyes was analyzed using spectrophotometry (JASCO V-560; Japan). Dye solution was pretreated through centrifugation (10,000 g for 10 min) before analysis. Decolorization ratio was defined as (A0 − A1)/A0 × 100, where A0 and A1 were the absorbance of azo dyes before and after decolorization, respectively. COD was also monitored through the method described by Shi et al. (2015).
Acute toxicity of azo dyes before and after decolorization was assessed using Microtox method with luminescent bacteria Vibrio fischeri (NRRL B-11177) as described by Tan et al. (2016). Acute toxicity was represented by the inhibition ratio (IR) of luminescence by IR (%) = (S0 × Cf − St)/(S0 × Cf), where S0 and St were the luminescence intensities of the samples at 0 and t min, respectively; Cf was the relative luminescence intensity variations of negative sample at 0 – t min. Detoxification percentages were calculated by detoxification (%) = (IR0 − IRf)/IR0 × 100%, where IR0 and IRf were the IRs of the dye solution before and after decolorization. In addition, to eliminate the interference of abiotic solids in the sea mud, concentration of microbes was represented by mixed liquor volatile suspended solids (MLVSSs), which was measured through the method described by Hu et al. (2017).
All the experiments were carried out in triplicate.
DNA extraction, polymerase chain reaction amplification, and MiSeq sequencing
The MMCs after treatment without and with SMF of the optimal intensity, as well as the original sea mud, were gathered and conserved at −80°C for further analysis. Genomic DNA was first extracted, purified, and quantified according to the method described by Tan et al. (2013). Then, the DNA samples were amplified through polymerase chain reaction (PCR) with the primers of 515F/907R and 1737F/2043R for bacterial and fungal communities, respectively (White et al., 1990; Xiong et al., 2012). PCR products were purified and analyzed by Sangon Biotech Co., Ltd. (Shanghai, China) with an Illumina MiSeq platform (Illumina). Statistical and bioinformatics analyses of sequencing results were performed on the basis of previous procedures (Li et al., 2015).
Statistical analysis
Statistic analysis of experimental data was performed through one-way analysis of variance using Microsoft Excel 2010 software. The p-value of <0.05 suggests that data were considered significant.
Results
Biodecolorization of azo dyes by the MMC with SMF of different intensities
To investigate azo dye (ARB) removal through abiotic processes (adsorption) by the MMC, a group of experiments were performed in advance with sterilized sea mud as blank control. As shown in Figure 1A, color removal percentage was >40% during the first batch of test. However, just <12% of ARB was decolorized during the second batch of treatment. After treatment for four batches, color removal percentages were stabilized at <5%. Meanwhile, COD removal and detoxification percentages were also relatively high during the first batch; however, they dramatically decreased from the second batch (Fig. 1B, C). These results suggested that although part of color, COD, and acute toxicity were removed through adsorption at the beginning, the interference could be neglected after two or three batches when solid surface of the sea mud was saturated by azo dye and other organics.

