
Abstract
Abstract
Experiments were completed to assess the effectiveness of Biochar, ThiolSAMMS®, SediMite™, and Organoclay-199 to decrease methylmercury (MeHg) production by the known mercury methylator, Desulfovibrio desulfuricans ND132 (ND132), and to decrease aqueous (i.e., 0.2 μM filter passing) MeHg concentrations. Mercury (Hg) was introduced as Hg(II) or a preequilibrated Hg-DOM (dissolved organic matter) complex. Individual experiments tested the effect of sulfate concentration and Hg-sorbent equilibration time (up to 1 year). The sorbents provided little, if any, inhibition of total MeHg production, but decreased the percentage of MeHg that passed a 0.2 μm filter. Total MeHg produced was substantially greater than the calculated equilibrium aqueous inorganic Hg (Hgi), based on sorption isotherms, implying mercury assumed to be sorbed at the start of the assay was bioaccessible over the 24-h methylation timeframe. MeHg production increased when Hgi was introduced as an Hg:DOM complex, compared to Hg(II). Some of the increased MeHg production may be attributed to lower Hgi sorption with DOM present, but a substantial amount of sorbed Hg must have been accessible also. DOM also increased the filter-passing MeHg fraction, stressing the importance of Hg-DOM and MeHg-DOM interactions in experimental investigations. Sulfate concentrations did not affect Hg(II) methylation and could not explain the increased methylation seen with the Hg:DOM complex. Extended equilibration of Hg-sorbent mixtures for up to 1 year before the methylation assays did not affect the amount of MeHg produced for Biochar or SediMite treatments, but significantly lowered MeHg production for ThiolSAMMS. Inclusion of DOM in these experiments presented the sorbents with a more environmentally relevant form of Hg and lowered sorbent effectiveness to decrease MeHg production and sorb produced MeHg.
Introduction
M
Following the conventional line of logic that sorbed contaminants may have reduced bioavailability (Mihelcic and Luthy, 1991; Cornelissen et al., 2005), sorbent materials have been widely investigated as a potential remedial solution to minimize Hg flux and/or Hgi available for microbial methylation. High-affinity sorbent materials can sequester Hgi (Gilmour et al., 2013b; Gomez-Eyles et al., 2013; Tan et al., 2016; Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]), Hg complexed with dissolved organic matter (Hg:DOM complexes) (Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]), and MeHg (Gilmour et al., 2013b; Gomez-Eyles et al., 2013) through enhanced partitioning to the solid phase. The bulk of sorbent testing has focused on the ability of these materials to sorb aqueous Hgi; however, these same materials may also be able to sequester aqueous MeHg (Gilmour et al., 2013b; Lewis et al., 2016).
To date, only a limited number of studies have tested whether sorbent materials decrease Hgi methylation and/or MeHg bioaccumulation. Most studies conclude that sorbent treatments do not reduce (Lewis et al., 2016) or even lead to elevated solid-phase MeHg concentrations (Gilmour et al., 2013b; Shu et al., 2016; Zhang et al., 2018). This increase may be an indirect outcome of sorbent application due to sulfate being released from the sorbents, thus stimulating microbial methylators (i.e., sulfate-reducing bacteria [SRB]) (Shu et al., 2016). Although most studies suggest sorbents do not decrease methylation, Bussan et al. (2016), found that the addition of 5% activated carbon or biochar (BC) to wetland sediments decreased the methylation potential by 80% and 88%, respectively, whereas demethylation potentials were unaffected (Bussan et al., 2016).
Nevertheless, sorbents have consistently been shown to decrease aqueous MeHg concentrations. Sorbents may not inhibit MeHg production, but rather partition produced MeHg to the solid phase (Gilmour et al., 2013b; Lewis et al., 2016; Shu et al., 2016; Zhang et al., 2018). For example, when wetland sediments were amended with granular activated carbon and zero-valent iron, MeHg pore water concentrations decreased; however, MeHg and THg sediment concentrations remained unchanged (Lewis et al., 2016).
Similarly, coblending of biochar with river sediments (5% wt.) decreased aqueous THg and MeHg levels over the long term (t > 600 days), but led to elevated solid-phase MeHg concentrations on the sorbents (Liu et al., 2017). Due to concerns over potential MeHg accumulation in rice plants, several recent studies have investigated the effects of biochar treatments on aqueous, soil, and rice plant MeHg levels. One study found that the addition of straw and biochar increased MeHg production in paddy soils, while decreasing MeHg uptake in rice grains (Zhang et al., 2018). Likewise, an application of 1–4% (wt.) biochar increased MeHg soil concentrations, while decreasing aqueous MeHg and subsequent bioaccumulation in rice plants (Shu et al., 2016). Relevant studies to date and their findings are summarized in Supplementary Table S1.
Environmental microbial mercury methylation is highly complex, where environmental conditions affect both mercury bioavailability and the methylating microbial communities. Hg(II) is generally assumed to be the most bioavailable form of Hgi, with bioavailability differences occurring when mercury is complexed, for example, with DOM (Watras et al., 1998; Haitzer et al., 2002; Chiasson-Gould et al., 2014) or sulfide (Benoit et al., 1999; Zhang et al., 2014). DOM is ubiquitous in natural waters and dominates Hg speciation in the absence of sulfide through strong interactions with the thiol groups present on the DOM (Haitzer et al., 2002). However, because of the highly complex nature of Hg speciation and microbial activity surrounding DOM, conflicting outcomes regarding the effect of DOM on methylation have been observed (Moreau et al., 2015; Bravo et al., 2017; Zhao et al., 2017).
The presence of sulfur acts also on both fronts, with sulfide affecting Hg speciation, and subsequent Hg bioavailability (including formation of HgS particles) (Benoit et al., 1999, 2001; Zhang et al., 2014) and sulfate governing the activity of methylating SRB (Gilmour et al., 1992; Jeremiason et al., 2006; Shao et al., 2012). Furthermore, extended equilibration of Hg solutions has also been linked to decreased bioavailability, likely due to kinetically slow changes in Hg aqueous speciation and formation of nanoparticulate and cryptocrystalline HgS solids (Hintelmann et al., 2000; Zhong and Wang, 2009; Zhang et al., 2012, 2014; Hsu-Kim et al., 2013).
