
Abstract
Abstract
Chromium pollution has grave consequences and can cause serious damage to biological systems. Recently, polyamines have received significant attention due to their biotechnological interests. This study is aiming to test the effect of putrescine on hexavalent chromium [Cr(VI)] removal by Wickerhamomyces anomalus and cell surface physicochemical properties of this strain. The experiment was conducted in modified yeast extract, peptone, and glucose medium contaminated with 50 mg/L Cr(VI) and treated with putrescine at 0 (control), 1, and 10 mM. Results showed that putrescine enhanced Cr(VI) removal potential, accompanied by an increased hydrophobicity and electron acceptor character and a decreased electron donor character of surface cells. This putrescine action was dependent on its concentration with a higher effect at 10 mM. At this concentration, scanning electron microscope/energy dispersive X-ray analysis showed a modification of the chemical composition and a high concentration of nitrogen on yeast cell wall. Sorption data were modeled using the pseudofirst-order and pseudosecond-order equations. Results indicated that a pseudosecond-order kinetic model could better describe the metal sorption kinetics by W. anomalus cells at different putrescine concentrations. This work highlights the possibility of enhancement of wastewater treatment, especially heavy metal uptake, by modifying cell surface characteristics.
Introduction
Due to their very toxic nature, even at trace concentrations, nonbiodegradability, and bioaccumulation, heavy metal pollution is a persistent threat to our life and environment (Batayneh, 2011; Dhankhar and Hooda, 2011).
Microbial biosorption of toxic metals is a promising approach that offers great potential for industrial wastewater treatment. Biosorption can be defined as the ability of biological material to accumulate heavy metals from wastewater. This may occur through physicochemical interactions such as adsorption to cell walls or via metabolically mediated pathway uptake such as transport and extracellular precipitation by excreted metabolites (Fourest and Roux, 1992). It represents the advantages of being a safe, effective, and a cheap process for treating chromium pollution (Pan et al., 2014). It is a potential alternative to the commonly used treatment technologies such as filtration, electrochemical treatment, evaporation, and ion exchange, presenting the disadvantages of producing large amounts of sludge, low efficiency, high energy, and chemical requirements (Radhika et al., 2006).
Biosorption efficiency of heavy metals by microbial biomass is mainly connected to cell wall structure, which determines the nature of the interactions between the organisms and metals. The effect of different products on cell surface such as biosurfactants has been previously investigated (Liu et al., 2011; Ławniczak et al., 2013). These products lead to a modification in physicochemical properties of microorganisms' surface and therefore influence their bioremediation efficiency. Polyamines represent another group of compounds that have a potential role in cell surface characteristics since their effects in biofilm formation are studied (Lee et al., 2009; Goytia et al., 2013). In the present investigation, putrescine or tetramethylenediamine [H2N(CH2)4NH2] was studied. It is a ubiquitous low-molecular-weight polyamine that plays multifunctional roles in cell growth, differentiation, and survival (Paz et al., 2011). This compound is formed in animal tissues by the breakdown of amino acids or by intestinal bacteria by decarboxylation of lysine and ornithine in the intestine (Chmielewski et al., 2013). Nevertheless, little is known about its interaction with heavy metals and its involvement in the bioremediation process (Tahri Joutey et al., 2014). Relevant works have shown that the interaction between microorganisms and the abiotic surfaces is of paramount importance in environmental systems (Grasso et al., 2002). In microbially mediated in situ pollutant degradation, these interactions are the key event that determines the microorganisms' migration in geological formation and consequently the efficacy of pollutant removal (Grasso et al., 1996; Grasso and Smets, 1998).
On this basis, we hypothesize that the effect of putrescine on the physicochemical properties of cellular wall could increase the adsorption uptake of heavy metals.
In the present investigation, Wickerhamomyces anomalus as a good chromium biosorbent (Bahafid et al., 2013) was chosen to study the effect of putrescine on its cell surface properties, and on its bioremediation potential of chromium in aqueous solutions.
Materials and Methods
Microorganism
The yeast W. anomalus was previously isolated from contaminated soil with chemical industrial wastes, including tanning processing in Fez, Morocco (Bahafid et al., 2013). This strain showed a high ability to remove chromium and is considered an excellent adsorbent.
Yeast strain was seeded on yeast extract, peptone, and glucose (YPG) medium agar plates (1% peptone, 1% yeast extract, 2% glucose) and incubated for 48 h at 30°C.