Blank control experiments with autoclaved sea mud:
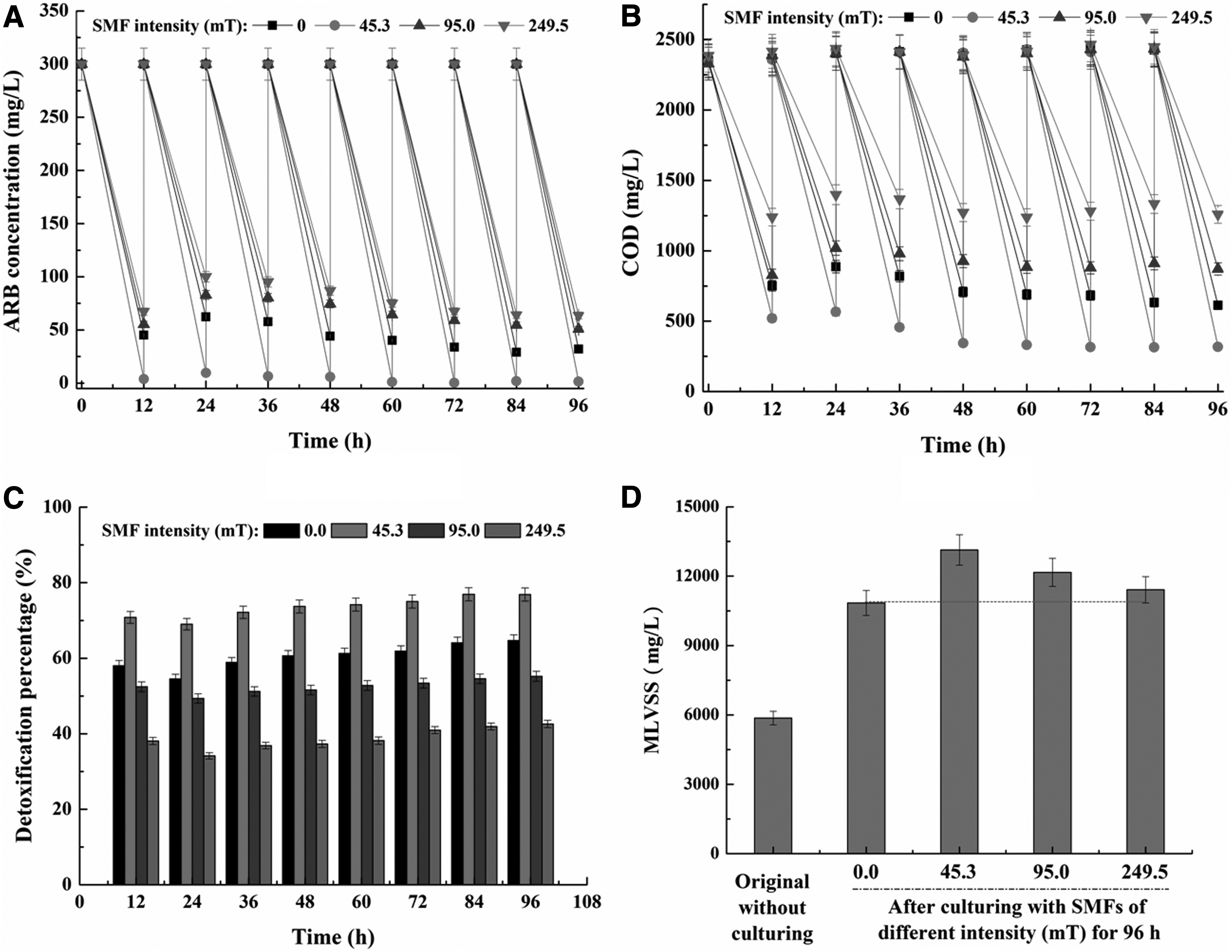
The result of ARB decolorization by the MMC with SMF of different intensities is shown in Figure 2A. Less than 85.0% and 79.2% of 300 mg/L ARB was decolorized (corresponding to effluent ARB concentrations of 45.2 and 62.3 mg/L) by the MMC during the first and second batches of treatment in the absence of SMF, respectively. In comparison, 98.7% and 96.8% of ARB were decolorized (2.3 and 3.1 mg/L ARB in effluent) by the MMC when exposed to 45.3 mT SMF during the first two batches of treatment, which were 16.1% and 22.2% higher than the corresponding decolorization percentages without SMF. On the contrary, the corresponding decolorization percentages by the MMC with 95.0 and 249.5 mT SMFs were 81.6%/72.4% and 77.6%/66.7%, which were 4.0%/8.6% and 8.7%/15.8% lower than those without SMF, respectively.
Effects of SMF intensity on performance of the MMC:
Moreover, decolorization percentages of all the four groups slightly increased with time because of acclimatization. At the end of the eighth batch of treatment, decolorization percentages were 89.4%, 99.5%, 83.0%, and 78.8% corresponding to the SMF intensities of 0, 45.3, 95.0, and 249.5 mT, respectively. It was obvious that decolorization percentage with 45.3 mT SMF was the highest among the four groups, followed by that without SMF and then those with 95.0 and 249.5 mT SMFs. These results suggested that SMF of certain intensity could improve ARB decolorization by the MMC; however, excessively strong SMF would adversely affect the bioprocess.
Meanwhile, as shown in Figure 2B, COD removal efficiency of all the four sets was also relatively low at the beginning and then gradually increased with time. The final effluent COD was 613.8 mg/L with the influent concentration of 2426.8 mg/L (74.7% removal percentage) in the absence of SMF. In comparison, the corresponding effluent COD was 317.9 mg/L (86.9% removal percentage) with 45.3 mT SMF, which was 48.2% lower (16.3% higher corresponding to the removal percentage) than that without SMF. However, when SMF intensity increased to 95.0 and 249.5 mT, effluent COD levels were 871.1 and 1258.8 mg/L (corresponding to 64.1% and 48.6% removal percentages), respectively, which were both higher (lower corresponding to removal percentages) than that without SMF. It was obvious that 45.3 mT SMF also promoted COD removal effectiveness by the MMC.
In addition, as shown in Figure 2C, 76.9% of acute toxicity was removed by the MMC when exposed to 45.3 mT SMF during the eighth batch of treatment; in comparison, the corresponding toxicities with 0, 95.0, and 249.5 mT SMFs were 64.7%, 55.2%, and 42.6%, which were 15.9%, 28.2%, and 44.6% lower than that with 45.3 mT SMF, respectively. It was suggested that ARB and its toxic by-products were further degraded when exposed to 45.3 mT SMF, thus resulting in higher detoxification percentages than the other three groups.