The overall objective of this work was to determine if sorbents decreased MeHg production. Hg or Hg-DOM solutions were equilibrated with one of four sorbents (Biochar [BC], ThiolSAMMS® [TS], SediMite™ [SM], and Organoclay-199). These suspensions were then inoculated with the known Hg methylator Desulfovibrio desulfuricans ND132 (ND132) and total and aqueous MeHg (MeHgT and MeHgFP, respectively) were subsequently quantified. Total methylation was compared to previously developed Hgi sorption isotherms (Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]) that establish a theoretical upper limit to methylation under the assumption that sorbed mercury is not bioaccessible. In addition to investigating the effect of DOM, experiments were designed to address the effects of sulfate concentration and Hg-sorbent equilibration time (up to 1 year) on MeHg production and aqueous–solid partitioning.
We hypothesized that DOM and sulfate addition would increase total MeHg production through decreased Hg sorption and increased bacterial activity, respectively. Conversely, we hypothesized that longer Hg-sorbent equilibration would decrease MeHg production through intraparticle diffusion and formation of stronger Hg-sorbent bonds over time, particularly for heterogeneous sorbents like Biochar. These long-term equilibration studies provide initial insight into the potential effects of long-term sorbent deployments.
Materials and Methods
Mercury solutions
Two sources of mercury were tested: (i) Hg(II) added as Hg(NO3)2 and (ii) Hg complexed with natural DOM (Hg:DOM complex). Both mercury solutions were prepared in a degassed and autoclaved phosphate-buffered saline (PBS) (0.14 M NaCl, 3 mM KCl, 3 mM KCl, 10 mM Na2H2PO4, and 2 mM KH2PO4, pH 7.4). Suwannee River natural organic matter (SRNOM) from International Humic Substances Society was used in the experiments as the standard and well-defined source of NOM. SRNOM (2R101N) contains 50.70% C, 3.97% H, 41.48% O, 1.27% N, and 1.78% S (by wt.). SRNOM was dissolved in PBS, equilibrated overnight at 4°C, and subsequently filtered through a sterile 0.2 μm polyethersulfone (PES) Nalgene rapid-flow filter. Hg(NO3)2 stock solution (1,000 ppm; Brooks Rand standard) was added and the pH was readjusted to 7.4 with NaOH. The Hg:DOM solution was then equilibrated for 5 days at 4°C. The molar ratio of carbon in the DOM to mercury (CDOM:Hg) for all Hg:DOM solutions was ∼450,000:1. This CDOM:Hg ratio was selected based on measurements from a local Hg-contaminated creek because it represents an environmental system where in situ sorbent remediation might be considered for cleanup efforts. Similarly, the initial Hg(NO3)2 [i.e., Hg(II)] solution was made in PBS, readjusted to pH of 7.4, and allowed to equilibrate for 5 days at 4°C.
ND132 bacterial culturing
Desulfovibrio desulfuricans ND132 (ND132) was selected as a model methylator due to its high Hg methylation rates (Gilmour et al., 2011). Following a similar procedure as outlined by Liu et al. (2016), an ND132 culture was grown in modified, sulfate-free, MOY medium (Gilmour et al., 2011). The modified Missouri media modified with yeast extract (MOY) (Zane et al., 2010) medium consisted of 40 mM pyruvate, 40 mM fumarate, 30 mM Tris-Cl, 20 mM NH4Cl, 8 mM MgCl2, 2 mM NaH2PO4-K2HPO4, 1.2 mM sodium thioglycolate, 0.6 mM CaCl2, 0.1% (w/v) yeast extract, 0.5× Trace Elements, 0.1× Thauers Vitamins, and FeCl2/EDTA (0.06 mM/0.12 mM) at pH 7.2. ND132 was grown under anaerobic conditions in crimp cap sealed glass containers, degassed, and replaced with an oxygen-free headspace (80% N2/20%CO2). The culture was grown to mid-log phase at 30°C, while being gently shaken at 100 rpm on an orbital shaker. After 4 days and checking the optical density (λ = 600 nm) to ensure sufficient growth, the ND132 culture was brought into an anaerobic chamber (∼98% N2/1–2% H2). The cells were centrifuged at 5,000 g for 5 min and the cell pellet was resuspended in pH 7.4, deoxygenated PBS. The cells were washed three times before the final resuspension in PBS to ensure negligible MOY media remained. Note that the media were specifically altered to be sulfate free to avoid production and carryover of sulfide into methylation experiments. Nevertheless, the cells require a source of sulfur for normal healthy growth and it remains possible that minute amounts of reduced S, primarily from the thioglycolate, were carried over following the triple-rinse procedure. The final cell suspension of ∼109 cells/mL was amended with pyruvate and fumarate for a final concentration of 1 mM, respectively, directly before culture addition into the individual Hg methylation experiments. To control for any differences in cell culturing between experiments, each bioassay had its own set of no sorbent (NS) Hg(II) control experiments (which were completed in parallel) and the results were internally compared against its specific control experiment. Differences in the microbial activity, or other experimental conditions, were minimized through this internal comparison.
Sorbent materials
Four sorbent materials were tested: BC, TS, SM, and Organoclay PM 199 (OC199). BC was purchased from Biochar NOW, where a feedstock of woody materials is converted to biochar by slow pyrolysis kilns. All tested Biochar NOW was from the same batch to ensure uniform sorbent properties. ThiolSAMMS (Steward Environmental Solutions) is a functionalized self-assembled monolayer on mesoporous support material that was developed specially for mercury removal, although the material has a high binding affinity for other metal species such as Ag, Cd, Cu, and Pb. SediMite (Sediment Solutions LLC) pellets contain 50% powdered activated carbon (by wt.), which is weighed down with clay, starch, and sand to aid in delivering the activated carbon to the sediments. OC199 is a proprietary sorbent developed by CETCO with a high capacity for oils, nonaqueous-phase liquids, and dissolved high molecular weight compounds.