Biosorbent preparation
Fifty microliters of YPG medium was added into a 250 mL Erlenmeyer flask, sterilized at 120°C for 15 min, and inoculated by a loopfull of yeast cells. The culture was incubated at 30°C, 150 rpm for 48 h.
After incubation, the cells were then harvested by centrifugation at 6000 rpm for 15 min and washed several times with distilled water. The cell concentration was adjusted to an optical density of 0.7–0.8 at 660 nm (A0) (∼1.0 × 107 colony forming unit/mL cell density) (Bahafid et al., 2013) and used in further experiments.
Effect of putrescine on Cr(VI) removal
A modified YPG medium (0.2% yeast extract +0.2% peptone +2% glucose) was prepared in sterile distilled water and distributed into 250 mL Erlenmeyer flasks before autoclaving (121°C for 20 min) (Bahafid et al., 2011). Then, 1.0 mL of yeast suspension with 1.0 × 107 cells previously prepared was inoculated into each Erlenmeyer flask. The culture was incubated at 30°C, 150 rpm for 48 h.
Potassium dichromate (K2Cr2O7) was used to obtain the hexavalent chromium concentration of 50 mg/L. Putrescine was also added into the culture medium to achieve the final concentrations of 0 (control), 1, and 10 mM separately. Both chromium and putrescine were filtered through a 0.22 μm cellulose acetate membrane filter before addition into the culture medium. To reflect the abiotic reduction of Cr(VI), noninoculated mediums were prepared in the same way. The experiments were then incubated at 30°C in a shaker incubator (150 rpm). Chromium reducing activity was estimated as the decrease in chromium concentration in supernatant using Cr(VI)-specific colorimetric reagent S-diphenylcarbazide (DPC) 25% (w/v) prepared in acetone. After 48 h of incubation, aliquots of cultures (1 mL) were centrifuged in an Eppendorf tube for 10 min at 7,000 rpm. Then, a volume of 200 μL supernatant was added to the reaction mixture set up in a 10 mL volumetric flask as follows: 330 μL of H2SO4 (6 M) followed by addition of 400 μL of DPC, and a final volume was made to 10 mL using distilled water. Spectrophotometric measurements were made immediately at 540 nm (Chrom Tech V-1200) (Pattanapipitpaisal et al., 2001).
All the adsorption experiments were performed in triplicate, and means were used in the data analysis.
Effect of putrescine on cell surface characteristics of W. anomalus
Different techniques can be used in the physicochemical characterization of microbial cell surface properties, such as contact angle measurements by the sessile drop (Busscher and Weerkamp, 1984). In this work, this technique served as a prompt and a simple way to obtain precise and valuable surface energy values. It yields precise estimations of cell surface hydrophobicity (CSH) and electron donor/acceptor character (Busscher and Weerkamp, 1984; Van Der Mei et al., 1998; Hamadi et al., 2008).
Microbial lawn preparation
In parallel to biosorption experiments, contact angle measurements were conducted. These measurements were assessed on lawns of organisms deposited on membrane filters (Van Der Mei et al., 1998). To prepare microbial lawns, 1.0 mL of yeast suspension with 1.0 × 107 cells previously prepared was added into the modified yeast medium (YPG) and incubated at 30°C for 24 h under a shaker system where the log phase is reached. Yeast cells were obtained by centrifugation at 8,000 g. The cell pellet was washed twice using sterile KNO3 (0.1 M) and resuspended in the same solution. The cell concentration was adjusted to an optical density of 0.7–0.8 at 660 nm (A0) in 0.1 M KNO3. An examination using light microscope allows the observation of cells, presenting at this log phase characteristic of yeast strains. Microbial cell suspensions were then deposited on a cellulose acetate membrane filter (0.45 μm) using negative pressure. Cells lawns were placed to air dry for 30–60 min to be stabilized (Busscher and Weerkamp, 1984; Hamadi et al., 2008). Microbial lawns for contact angle measurement were realized in triplicate with separately cultured yeast strains.