On the contrary, biomass represented by MLVSS before and after treatment for 96 h (eight batches) with different SMF intensities is shown in Figure 2D. MLVSS of the group without SMF increased from 5.86 to 10.84 g/L after treatment for eight batches. Meanwhile, the final MLVSSs were 13.13, 12.16, and 11.41 g/L with 45.3, 95.0, and 249.5 mT SMFs, which were all higher than that without SMF up to 21.13%, 12.18%, and 5.26%, respectively. It was obvious that biomass multiplication of the MMC was higher with all the tested SMFs than that without SMF. Meanwhile, among the three groups exposed to SMF of different intensities, the group with 45.3 mT SMF showed the fastest biomass multiplication rate.
Treatment of other five azo dyes by the MMC with and without 45.3 mT SMF was also investigated and the results are given in Table 1. It was obvious that decolorization efficiencies of the other five azo dyes were all higher with 45.3 mT SMF than those without SMF up to 18.3–40.6%. The corresponding COD removal and detoxification effectiveness were also improved by the 45.3 mT SMF up to 23.0–33.6% and 36.8–65.0%, respectively. Therefore, it was suggested that biodegradation of azo dyes besides ARB by the MMC could also be improved by the 45.3 mT SMF.
decolorization, COD removal, and detoxification percentages of the first and eighth (the last) runs.
1, system inoculated with autoclaved sea mud as blank control; 2, system inoculated with MMC in absence of SMF; 3, system inoculated with MMC in the presence of 45.3 mT SMF.
SMF, static magnetic field; MMC, marine microbial community; COD, chemical oxygen demand.
Halotolerance of the MMC with and without SMF
As shown in Figure 3A, 94.6–98.9% of 300 mg/L ARB (resulting in 3.1–23.7 mg/L ARB in effluent) was decolorized by the MMC with 45.3 mT SMF and 0 g/L NaCl throughout the whole treatment process. When salinity increased to 30 g/L NaCl, the corresponding decolorization percentages ranged from 92.1% to 98.9%. When salinity further increased to 50 g/L NaCl, decolorization percentages decreased to 68.7. In comparison, 79.9–97.1% of 300 mg/L ARB was decolorized by the MMC in the absence of SMF with 0 g/L NaCl, which was 1.8–15.5% lower than that with 45.3 mT SMF. When NaCl concentrations increased to 30 and 50 g/L, the corresponding decolorization percentages were 77.2–90.3% and 49.1–58.8%, respectively, which were 8.7–16.2% and 28.2–28.5% lower than those with 45.3 mT SMF.

Effects of 45.3 mT SMF on performance of the MMC with different salinities:
As shown in Figure 3B, COD removal percentages were 77.8–88.3%, 73.7–87.3%, and 57.3–60.3% with 0, 30, and 50 g/L NaCl when exposed to 45.3 mT SMF, respectively. In comparison, the corresponding COD removal percentages were 72.5–78.9%, 58.1–69.3%, and 38.9–50.6% in the absence of SMF, which were also lower than the corresponding percentages with 45.3 mT SMF.
Similarly, detoxification efficiencies were also higher with 45.3 mT SMF than those in the absence of SMF under the same saline conditions, as shown in Figure 3C. It could be concluded that decolorization, COD removal, and detoxification efficiencies with 45.3 mT SMF were always higher than those in the absence of SMF under a wide range of saline conditions.
In addition, MLVSSs after the eight-batch operation with 45.3 mT SMF were always higher than the corresponding MLVSSs without SMF, as shown in Figure 3D. Although growth of MMC was inhibited with 50 g/L NaCl, the final MLVSS with 45.3 mT (3907.1 mg/L) was still 252.7% higher than that without SMF (1107.8 mg/L). It was suggested that 45.3 mT SMF also promoted biomass multiplication of the MMC with 0–50 NaCl. Therefore, it could be concluded that 45.3 mT SMF enhanced halotolerance of the MMC.
Meanwhile, the mentioned results also suggested that growth and metabolism activities of MMC were higher in lower salt environments. As already mentioned, halophilic or halotolerant microorganisms could keep relatively high biodegradation efficiency under hypersaline conditions. It was shown that halotolerant microorganisms could tolerate hypersaline conditions; however, they were generally inhibited by high salinity (Tan et al., 2016). In comparison, halophilic microorganisms always grew and metabolized obligately with high concentration of salt (Oren, 2010). Thus, the possible reason was that dominant species in MMC after treatment were mainly halotolerant microorganisms, which would be further validated through microbial community analysis.