Experimental setup
Approximately 0.1 g of sorbent material was weighted into 15 mL sterile, metal-free, polypropylene plastic centrifuge vials and wrapped in aluminum foil to limit light exposure. These vials and the preequilibrated Hg solutions were brought into the anaerobic chamber (∼98% N2/1–2% H2) to equilibrate overnight in the oxygen-free environment; 1.5 mL of Hg solution was then added to each of the vials and allowed to equilibrate for an additional 48 h. The 48-h sorbent-Hg equilibration time was deemed sufficient based on kinetic sorption experiments previously conducted for each of the sorbent materials (Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]). After the ND132 culture was concentrated and rinsed free of MOY media, 1.5 mL of culture was added to each vial, resulting in an initial Hgi concentration of 15–20 ng/mL and an ND132 cell concentration of 5 × 108 cells/mL.
Following a 24-h incubation period with ND132, vials were removed from the anaerobic chamber for MeHg sampling and analysis. For the SM samples, since aqueous–solid phase separation by gravity was not sufficient for successful aqueous-phase sampling, these vials were centrifuged for 15 min at 2,500 rpms before sampling. Approximately 1 mL of aqueous phase was filtered through a 0.2 μm PES syringe filter into a new 4 mL amber glass vial (National Scientific). Both the original and the amber glass vials were immediately acidified to 0.5% hydrochloric acid (trace metal grade) to stop microbial activity and preserve the samples for MeHg quantification. Hg methylation experiments for each treatment were completed in replicates (n = 5).
Two types of control experiments were run: no Hg controls to check for the presence of any available background Hg and ambient MeHg on the sorbents, and no ND132 controls to ensure no abiotic Hg methylation occurred and that no MeHg artifacts were created during the extraction and analysis procedures. To keep the final volume constant, 1.5 mL of deoxygenated PBS was added to replace either the Hg or ND132 solutions in the control experiments. Control experiments confirm that there is negligible MeHg contamination on the sorbents, negligible MeHg created in the extraction, distillation, and MeHg measurement processes, and negligible abiotic methylation occurring over the course of the experiments (Supplementary Tables S11–S13).
In control studies where we tested for loss of Hg to container walls (without sorbent present), 95% ± 3% (n = 6) of the added Hgi was recovered in the aqueous phase (Supplementary Table S14). pH measurements confirmed that BC, SM, and OC199 had minimal effect on the pH of the PBS. Regardless of Hg source, less than a 0.35 pH unit change occurred for BC (final pH = 7.60 ± 0.12), SM (final pH = 7.71 ± 0.03), and OC199 (final pH = 7.38 ± 0.05), whereas TS decreased pH from 7.4 to 6.74 ± 0.01. Since ND132 is known to have a wide range of pH tolerance (Gilmour et al., 2011), the small pH changes created from sorbent addition were assumed not to affect ND132 activity.
Sulfate addition experiments
Sulfate release from SRNOM and sorbents was analyzed by ion chromatography (Dionex ICS-2100 IC system with an IonPac AG9-HC RFIC guard [4 × 50 mm] and analytical column [4 × 250 mm]). The addition of SRNOM resulted in a final sulfate concentration of ∼30 mg/L in the bioassay experiments. Testing indicated that sorbents also release sulfate to varying degrees, and thus also supply additional sulfate to the system. For our experimental setups, BC and TS released minimal sulfate, whereas both SM and OC199 released ∼65 mg/L SO42−.
To test the influence of additional sulfate on Hg methylation, additional bioassay experiments were completed with five treatment groups: (a) 0 mg/L; (b) 25 mg SO42−/L; (c) 50 mg SO42−/L; (d) 150 mg SO42−/L; and (e) Hg:DOM. The sulfate addition concentrations were selected to bracket the end members of possible sulfate release due to either DOM or sorbents. In experiments a–d, Hg was added as Hg(II) and sulfate from a stock solution of Na2SO4. For experiment e, Hg was added as Hg:DOM with no additional sulfate added. Treatment groups were set up in replicates (n = 5). Control experiments without ND132 were conducted alongside the bioactive experiments.
Preequilibration of Hg(II)
To test the effect of Hgi-sorbent contact time on MeHg formation by ND132, Hg(NO3)2 solutions were preequilibrated with sorbents for 2, 144, or 383 days. The 144-day long-term Hg(II) equilibration experiments were run in the manner described in the experimental setup section and thus are directly comparable to the 2-day short-term Hg equilibration experiments. Because of method improvement over time, there are slight differences in the setup methods for the 383-day Hgi-aging experiments. BC was the only sorbent tested in the 383-day Hg(II) equilibration studies. These aging experiments were conducted in 60 mL glass vials containing 25 mL PBS, 15 pg/μL Hg [as Hg(II)], and 0.1 g BC, in the sorbent treatment experiments. The vials were degassed with N2 (nitrogen gas), sealed, wrapped in foil, and placed on a shaker tray for the duration of the equilibration period.
Approximately 1 week before the methylation experiment, the headspace was purged with N2 again, and the vials were moved into the anaerobic chamber. To ensure the same final ND132 cell concentration of 5 × 108 cells/mL, 30 mL of the rinsed and concentrated culture was added to each of these experiments. The solid–liquid ratio of the 383-day Hg equilibration experiments was 20 times lower than the rest of the experiments. Although the longer-term equilibration methods are slightly different than the other Hg methylation studies, these results still provide insight into the effect of Hg aging since NS control experiments were run alongside the BC treatment experiments, providing an internal comparison. Because of these experimental differences, only the ratio of the BC treatment to the NS control for the long-term (383-day) experiments should be used to draw any conclusion on treatment effectiveness for aged Hg.