Contact angle measurements and calculation of physicochemical characteristics
The measurement of yeast CSH was carried out using a goniometer (GBX Instruments, France), by the sessile drop method (Elabed et al., 2011). By using three different liquids with known surface parameter values γlLW, γl+, and γl−, two polar liquids (water and formamide) and one nonpolar liquid (diiodomethane) (Table 1), the unknown surface tension components of a microbial surface [The Lifshitz–van der Waals component (γLW), the electron donor or Lewis base (γ−), and the electron acceptor or Lewis acid (γ+)] were calculated using Young's equation, Equation (1); the surface free energy is proportional to cosine of the contact angle (Absolom et al., 1983). In principle, Young's equation assumes that the entire system is at thermodynamic equilibrium and that the solid surface is chemically homogeneous, flat, and not influenced by chemical interaction or adsorption of the liquid to the surface (De Meijer et al., 2000). The contact angle measurement was carried out in triplicate and an average of the contact angles and the standard deviation were calculated and reported [(S) and (L) denote solid surface and liquid phases, respectively]:
Energy Characteristics [Lifshitz–van der Waals (γLW), Electron-Donor (γ−), and Electron-Acceptor (γ+) Parameters (mJ/m2)] of Pure Liquids Used to Measure Contact Angles
Where γ is the surface tension (in mJ/m2) of the solid (s), and the liquid (l) interface γLW is Lifshitz–van der Waals component of the solid (s), and the solid (l) and γ− the electron donor character of the solid (s), and the liquid (l) interface and γ+ the electron acceptor character of the solid (s), and the liquid (l) interface, respectively.
The Lewis acid–base component was obtained by the following:
CSH was also estimated using the Van Oss approach (Van Oss et al., 1988; Elabed et al., 2011), where the degree of hydrophobicity of a given surface (i) is expressed as the free energy of interaction between two entities of that material immersed in water (w). The materiel is hydrophobic when ΔGiwi is negative, and conversely, when ΔGiwi is positive, the material is considered being hydrophilic. ΔGiwi is calculated according to the following Equation (3):
Where γiLW is Lifshitz–van der Waals component, γwLW is Lifshitz–van der Waals component of water, γi+ electron acceptor of a given material (i), γi− electron donor of a given material (i), γw+ and γw− are, respectively, electron acceptor and electron donor characters of water.
Sorption kinetics
To investigate the mechanism of biosorption and potential rate controlling steps, pseudofirst-order and pseudosecond-order kinetic models have been applied to test experimental data and compare the kinetics of metal sorption at different putrescine concentrations. The pseudofirst-order and pseudosecond-order equations were used in this study. The experimental data of Cr(VI) removal by W. anomalus cells in the presence of different putrescine concentrations were analyzed using both adsorption kinetic models, and kinetic constants were calculated.
The Lagergren's first-order rate expression describes the adsorption rate based on the adsorption capacity. The pseudofirst-order equation is expressed as follows (Ho, 2006):
Pseudosecond-order equation is expressed as follows (Ho and McKay, 1999):
where t is the contact time (h); qt is the mass of the sorbed metal per unit mass of yeast cells at time (mg/g); qe is the amount of metal sorbed at equilibrium (mg/g); k1 (h−1) and k2 (g/[mg · h]) are the pseudofirst- and pseudosecond-order rate constants, respectively. qe and k2 were related to the slope and intercept, respectively, of the produced graph (Dhankhar and Hooda, 2011).
Biosorption characterization
Surface structure of biosorbents without and with different concentrations of putrescine was analyzed by an energy dispersive X-ray (EDX) spectrometer with the scanning electronic microscope (SEM, SEM-EDX; Qutanta 200).
Statistical analysis
Results were subjected to statistical calculations for mean comparison using XLSTAT software. One-way analysis of variance was used to test the significance of results followed by post hoc tests (Tukey HSD).
Results
Effect of putrescine on Cr(VI) removal
Measurement of Cr(VI) concentration in the medium using the diphenylcarbazide method has shown a removal percentage of 53.25% by control biomass of W. anomalus cells. With a concentration of 1 mM of putrescine, a slightly significant effect was noticed on the yeast cells' Cr(VI) removal potential, a removal of 59.19% was obtained. While a concentration of 10 mM of putrescine showed a highly significant effect on Cr(VI) biosorption (Fig. 1), Cr(VI) removal reached 80.50% (about 51% increase, compared with the control cells).

Effect of putrescine at 1 and 10 mM on chromium removal by Wickerhamomyces anomalus yeast cells.