Effects of SMF on microbial community structure
It was indicated that bacteria were the main components of a wastewater treatment microbial system such as activated sludge (Seviour and Nielsen, 2010). Meanwhile, fungi were also important for biological treatment (especially for recalcitrant organics) because of their nonspecific degrading enzymes (Li et al., 2015; Tan et al., 2016). Thus, the structure of bacterial and fungal communities before and after treatment with 0 and 45.3 mT SMFs was analyzed with the MiSeq sequencing method and the results are shown in Figures 4–6 and Tables 2 and 3. More than 58,772 and 44,047 sequences for bacterial and fungal communities were yielded after preliminary removal of short and low-quality reads, respectively.
Rarefaction curves of
Relative abundance of bacteria at
Relative abundance of fungi at
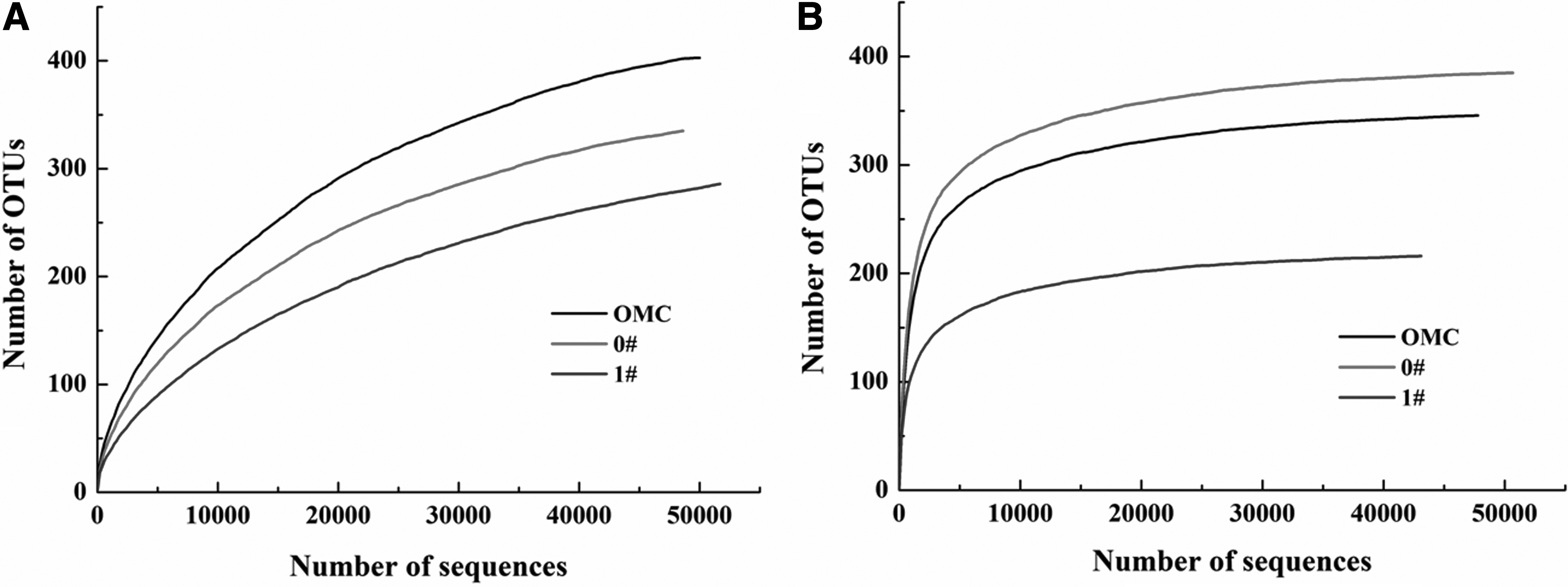
Rarefaction curves shown in Figure 4, which were drawn based on sequencing results, indicated that richness of fungal community was complete; however, that of bacterial community was not (Wang et al., 2012b).
Richness and evenness of microbial communities could be represented by the Shannon's diversity index (Ma et al., 2013). As given in Table 2, the Shannon's diversity index of bacterial species in the original MMC was higher than that of the other two. The Shannon's diversity index decreased from 2.58 before treatment to 2.25 and 1.93 after treatment without and with SMF, respectively, suggesting that the 45.3 mT SMF adversely affected the richness and evenness of bacterial community. On the contrary, the Shannon's diversity index given in Table 3 indicated that in comparison with the original MMC, the richness and evenness of fungal species increased after treatment without SMF; however, they obviously decreased after treatment with 45.3 mT SMF. Meanwhile, the Chao1 index given in Tables 2 and 3 for estimating total operational taxonomic unit (OTU) numbers supported the results already described.