MeHg quantification
To quantify the total MeHg produced (MeHgT), a nonaqueous-phase extraction was first conducted (Bloom et al., 1997) to extract MeHg from the solid, sorbent materials, and bacterial cells. Before extraction, an internal standard of either Me200Hg or Me202Hg, depending on the expected MeHg concentration, was added. Next, two extraction solutions, one containing potassium bromide (KBr) and sulfuric acid (H2SO4) and another containing copper sulfate (CuSO4), were added to each vial to produce a final concentration of 1.26 M KBr, 0.75 M H2SO4, and 0.17 M CuSO4. The vials were shaken vigorously overnight. Next, 10 mL of methylene chloride (CH2Cl2) was added and vials were shaken again for 1 h. The nonaqueous phase, CH2Cl2, was then transferred into a 50 mL polypropylene centrifuge tube containing 20 mL of distilled water. The MeHg was then back-extracted into the aqueous phase as the CH2Cl2 was driven off, while the vials were placed in a hot water bath and bubbled with N2. To ensure sufficient removal of the Hgi, both the MeHgT and MeHgFP samples were distilled and analyzed following EPA Methods 1630 (EPA, 2001). MeHg concentration was measured using a Brooks Rand MERX Hg Speciation GC and Pryolysis coupled with an ELAN DRC-e ICP-MS. Quality assurance/quality control (QA/QC) for MeHg analysis is presented in the Control Experiments and MeHg Analysis QA/QC section of Supplementary Data.
Statistical analyses
Statistical analyses were conducted in R (R Core Team, 2016) using the “agricolae” package (Mendiburu, 2017). Significant differences among treatments within an experiment were examined using a Tukey Honestly Significant Differences Test holding the family-wide Type I error rate at 5%. The full statistical output is provided in the Supplementary Data.
Aqueous Hgi estimates
Using developed sorption isotherms from Eller et al. (2018), Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, (in review) for BC, TS, SM, and OC199, for Hg(II) and Hg:DOM, the amount of aqueous and solid-phase Hgi in the system was estimated. These isotherms were developed using comparable methods (i.e., background solution, CDOM:Hg ratio, aqueous–sorbent ratio, and equilibration time). Uncertainty estimates were calculated using standard rules for the propagation of uncorrelated uncertainty (Taylor, 1997) using the standard error associated with all inputs, including sorption isotherm parameters.
Results and Discussion
The ability of sorbent materials to decrease total MeHg production (MeHgT), for two different sources of Hg [i.e., Hg(II) and Hg complexed with DOM], was tested through a series of bioassay experiments using the known methylator D. desulfuricans ND132. A set of NS control experiments was conducted in parallel to provide a means of direct internal comparison. Caution should be exercised if attempting to compare across different bioassay experiments, and we recommend that only the ratio of the treatment to the NS control should be used across experiment comparisons. It should be noted that there are no data for the Hg(II)-SM experiments because we were unable to quantify MeHgT in that treatment. The fraction of Hgi that was methylated over a 24-h contact period with ND132 (FMeth) was used to assess the effect of the various sorbent treatments on microbial methylation. FMeth is used as a metric to account and normalize for the slight differences between the initial concentration of the Hg(II) and Hg:DOM solutions used in the experiments. Aqueous MeHg (MeHgFP) is quantified and expressed as the fraction of MeHgT that passed a 0.2 μm filter (FFP).
Effect of sorbents on total MeHg production
Sorbents were ineffective at decreasing total MeHg production and this inability to decrease MeHg production supports similar findings for a variety of sorbent materials (Gilmour et al., 2013b; Lewis et al., 2016; Zhang et al., 2018). None of the sorbent treatments supplied a statistically significant reduction in methylation for Hg(II) and only ThiolSAMMS provided significant treatment (60% reduction) when DOM was present. When Hg was provided as Hg(II), there were no significant differences in Hg methylation among the NS and sorbent treatments (Fig. 1a, Supplementary Fig. S1, Supplementary Tables S3 and S4). The baseline fraction methylation [i.e., Hg(II)-NS control] was 0.13 ± 0.0030 (mean ± standard error; n = 5), which is consistent with other ND132 methylation studies conducted under similar experimental conditions (Lin et al., 2015; Zhao et al., 2017). Sorbent treatments did not reduce mercury methylation—Fmeth with OC199 treatment was 0.16 ± 0.019 (n = 5); TS was 0.13 ± 0.024 (n = 5); and BC was 0.092 ± 0.020 (n = 2). Although the BC treatment did show less methylation, the decrease was not statistically significant (p = 0.64). Conversely, on average, the OC199 treatment produced slightly more MeHgT than the NS control, but again this effect was not statistically significant (p = 0.64).

Total MeHg production by ND132 over a 24-h contact period shown as both fraction methylated, FMeth (i.e., MeHgT/Hgi) and total mass of MeHg produced, MeHgT, when Hg is added as
The sorbents were more effective at decreasing MeHgT for the Hg:DOM complex, although the effects were still rather limited (Fig. 1b, Supplementary Fig. S1, Supplementary Tables S3 and S4). Methylation of Hg:DOM in the NS and sorbent treatments was as follows: NS = 0.47 ± 0.049 (n = 5); BC = 0.38 ± 0.039 (n = 5); SM = 0.33 ± 0.048 (n = 3); OC199 = 0.31 ± 0.022 (n = 3); and TS = 0.19 ± 0.017 (n = 5). TS was the only sorbent treatment to provide a statistically significant reduction and decreased MeHg production by 60% ± 27% (p < 0.001). OC199 decreased production by 34% ± 25% (p = 0.092) compared to the Hg:DOM-NS control. On average, the BC and SM treatments also lowered total MeHg, although these reductions were not statistically significant (p = 0.39, p = 0.15, respectively).
Effect of sorbents on filter-passing MeHg
Although the tested sorbent materials in most cases provided little, if any, inhibition of MeHgT production, perhaps of greater relevance is the amount of aqueous, or 0.2 μM filter-passing MeHg (MeHgFP). MeHg that does not pass the 0.2 μm filter is assumed to be associated with either the sorbent or ND132 (either inside the cells or sorbed to their surface). Without a sorbent, a large fraction of the total MeHg produced remained associated with the cells [i.e., MeHgFP = 0.24 for Hg(II) source and MeHgFP = 0.52 for Hg:DOM]. Even so, as hypothesized, all sorbent treatments significantly decreased FFP (Fig. 2, p < 0.001, all treatments) through partitioning to the solid phase.