Effect of putrescine on cell surface characteristics of W. anomalus
Effect on W. anomalus CSH
The results of hydrophobicity of untreated and putrescine-treated yeast cells are expressed qualitatively (θw) and quantitatively in terms of free energy of interaction ΔGiwi (Table 2). Qualitative hydrophobicity was analyzed from the water contact angles. According to Vogler (1998), hydrophobic surfaces exhibit a water contact angle >65°, while hydrophilic ones exhibit a water contact angle <65°. Accordingly, untreated W. anomalus cells present a hydrophilic surface (θw = 18.1° ± 2.35 < 65°). The yeast cells pretreatment with putrescine has lessened the hydrophilic character of microbial surface. A concentration of 1 mM has resulted a water contact angle of θw = 23.9° ± 0.86, while in the case of 10 mM of putrescine, a water contact angle of θw = 24.8° ± 2.89 was obtained.
Contact Angle Values (θw, θF, θD), Lifshitz–van der Waals (γLW), Electron-Donor (γ−), and Electron-Acceptor (γ+) Parameters, and Surface Energies (ΔGiwi) of Control and Putrescine-Treated Wickerhamomyces anomalus Cells
All contact angle measurements were computed and expressed as mean ± standard deviations from three independent measurements.
According to the approach of Van Oss et al. (1988), the absolute degree of hydrophilicity in terms of free energy of interaction was investigated (Elabed et al., 2011). Thus, the untreated surface exhibited hydrophilic character with a positive value of ΔGiwi (50.4 ± 1.74 mJ/m2). The ΔGiwi value of yeast cells decreased with putrescine treatment. Thus, values of 36.3 ± 2.48 mJ/m2 and 28.6 ± 2.58 mJ/m2 were obtained with 1 and 10 mM of putrescine, respectively.
Effect on electron donor/acceptor character
Lifshitz–van der Waals (γLW) and acid–base (γAB) surface tension components and electron donor (γ−) and electron acceptor (γ+) parameters of substrates and cells are presented in Table 2. Untreated W. anomalus cells present a dominant electron donor character with a high value of γ− (63.6 ± 1.47 mJ/m2), which is much greater than γ+ value (0.07 ± 0.01 mJ/m2) (Table 2). The treatment of yeast cells has resulted in the increase of the electron acceptor character with γ+ values of 0.13 ± 0.08 mJ/m2 and 1.73 ± 0.36 mJ/m2 with 1 and 10 mM of putrescine, respectively. It was also accompanied by a decrease of electron donor character (γ−) of microbial surface cells. γ− values of 54.5 ± 1.20 mJ/m2 and 50.8 ± 3.41 mJ/m2 were obtained with 1 and 10 mM of putrescine, respectively.
Sorption kinetics
Uptake of Cr(VI) at different putrescine concentrations is presented in Fig. 2. It shows that the metal uptake increases with the increase of putrescine concentration. The uptake at different concentrations of putrescine increases rapidly during the first hours and starts to reach equilibrium after 24 h. The maximum Cr uptake after 24 h was 13.31, 14.80, and 20.13 mg/g at 0, 1, and 10 mM of putrescine, respectively.

Uptake of Cr(VI) by W. anomalus yeast cells with different putrescine concentrations.
Adsorption kinetics was investigated to understand the mechanism of metal ion sorption onto the yeast cells. Sorption kinetics expressed as the solute removal rate is controlled by the residence time of the sorbate in the solid–solution interface (Pandey et al., 2010). In the present study, pseudofirst- and pseudosecond-order kinetic models were applied to the kinetic data. The obtained rate constant values k1 and k2 and correlation coefficient (R2) values are presented in Table 3. Figure 3 presents the results for the fit obtained using both models for different putrescine concentrations.