It was suggested that impacts of SMF on bacterial and fungal community diversity were different. The reason was that SMF could improve effectiveness of biological treatments through influencing biomass metabolism, enzyme activity, and cell membrane permeability as already mentioned (Kovacs et al., 1997; Liu et al., 2008), which were different between bacteria and fungi. For instance, plasma membrane fatty acid unsaturation of bacteria (e.g., Aeromonas species) decreased with high salinity (Chihib et al., 2005); however, that of fungi (e.g., Debaryomyces hansenii) increased (Turk et al., 2007), suggesting that their regulation mechanisms under hypersaline conditions were different. Therefore, impacts of SMF on bacterial and fungal communities were different.
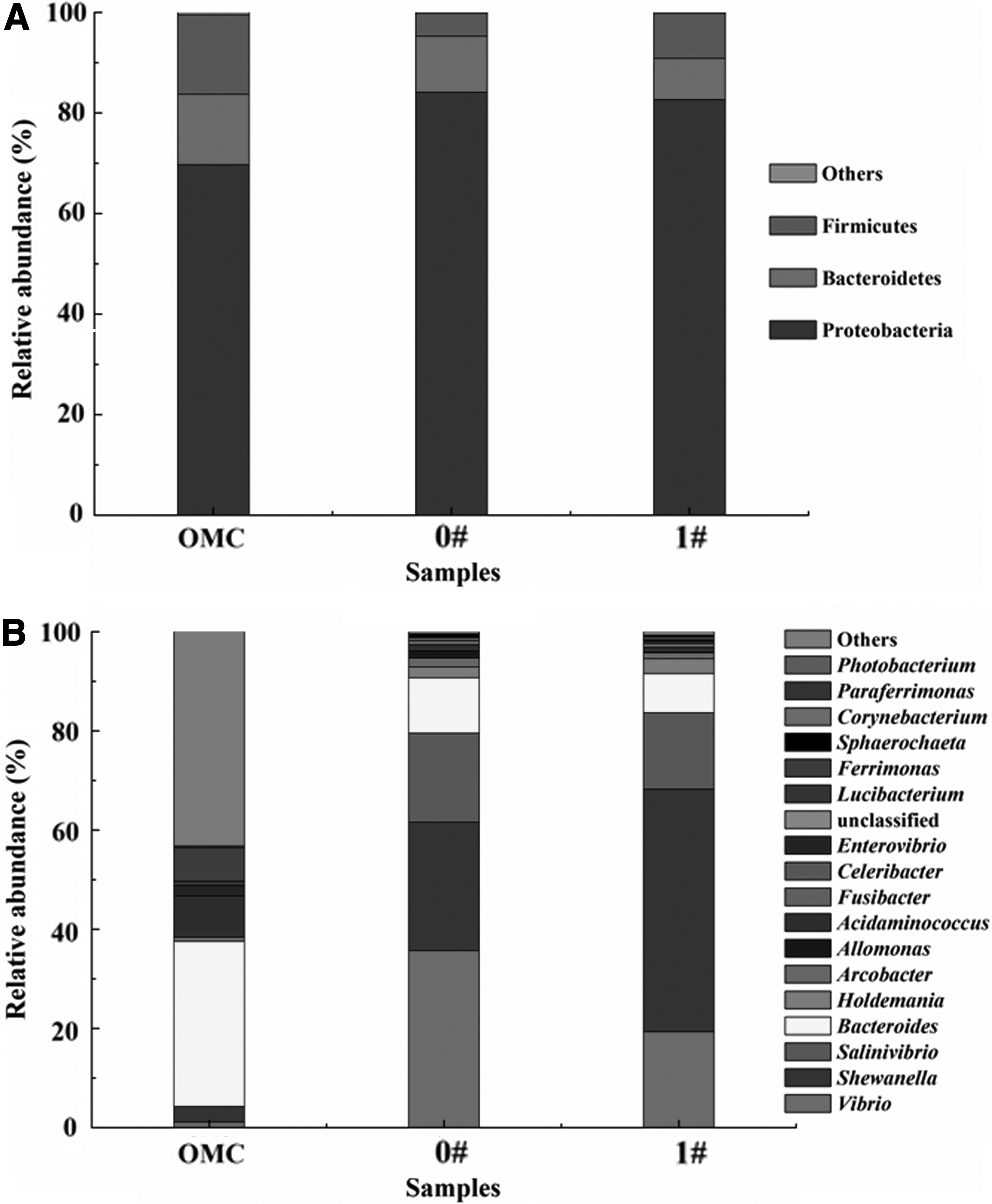
The relative abundance of bacteria at phylum and genus levels is displayed in Figure 5. It was shown that dominant bacteria in all the three microbial communities mainly belonged to three phyla, among which the relative richness of Proteobacteria was the highest, followed by Bacteroidetes and Firmicutes. In addition, <0.5% of dominant bacteria were affiliated to other phylum. The relative abundance of Proteobacteria increased from 69.7% in original MMC to 84.1% and 82.7% after treatment without and with 45.3 mT SMF (0# and 1#), respectively. However, the relative abundance of Bacteroidetes and Firmicutes decreased after treatment.