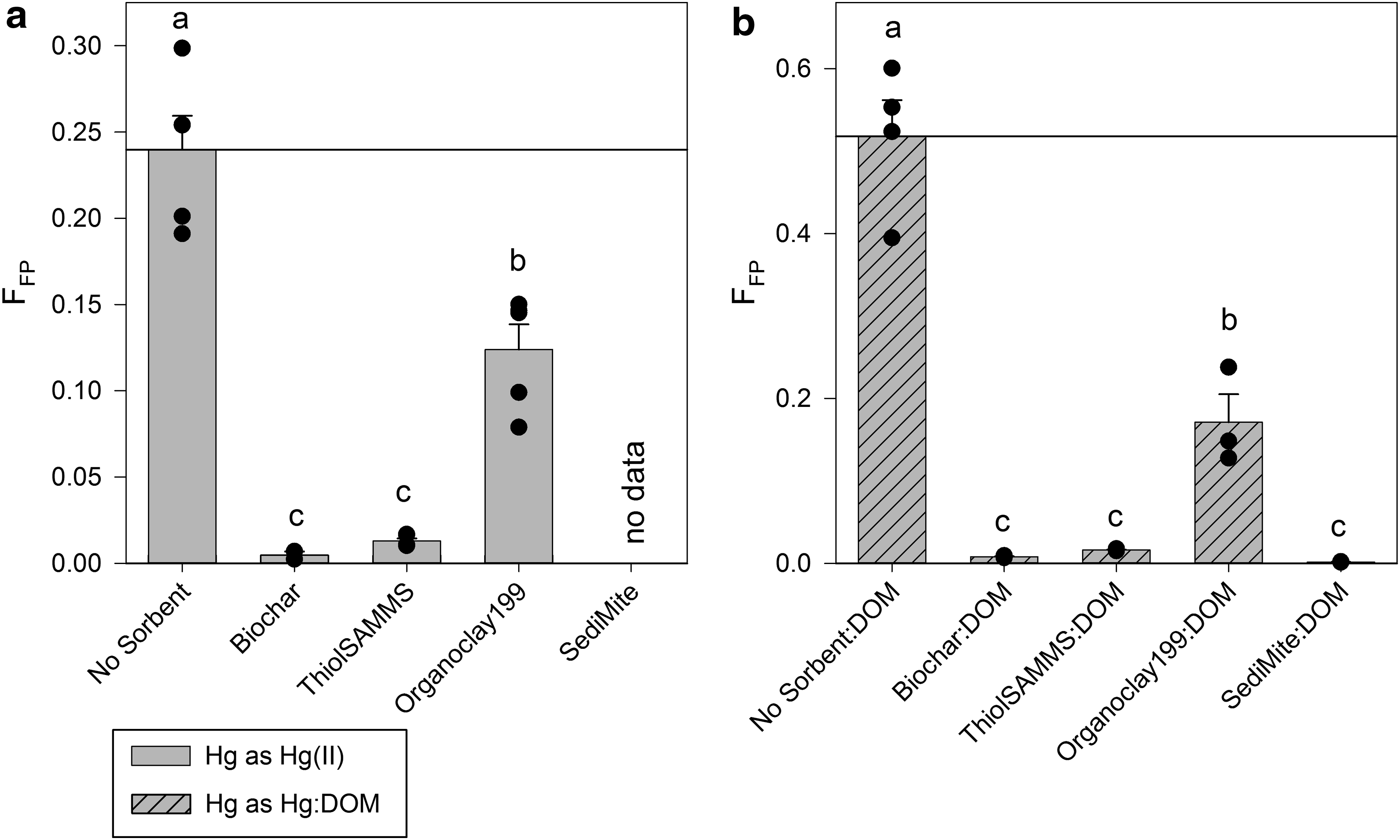
Filter-passing (0.2 μm) MeHg (FFP) as the fraction of the total MeHg produced that was filter passing when Hg is added as
Regardless of Hg source, Biochar decreased FFP by 98%; ThiolSAMMS reduced FFP between 95% and 97%, for Hg(II) and Hg:DOM, respectively; and SediMite decreased aqueous MeHg levels by 99%. OC199 was the least successful sorbent at removing aqueous MeHg, but still provided a 48–67% reduction for Hg(II) and Hg:DOM, respectively. These results do not directly answer the larger question of whether sorbed MeHg is available for bioaccumulation. Nevertheless, since MeHg bioaccumulation in aquatic organisms has been linked to aqueous MeHg levels rather than the total MeHg present (Watras et al., 1998; Munthe et al., 2007; Chasar et al., 2009), our results show promise for sorbents as a remedial tool by lowering MeHgFP concentration. Therefore, MeHgFP is an important parameter for assessing the remedial potential of sorbent treatments for aquatic ecosystems; however, further testing is needed to determine if nonfilter-passing MeHg is sufficiently sequestered and thus unavailable for bioaccumulation.
Aqueous–sorbent partitioning
Based on accompanying Hg sorption isotherms (Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]), the sorbents retained between 95.5% and 99.9% of the Hg(II) and between 89.1% and 99.8% of the added Hg:DOM (Supplementary Table S2). The amount of Hgi left in the aqueous phase after equilibration with the sorbents was compared to the amount of MeHg produced in each of the experiments. In each case, substantially more MeHg was produced than the amount of Hgi calculated to be in the aqueous phase in the presence of the sorbent. When Hg was added as Hg(II), between 358% (OC199) and 138,000% (TS) of the estimated equilibrium aqueous-phase Hgi mass was methylated. Similarly, for Hg:DOM, between 233% (BC) and 11,800% (TS) of aqueous Hgi was methylated. The experimental results demonstrate that methylation occurred above the levels predicted by Hgi sorption isotherms, based on the assumption that only the equilibrium aqueous Hgi mass is available for microbial methylation.
Our calculations imply that (i) ND132 accessed the sorbed Hgi, (ii) ND132 drastically altered the Hgi equilibrium partitioning between solid and aqueous phases, or (iii) desorption and reequilibration of the sorbed Hgi was sufficiently fast compared to the ND132 methylation rate. Analogous to our findings are studies showing that systems containing metacinnabar [i.e., HgS (s) or β-HgS (s) nanoparticles] are methylated at levels much higher than aqueous equilibrium values would predict (Graham et al., 2012; Zhang et al., 2012).