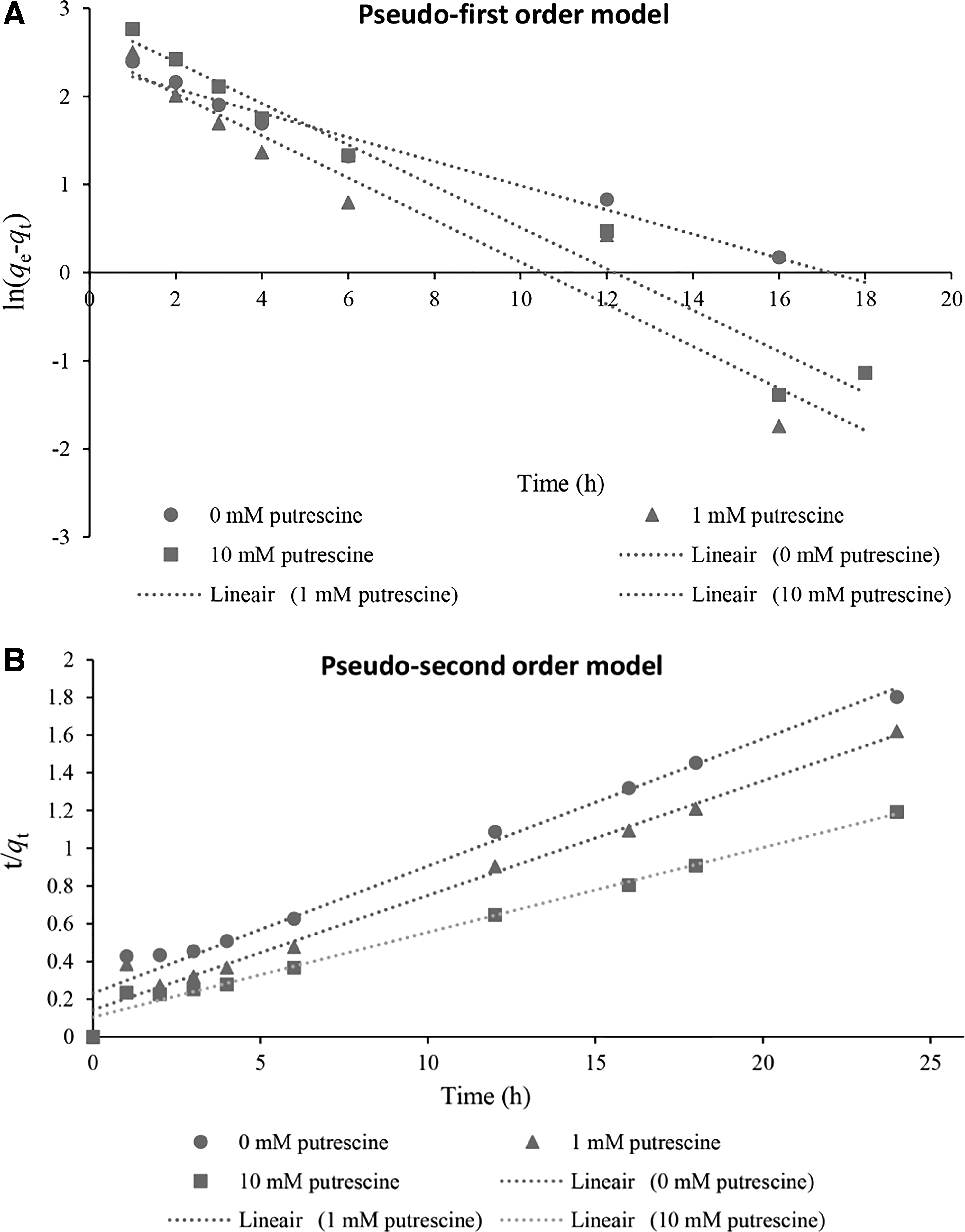
Kinetic data for Cr(VI) sorption by W. anomalus yeast cells and attempted fitting by two different models.
Constant Parameters of Pseudofirst- and Pseudosecond-Order Kinetic Models for Different Putrescine Concentrations for Initial Cr(VI) Concentration of 50 mg/L
SEM-EDX analysis
Elemental analysis by SEM-EDX of control and putrescine-pretreated yeast cells is shown in Table 4. The results have indicated that the treatment of yeast cells by putrescine has changed the chemical component of the yeast biomass. The major effect was noticed with a greater amount of nitrogen (44.15%) obtained with the addition of 10 mM, compared with untreated cells (32.19%).
The Element Percentage of the Control and Putrescine-Treated Wickerhamomyces anomalus Cells
Discussion
Effect of putrescine on Cr(VI) removal
Putrescine has shown a positive effect on Cr(VI) biosorption by W. anomalus. In a similar study, Tahri Joutey et al. (2014) reported that the effect of exogenous polyamines on the reduction of Cr(VI) by bacterial cells varies from one isolate to another and depends on the added amount of these polyamines. It is noteworthy that in the present study, no concentration of putrescine has incurred a negative effect on the chromium removal potential of yeast cells. This can be ascribed to the absence of competitive adsorption between chromium polyanions and putrescine acting as polycations (putrescine2+) mainly adsorbed to the negatively charged groups of the cell surface.