At the genus level, the relative abundance of Bacteroides, Ferrimonas, and Acidaminococcus was lower in 0# and 1# samples than that in the original MMC. In comparison, some other genera such as Salinivibrio, Shewanella, and Vibrio were relatively enriched after treatment. Furthermore, the relative abundance of Shewanella was obviously higher in 1# sample than in 0# sample. By contrast, the relative abundance of Bacteroides, Salinivibrio, and Vibrio was lower in 1# sample than in 0# sample.
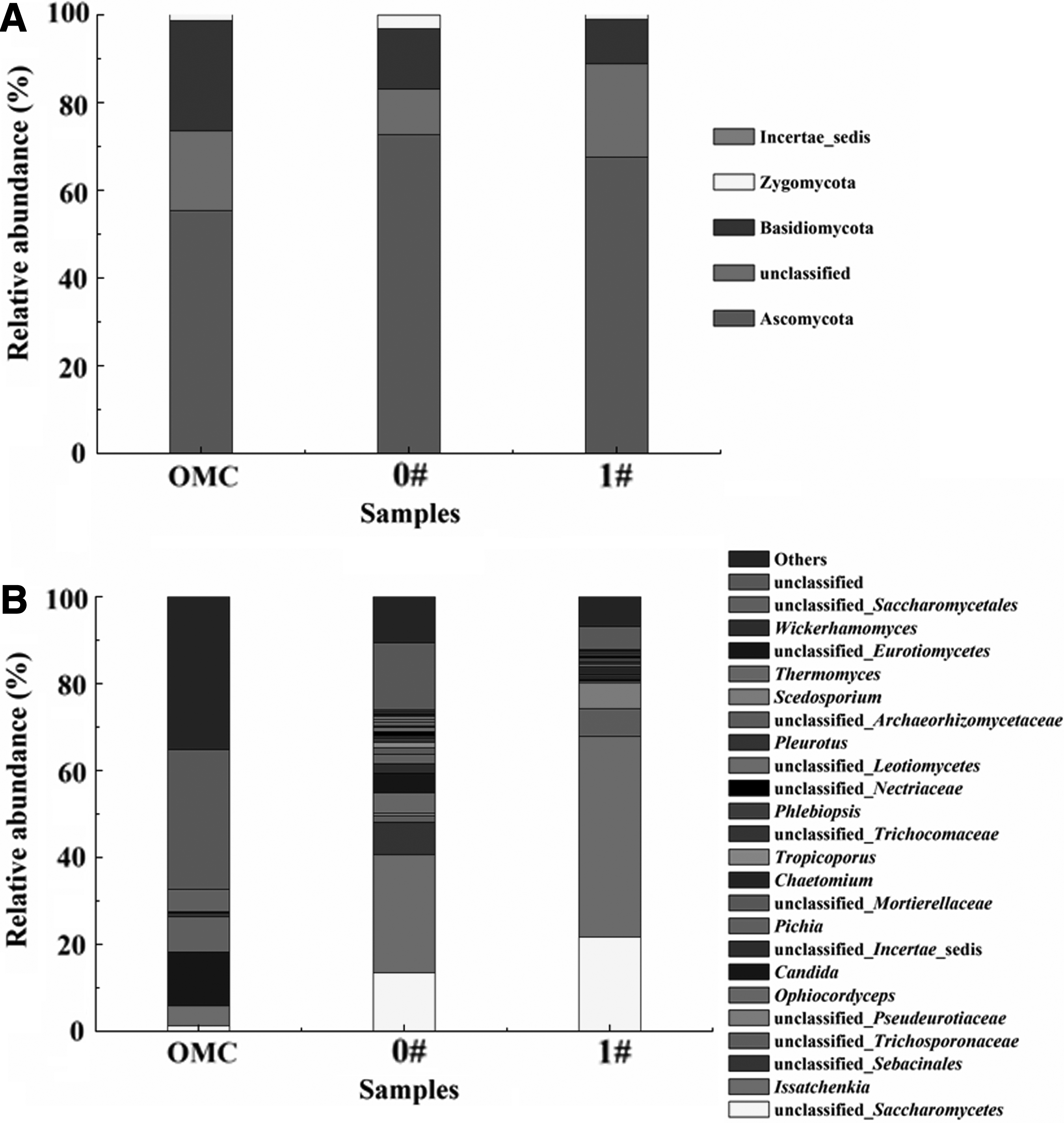
On the other hand, the relative abundance of fungi at phylum and genus levels is shown in Figure 6. Ascomycota, Basidiomycota, and some unclassified phyla were dominant fungal phyla. The relative abundance of Ascomycota increased from 55.4% in the original MMC to 72.7% and 67.6% in 0# and 1# samples, respectively. By contrast, the relative richness of Basidiomycota was lower in 0# and 1# samples than in the original MMC.