Effect of DOM
The baseline Fmeth for Hg(II) [i.e., Hg(II)-NS control] was 0.13 ± 0.0030 (n = 5), whereas the Hg:DOM-NS control's baseline methylation was 0.47 ± 0.049 (n = 5)—a marked 3.6 ± 0.84-fold increase over the Hg(II) experiments (Fig. 1). Note that all experiments presented in Fig. 1 were conducted in parallel, and thus, all these FMeth data are directly comparable. Since ND132 is a sulfate-reducing bacterium, we hypothesized that the increased MeHgT for the Hg:DOM complex may have been confounded by the unavoidable addition of sulfate by the SRNOM. Therefore, additional experiments were conducted to test the effect of added sulfate in the absence of sorbents or DOM (Effect of Sulfate section).
SRNOM also increased FFP (p < 0.001) from 0.12 ± 0.022 for the Hg(II)-NS up to 0.52 ± 0.088 for the Hg:DOM-NS experiments (Fig. 2), for an average increase of 322% ± 46% [i.e., Hg:DOM-NS compared to Hg(II)-NS]. This finding highlights the importance of DOM for MeHg speciation and supports the need to conduct experiments under conditions that mimic environmental systems. For example, without consideration of MeHg-DOM interactions, aqueous MeHg levels would be grossly underestimated, potentially leading to miscalculating the risk of MeHg exposure and bioaccumulation.
Effect of sulfate
To test if the increase in Hg methylation with DOM was due to an unavoidable addition of sulfate from the SRNOM, which subsequently increased microbial activity, an additional methylation experiment was completed. Although the primary goal in this study was to discern if the sulfate released from the DOM could explain the increase in methylation, these experiments also addressed a similar hypothesis that differential sulfate release from sorbents accounted for some variability in methylation among the sorbent treatments. At the DOM level used in experiments, the final SO42− concentration in the vials was 29.5 ± 0.1 mg/L. Sulfate release from sorbents ranged from less than 1 to 65 mg/L in our experiments (SM = OC199 > TS > BC), and our five sulfate treatments were established to determine if sulfate concentrations over this range increased Hg(II) methylation. Sulfate concentration did not influence Hg methylation (Fig. 3, Supplementary Fig. S2, Supplementary Tables S5 and S6) and therefore, sulfate release could not account for the increased methylation seen in the Hg:DOM experiments. Based on these findings, an additional unknown mechanism beyond sulfate concentration must be responsible for the differences in methylation between the two sources of Hg. Furthermore, these results suggest that sulfate release from sorbent materials did not influence methylation results.
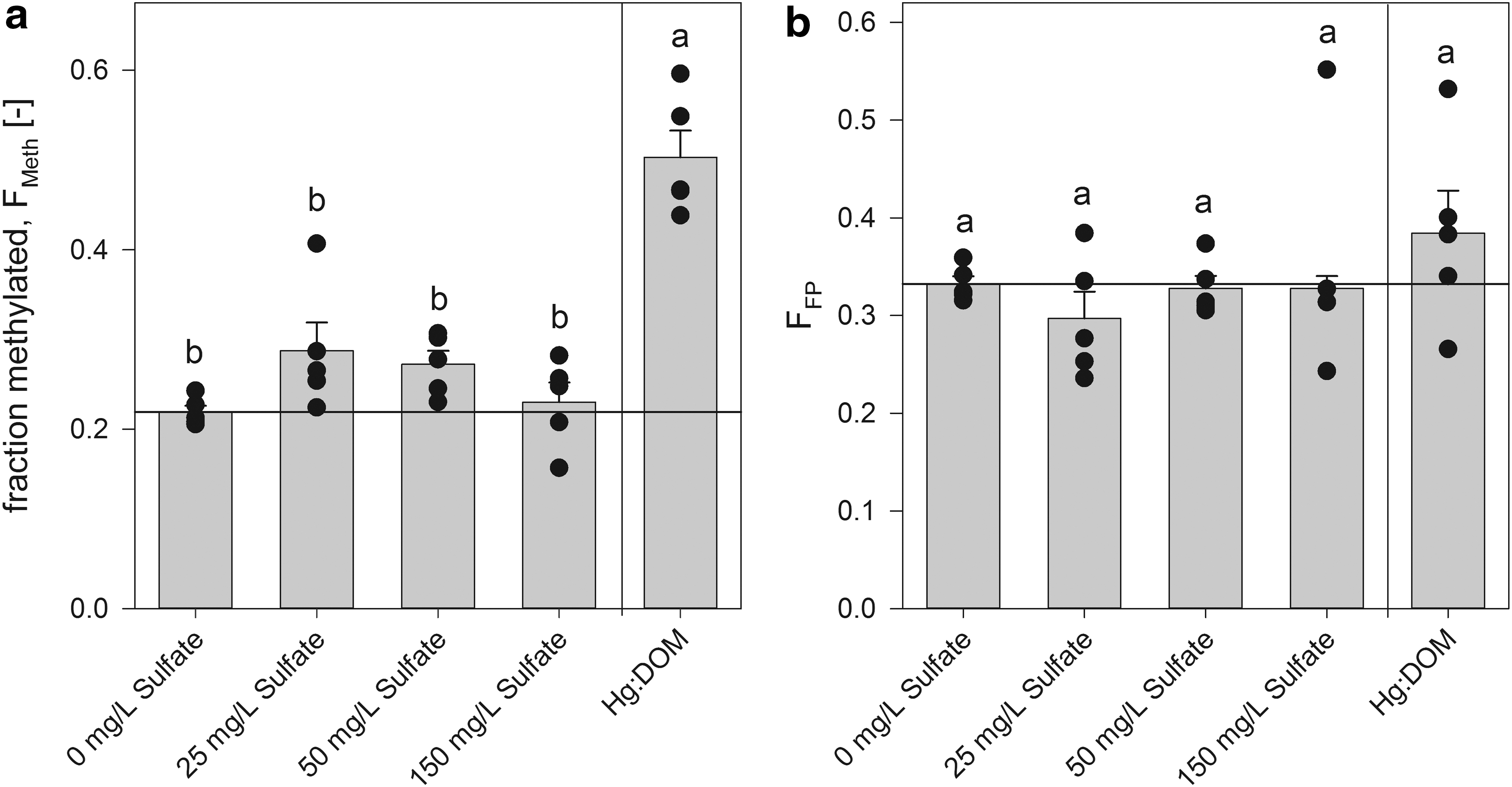
Effect of sulfate additions on
In addition, these results suggest that potential sulfide production by ND132 from added or released sulfate and consequent effects on Hgi speciation had no discernible effect over the time course of these experiments. MeHg production was the same in the presence of BC, which released no detectable sulfate, and SM, which released the most sulfate. It is possible that minute amounts of sulfide were carried over during the triple cell washing process, but inclusion of some sulfur in the growth media is unavoidable as the cells require a source of sulfur. Finally, if sulfide had a significant effect on Hgi speciation and MeHg production, we would have expected a much smaller difference between the Hg(II) and Hg:DOM NS control experiments (Fig. 1).