The polyamines are cationic molecules at the cytoplasmic pH and can act as enzymatic cofactors in the Cr(VI) reduction reaction. Several authors have reported that Cr(VI) removal is stimulated by cations, including Cu2+ in many bacteria (Van Oss, 1996; Ibrahim et al., 2011).
Extensive research has been reported on physical adsorption of heavy metals to the microbial cell wall. It was associated with the presence of van der Walls forces (Crowell, 1967). It was also related to the establishment of electrostatic interactions between heavy metal ions in solutions and fungal biomass cell walls (Kuyucak and Volesky, 1988) or bacterial cell surface (Aksu et al., 1992), as well as between cadmium ions and algal wall (Holan et al., 1993). Cell surface properties are dynamic parameters that can be influenced by different culture conditions (Holan et al., 1993; Gallardo-Moreno et al., 2004; Bahafid et al., 2011; Liu et al., 2011). Previous investigations have reported the influence of some compounds such as biosurfactants on the performance of microbial strains on cadmium (II) adsorption in relation to their effect on cell surface properties (Liu et al., 2011). Thus, the influence of putrescine on Cr(VI) biosorption could be via its modification of cell surface properties by different mechanisms. It may increase the amount of adsorption sites (Liu et al., 2011) and/or decrease membrane permeability (Kaczorek et al., 2010). It may also cause the release of some functional components able to adsorb heavy metals from the inside of cells, such as proteins and polysaccharides. This may change their cell wall characteristics and influence their bioremediation capacity (Guibaud et al., 2008).
Effect of putrescine on W. anomalus CSH
Numerous studies have reported that cell surface properties result from the unique chemical structure of the cell wall (Al-Tahhan et al., 2000). It is generally agreed that the most hydrophobic microbial surfaces tend to have greater protein/carbohydrate ratios (Holan et al., 1993; Van Der Mei et al., 1998; Guibaud et al., 2008).
Statistical analysis shows that a significant increase (43.25% compared with the control) of CSH of W. anomalus cells (θw = 18.1° ± 2.35, ΔGiwi = 50.4 ± 1.74 mJ/m2) was noticed with the addition of 10 mM of putrescine (θw = 28.6° ± 4.58, ΔGiwi = 28.6 ± 2.58 mJ/m2).
Our results demonstrate that the greatest Cr(VI) biosorption potential was obtained with the higher CSH, obtained with the addition of 10 mM of putrescine. Previous works aiming to establish a correlation between microbial cell surface properties and their bioremediation capacities have shown that the increase of CSH is a defense system to the toxicity of the added substrates (Zhang et al., 2010). They also showed that a higher CSH was advantageous for petroleum bioremediation and stress condition endurance (Al-Tahhan et al., 2000). It was also reported that the Serratia spp. with higher hydrophobicity has a better potential in beta-cypermethrin bioremediation (Zhang et al., 2010). Obuekwe et al. (2009) demonstrated that the most hydrophobic microbial strains were the best degraders of crude oil.
Most investigations aiming to identify the cell properties influencing bioremediation capacities mainly focus on CSH, especially to explain the interaction of microorganisms with hydrophobic organic compounds (Hamadi et al., 2005; Płaza et al., 2005; Kaczorek et al., 2010; Asri et al., 2017). Whereas in the case of charged metal ion bioremediation, CSH should not be the only selection criteria of performant bacterial strains, the energy of acid–base interactions can have a great significance (Van Oss, 1996).
Effect on electron donor/acceptor character
Microbial acid–base components of surface free energy may have an effect on bioremediation potential, especially against charged molecules in environmental applications (Liu et al., 2011). They may play a key role in the microbial cell interactions with heavy metals, soil surfaces, and hydrocarbons. Van Oss (1996) reported that the energy of acid–base interactions can be twice greater than that due to hydrophobic interactions, but their role in the phenomenon of bioremediation is rarely studied.