At the genus level, the relative abundance of Issatchenkia and some genera affiliated to unclassified Saccharomycetes increased from 4.6% and 1.3% before treatment to 27.2% and 13.5% after treatment, respectively, without SMF. Meanwhile, the corresponding relative richness of these two phylotypes was 46.2% and 21.6% after treatment with 45.3 mT SMF, respectively, which were higher than those without SMF. In addition, growth of some fungi affiliated to unclassified Pseudeurotiaceae and Trichosporonaceae was also stimulated by SMF. By contrast, the relative abundance of Candida, Pichia, and some unclassified genera decreased after treatment in the absence of SMF, then the relative abundance further decreased with SMF.
Discussion
Many reports indicated that biological treatment processes could be enhanced by SMF. Higher removal efficiency of organics (COD) or target compounds (e.g., phenol and ammonium) by single microorganisms or microbial consortia was observed when exposed to SMF of different intensities (Liu et al., 2008; Ji et al., 2010; Křiklavová et al., 2014). Meanwhile, acceleration of biomass multiplication was also observed in the above studies. It seems that promotion of biological efficiency by SMF related to the enhancement of biomass multiplication rate, which was also concluded by Santos et al. (2010) and Small et al. (2012).
In this study, SMF of different intensities showed entirely different promotion (45.3 mT) and inhibition (95.0 and 249.5 mT) effects on biotreatment efficiency; however, exposure to SMF of all the three tested intensities resulted in higher biomass multiplication than that in the absence of SMF. It was suggested that effects of SMF on microbial metabolic activity might sometimes not be only related to biomass. As reported by Niu et al. (2013) and Li et al. (2011), SMF could also increase enzymatic activity of microorganisms, thus causing enhancement of biodegradation efficiency.
Mechanism of magnetic effect on enzymatic reaction kinetics might be that diffusion resistance could be reduced by magnetic field so that enzymes approach and recombine with substrate at a faster rate (Kovacs et al., 1997). Another possible mechanism explained by Jung and Sofer (1997) was that more extracellular enzymes would distribute on the surface of bacteria when exposed to SMF, thus organics could be degraded extracellularly rather than being transferred into cells. In addition, it was indicated that effects of moderate/low-density SMF on eukaryotic microbes are generally associated with changes in Ca2+ influx and permeability of membrane ion channels (Rosen, 2003). Therefore, it was suggested that biotreatment efficiency might be improved because of the increase of biomass multiplication, enzymatic activity, or both of them.
In this study, improvement of biodegradation efficiency by 45.3 mT SMF was apparently related to enhancement of biomass multiplication, which could be estimated by the approximate promoting ratios (22.2% for decolorization, 16.3% for COD removal, 18.9% for detoxification, and 21.1% for MLVSS, higher than the corresponding index without SMF). However, although 95.0 and 249.5 mT SMFs also promoted biomass multiplication, they displayed negative effect on biodegradation. This result suggested that enzymatic activity of the MMC was also influenced by SMF.
It was previously reported that SMFs could also improve tolerance to extreme conditions of microorganisms. For instance, Niu et al. (2014) indicated that 20 mT SMF could strengthen cold adaptability of activated sludge, and some Gram-negative bacteria with better cold adaptability were enriched. The mechanism was that SMF would promote production of unsaturated fatty acids in cell membrane to stimulate 2,3,5-triphenytetrazolium chloride dehydrogenase activity, which was important for adapting to low temperature. The result of phospholipid fatty acid analysis showed that C16:1ω7c, C18:1ω7c, and C18:1ω9c were essential unsaturated fatty acids in cell membrane at low temperature (Niu et al., 2013). In addition, Turk et al. (2007) described that cell membrane remodeling was also essential for successful adaptation to altered physicochemical environments.
For instance, some yeast-like fungi could regulate composition of their fatty acyl chains to adapted hypersaline conditions (Turk et al., 2004). In this case, a low sterol-to-phospholipid ratio and increased unsaturated fatty acids would result in high plasma membrane fluidity over a wide range of NaCl concentrations. As already mentioned, regulation of cell membrane composition could be promoted by SMF. It could be proposed that SMF might accelerate or even change regulatory mechanism of fatty acids in cell membrane of microorganisms, thus enhancing their tolerance to hypersaline conditions.