Preequilibration of Hg(II)
Total MeHg production and aqueous MeHg concentrations were evaluated as Hgi [as Hg(II)] was preequilibrated in the system for extended periods of time. Equilibration durations of 2, 144, and 383 days are compared both with and without sorbents for total MeHg production and filter-passing MeHg. The effect of preequilibrating Hg in the NS controls was statistically significant, with the total MeHg production decreasing between the 144 and 2 day treatment by 57% ± 8% (p = 0.002), and by 98% ± 2% (p < 0.001) after the 383-day equilibration (Fig. 4a). This effect may have been due to Hg partitioning to the container walls (PES vials were used in the 2- and 144-day experiments, in attempt to limit Hg and MeHg sorption), regardless these trends serve as a control and comparison for long-term Hg equilibration with sorbents. Decreasing methylation may be the product of changing Hg speciation over time to less bioavailable forms (Zhang et al., 2012).
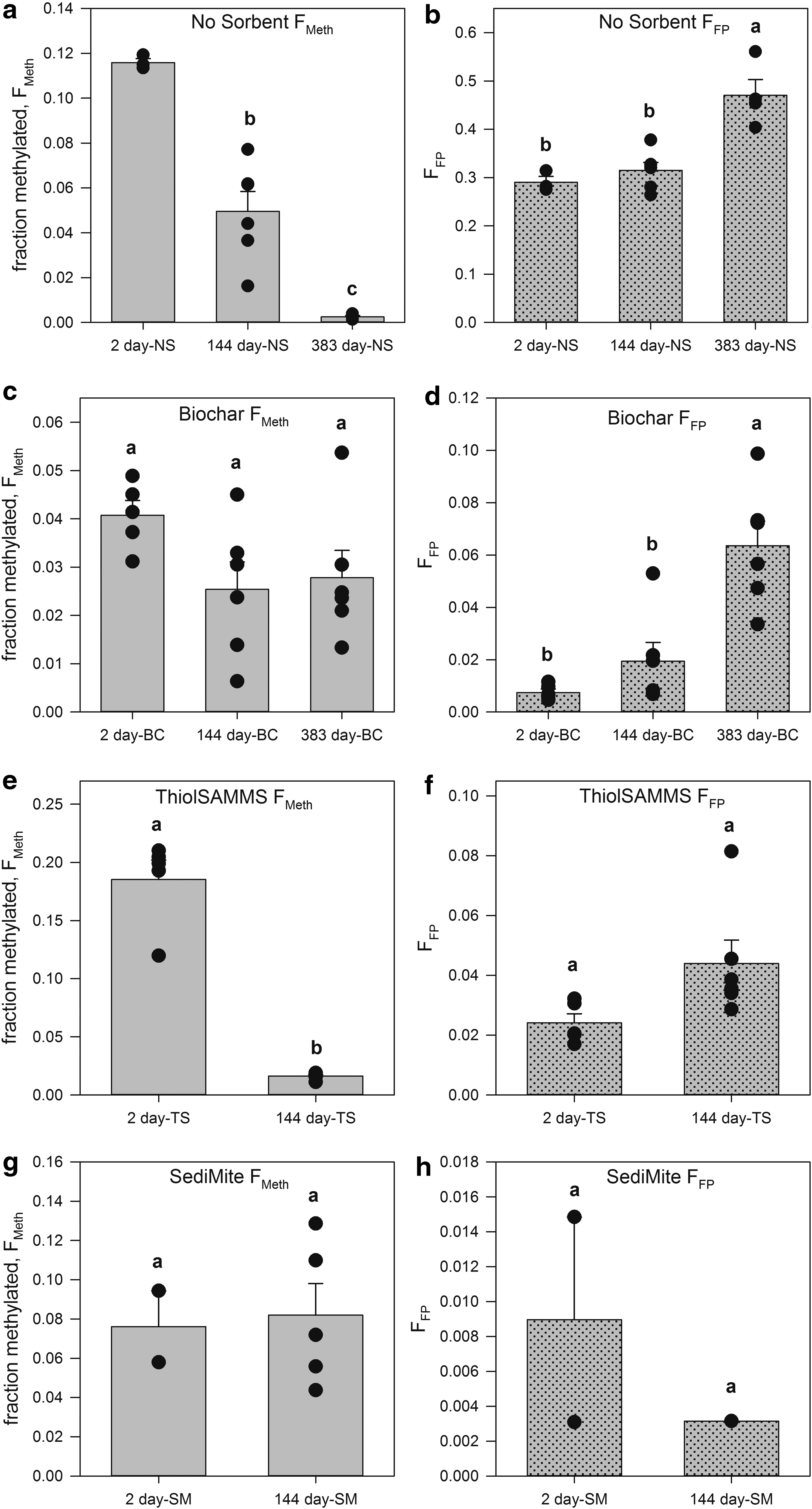
Effect of Hg-sorbent equilibration time on sorbent treatment efficiency expressed as fraction methylated, FMeth (left), and filter-passing MeHg as a fraction of the total MeHg produced, FFP (right), without a sorbent
Increased preequilibration time between Hg and sorbents was hypothesized to decrease methylation rates based on the body of literature suggesting that true sorption equilibrium occurs over very long-time scales and/or sorbates may become more recalcitrant over time (by e.g., intraparticle diffusion) (Ball and Roberts, 1991; Valderrama et al., 2008; Wu et al., 2009; Lamichhane et al., 2016). No clear trend emerged for FMeth as a function of varying Hg-sorbent preequilibration time. TS decreased FMeth from 0.19 ± 0.017 (n = 5) after the 2-day equilibration period down to 0.016 ± 0.0012 (n = 6) after 144-day Hg equilibration, providing a nearly 91% decrease in methylation (p < 0.001).