W. anomalus cells present a dominant electron donor character. These results are in agreement with a previous study presented by Van Der Mei et al. (1998), which showed that microbial cell surfaces are predominately electron-donating, while electron-accepting cell surfaces can be rarely found. This was also in agreement with the results reported by Mercier-Bonin et al. (2004) and Vichi et al. (2010) showing that Saccharomyces cerevisiae cells are exhibiting a great Lewis basic character. Microbial surface is essentially composed of polysaccharides, proteins, and lipids. This composition offers a dominant electron donor character due to the presence of many negatively charged functional groups such as carboxylate, hydroxyl, thiol, sulfonate, phosphate, amino, and imidazole groups (Mercier-Bonin et al., 2004; Liu et al., 2011). For yeast strains, Vichi et al. (2010) reported that yeast cells exhibit negatively charged surfaces due to the presence of phosphate groups in their cell wall, as suggested before by Amory and Rouxhet (1988). A previous Fourier-transformed infrared analysis of W. anomalus has confirmed the presence of phosphate groups on its cell surface (Bahafid et al., 2011).
A significant modification of cell surface donor/acceptor character could be observed with the addition of 10 mM of putrescine. At this concentration, the greatest chromium removal potential was noticed with the greatest value of electron acceptor character γ+. The acid component can markedly influence the capacity of Cr(VI) adsorption of yeast cells. These findings are consistent with previous articles reporting that the acid character of microbial cell walls favors the adsorption of chromium anions (Gupta and Rastogi, 2009).
Sorption kinetics
The correlation coefficient (R2) values indicate that at different putrescine concentrations, the pseudosecond-order model fits better than the pseudofirst-order model and provides the best correlation of the data for chromium uptake by W. anomalus cells. This model is based on the assumption that the reaction rate is controlled by surface reaction, with chemical sorption involving valency forces, through sharing or electron exchange between yeast cells and metal ions (Aksu and Dönmez, 2003; Quintelas et al., 2013). For the pseudofirst-order model, it assumes that the reaction rate is limited by one process or mechanism on a single class of sorbing sites and that all sites are of the time-dependent type (Fonseca et al., 2009). The calculated sorption capacity (qe) values of the pseudosecond-order model agreed very well with the experimental (qe) values. These results are in agreement with previous results reporting that metal sorption by microbial cells is generally described by the pseudosecond-order model (Ma et al., 2015; Rashid et al., 2016).
SEM-EDX analysis
SEM-EDX analysis is considered one of the useful tools aiming to evaluate the elemental characteristics of microorganism surface. Elemental analysis of control and putrescine-pretreated yeast (Table 3) indicated that the pretreatment of yeast cells by putrescine changed the chemical component of the yeast biomass. A greater amount of nitrogen was obtained with the addition of 10 mM. Thus, the adsorption of putrescine molecules on the surface of yeast wall increases the amount of amino groups, which may explain the increase of the acceptor character and their important chromium removal capacity. Indeed, previous work indicated the important role played by amino groups in the biosorption of heavy metals (Gong et al., 2005). It was also reported that the acetylation of amino and hydroxyl groups of Rhizopus nigricans biomass reduces chromium ion biosorption (Bai and Abraham, 2002).
Conclusion
Results obtained by contact angle measurements show that the physicochemical properties, including hydrophobicity and donor/acceptor electron properties of yeast cell surface, were markedly affected by the treatment by putrescine. Putrescine enhanced Cr(VI) removal by W. anomalus, accompanied by an increased hydrophobicity and electron acceptor character and a decreased electron donor character of surface cells. The SEM-EDX analysis confirmed that treatment by putrescine changes cell surface properties. Therefore, the element percentage of the biomass changed with a high concentration of nitrogen on yeast cell wall. The addition of 10 mM of putrescine enhanced the adsorption capacity of Cr(VI) by W. anomalus. It showed a high effect on Cr(VI) biosorption, it reached 80,50% compared with 53.25% by control biomass. The data of sorption experiments showed that the pseudosecond-order kinetic model best explained Cr(VI) sorption onto W. anomalus cells.
This work highlights the possibility of wastewater treatment enhancement, especially chromium uptake, by modifying cell surface characteristics (enhancing hydrophobicity and electron acceptor character).
Footnotes
Acknowledgments
The authors thankfully acknowledge the financial and scientific support of Microbial Biotechnology Laboratory and the “Centre Universitaire d'Interface” SMBA University, Fez, Morocco.
Author Disclosure Statement
No competing financial interests exist.