In addition, Liu et al. (2008) indicated that biodiversity of anammox community decreased when exposed to SMF; however, proportion of effective species dramatically increased, thus promoting the anammox process. It was suggested that treatment efficiency and adaptability of microbial community would also be improved by SMF through regulating microbial structure and selectively enhancing growth of potentially effective species. High-throughput sequencing results in this study showed that biodiversity of both bacterial and fungal communities with 45.3 mT SMF was lower than that without SMF, which was consistent with the conclusion by Liu et al. (2008). In addition, dominant bacteria mostly belong to Proteobacteria, Bacteroidetes, and Firmicutes, which were common bacterial phyla in hypersaline wastewater (Lefebvre et al., 2006; Sánchez et al., 2013).
It was obvious that Bacteroides, Salinivibrio, Shewanella, and Vibrio were four absolutely dominant bacterial genera, the total relative abundance of which was both >90% in 0# and 1# samples. Among them, many species belonging to Salinivibrio had been identified as halophilic bacteria (Amoozegar et al., 2008; Zhu et al., 2008). One Salinivibrio costicola strain GL6 was reported that could transform caffeine to theobromine under hypersaline conditions, suggesting that it might be capable of decomposing other recalcitrant aromatic organics that might be formed during biodecolorization of azo dyes (Ashengroph, 2017).
In addition, many bacteria belonging to Bacteroides and Shewanella had been related to biodegradation of azo dyes (Sun et al., 2013; Xu et al., 2016). Bacteroides was the most dominant bacterial genus in the original MMC; however, its relative abundance in 0# and 1# samples was obviously lower. In comparison, the relative abundance of Shewanella and Vibrio sharply increased after treatment with 45.3 mT SMF. Some reports showed that Shewanella and Vibrio could be stimulated by magnetic field based on different mechanisms (Li et al., 2011; Talà et al., 2014), which could support the result that they were enriched when exposed to SMF. Furthermore, ∼50% of bacteria in sample 1# were affiliated to genus Shewanella, a putative halotolerant bacterial group capable of degrading azo compounds (Meng et al., 2014; Xu et al., 2016).
On the other hand, fungal communities in three samples were all dominated by Basidiomycota, Ascomycota, and some unclassified phyla. It was reported that many fungal species belonging to Basidiomycota and Ascomycota were identified as efficient degraders of recalcitrant pollutants (Kües, 2015). Some fungal genera affiliated to unclassified families Trichosporonaceae and Pseudeurotiaceae were only dominant in 1# sample, which suggested that they might be stimulated and enriched with 45.3 mT SMF. It was reported that unclassified Pseudeurotiaceae was recently determined as a dominant phylotype in an SMF-enhanced fungal biotrickling filter for removing trichloroethylene (Quan et al., 2018), which suggested that some species affiliated to this phylotype was sensitive to magnetic field.
Meanwhile, Trichosporonaceae was recently determined to be one of the dominant families in biological wastewater treatment systems and aerobic granular sequencing batch reactors (Gonzalez-Martinez et al., 2018; Muñoz-Palazon et al., 2018), suggesting that some species belonging to this phylotype were related to biodegradation of organic pollutants. In addition, it was notable that Issatchenkia became the most abundant fungal genus in 0# and 1# samples. Furthermore, the relative abundance of Issatchenkia in 1# sample was higher than in 0# sample, suggesting that its growth was stimulated by SMF. It was previously reported that one yeast belonging to genus Issatchenkia could degrade azo dyes (Ramalho et al., 2004), thus the dominance of Issatchenkia supported the conclusion that decolorization efficiency was improved by SMF.
Therefore, it could be concluded from the mentioned results that some putative bacterial and fungal species were selectively stimulated by SMF, thus resulting in higher biodegradation efficiency.
Summary
The purpose of this study was to investigate effects of SMF on biodecolorization of azo dyes by MMC under high-salt conditions. It was shown that decolorization, COD removal, and detoxification were all improved by 45.3 mT SMF, by contrast, SMF of higher intensity displayed inhibitory effects. On the contrary, biomass multiplication of the MMC was not inhibited by SMF. In addition, 45.3 mT SMF also enhanced halotolerance of the MMC. Microbial community structure was analyzed through the MiSeq sequencing method to illustrate the effect of 45.3 mT SMF on dominant microbial species. The results demonstrated that the relative abundance of some putative bacterial and fungal species increased with SMF, resulting in better treatment performance.
Footnotes
Acknowledgments
The study was financially supported by the National Natural Science Foundation of China (Nos. 51108223 and 51508259) and the General Project of Scientific Research of Liaoning Provincial Education Department (No. L201783644).
Author Disclosure Statement
Neither the entire article nor any part of its content has been published or has been accepted elsewhere. Meanwhile, the article is not under consideration by another journal at the same time as EES. All the authors have approved the submission to EES.