However, when Hg(II) was equilibrated with BC for similar periods, no significant decrease in FMeth was measured. The fraction of methylated Hgi in the presence of BC went from 0.041 ± 0.0031 (n = 5) to 0.025 ± 0.0057 (n = 6) to 0.028 ± 0.0057 (n = 6) after a 2-, 144-, and 383-day Hgi-sorbent equilibration, respectively (p = 0.13, 0.23, respectively, when comparing aged experiment to the 2-day experiment; Fig. 4c). Furthermore, the extended equilibration period did not affect Hgi methylation percentages for the SediMite treatment with 0.076 ± 0.018 (n = 2) methylation after 2-day equilibration and 0.082 ± 0.016 (n = 5) after 144-day equilibration (Fig. 4g). Full statistical output on the effect of Hg-sorbent preequilibration time by sorbent treatment and duration is presented in the Effect of Hg-Sorbent Equilibration Time on Methylation (Statistical Outputs) section of Supplementary Data (Supplementary Figs. S3 and S4, Supplementary Tables S7–S10).
In the NS controls, FFP was significantly greater for the 383-day equilibration time than either the 2- or 144-day equilibration (p = 0.001 for both). The cause of this effect is unclear and the role of potential experimental setup differences cannot be entirely ruled out. Still, all sorbent materials reduced FFP compared to the NS control experiments for the same preequilibration period (p < 0.001 for all treatments). After a 2-day Hgi-sorbent equilibration, FFP was reduced by 97% ± 5.8%, 92% ± 5.7%, and 97% ± 6.1%, for the BC, TS, and SM treatments, respectively (Fig. 4b, d, f, and h). At the 144-day equilibration point, very similar reductions in FFP were measured, 94% ± 7.5%, 86% ± 7.3%, and 99% ± 7.3%, for the BC, TS, and SM treatments, respectively, when compared against the 144-day NS treatment control. In the longest Hg equilibration of 383 days, BC reduced the fraction of filter-passing MeHg over the NS control by 86% ± 9.4% (p < 0.001). These experimental results demonstrate the ability of the tested sorbent materials to effectively remove the produced MeHg from the aqueous phase through time. Nevertheless, within the BC treatment, FFP was higher after the 383-day equilibration period relative to either the 2- or 144-day equilibration treatments (p < 0.001 and p = 0.001, respectively).
Conclusions
This work provides further insight into the usefulness of sorbent materials for in situ Hg remediation. Most sorbent evaluations are either field microcosms, where the importance of specific environmental factors cannot be easily discerned, or simplified laboratory tests often conducted in the absence of relevant environmental ligands and over very short time frames. In this study, we start to fill the gap by conducting laboratory experiments under more environmentally relevant conditions (i.e., DOM, sulfate, and Hg-sorbent equilibration time).
Overall, sorbents did not reduce MeHgT production (or FMeth) by ND132. The only significant reduction from a sorbent treatment was with ThiolSAMMS, but only for the Hg:DOM complex. This finding is consistent with the growing body of evidence that finds sorbents provide limited, if any, treatment for total MeHg production (Gilmour et al., 2013b; Lewis et al., 2016; Zhang et al., 2018). Based on the Hgi-sorbent isotherms, the large amount of MeHg produced relative to the expected aqueous Hgi suggests that ND132 methylated a portion of Hgi typically considered not bioavailable. This result runs counter to the traditional assumption that sorbed contaminants are unavailable or at least less accessible for further transformations. Within the Hg microbial methylation literature, evidence indicates that methylators can access solid particles such as metacinnabar for methylation (Graham et al., 2012; Zhang et al., 2012). This and our findings support the notion that Hg species typically considered to have limited bioavailability may be more available for methylation than previously expected. Such findings suggest that remedial solutions may be overestimating their ability to successfully sequester contaminants and cleanup plans may need to be updated to account for the higher than assumed bioavailability of sorbed pollutants.
DOM increased MeHg production by ND132. Although the mechanisms for the boost in methylation are unknown, our finding is in agreement with most studies (Graham et al., 2012, 2017; Moreau et al., 2015; Bravo et al., 2017), although recent work indicates the effect of DOM on methylation may be bacterial strain dependent (Zhao et al., 2017). Previous work indicates that the presence of DOM decreases Hgi sorption onto all the sorbent materials (Eller et al., 2018, Evaluation of sorbent materials for removal of mercury from contaminated freshwater ecosystems, [in review]); however, methylation was still much greater than predicted by the equilibrium aqueous-phase concentrations. This again suggests that bacteria were able to access a fraction of the Hg associated with the sorbents. Hg:DOM complexes are more representative of mercury speciation under real environmental conditions and due to the dramatic influence of DOM on Hg and MeHg sorption, methylation, and aqueous concentrations, this study highlights the need to conduct experiments with DOM as opposed to simple Hg(II), as is commonly done in the laboratory. Without consideration of DOM, aqueous MeHg levels would be greatly underestimated, potentially leading to miscalculating and underrepresenting MeHg bioaccumulation and the associated risks.
All tested sorbents decreased FFP, regardless of the Hg source [i.e., Hg(II) or Hg:DOM]. These results imply sorbents can successfully partition MeHg in addition to Hgi. Because of the substantial MeHg solid–aqueous partitioning observed, further testing is needed to determine if sorbents can reduce bioaccumulation in target aquatic organisms. If organisms only uptake aqueous MeHg, then sorbents may be able to help break the chain of bioaccumulation; however, since there was no reduction in total MeHg production, if organisms access the MeHg bound to sorbents, or if the MeHg desorbs from the sorbents over time, then sorbent treatment may not effectively reduce bioaccumulation.
Footnotes
Acknowledgments
This work was funded by the U.S. Department of Energy's Oak Ridge Office of Environmental Management (ORO-EM) and URS | CH2M Oak Ridge LLC (UCOR) and is a product of ORNL's Mercury Remediation Technology Development Program.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.