
Abstract
Abstract
Membrane-based pretreatment can recover organic matter from the sewage and further reduce energy consumption, and the remaining wastewater often contains low C/N ratio. Anoxic/oxic membrane bioreactor (MBR) is considered as the effective way to remove nutrients from low C/N ratio wastewater. However, information on microbial community related to this process is limited. In this study, performance and bacterial communities in two systems, anoxic/oxic MBR, and anoxic/oxic conventional activated sludge (CAS), were compared after being fed with the organic matter-recovered sewage. The MBR system performed better than the CAS system at nitrogen removal when operated at all loading rates (5–12.5 L/day) and recirculation ratio (1 and 2). Higher bacterial diversity and a larger number of novel operational taxonomic units were found in the MBR system at a loading rate of 10 L/day. Proteobacteria clones were most abundant, followed by Bacteroidetes and Actinobacteria clones in all reactors, but the clone ratio within each phylum and the class of Proteobacteria were different. Larger differences were observed at the genus level in Proteobacteria in both systems. In the denitrification reactors, Tolumonas, Dokdonella, and Thauera genera were identified in the CAS system and Tolumonas, Accumulibacter, and Dechloromonas genera were identified in the MBR system. More genera, including Paracoccus, Woodsholea, Dechloromonas, Azospira, and Inhella, were identified in the nitrification reactor of the MBR system than the CAS system, in which only the genera, Leptothrix and Hirschia, were detected. These results identified different bacterial communities responsible for the stable performance of CAS and MBR systems.
Introduction
Activated sludge processes is the leading method for the treatment of domestic and industrial wastewater worldwide. However, its high costs and high levels of energy consumption, CO2 emission, and sludge generation, are some disadvantages of the conventional activated sludge (CAS) process (McCarty et al., 2011). This, together with the fact that the treatment and disposal of excess sludge accounts for 40–60% of the total operational cost in many sewage plants (Zessner et al., 2010), means that CAS is often considered to be cost ineffective, based on life cycle cost analyses.
Recently, a novel sewage treatment concept, namely the recovery of organic matter from wastewater before treatment, has shown promise. This process could further permit energy recovery from recovered organic matter and water reuse from sewage (McCarty et al., 2011; Verstraete and Vlaeminck, 2011). Many reports have confirmed that membrane-based pretreatment, such as membrane filtration, coagulation, and adsorption, can effectively recover organic matter from the sewage and further reduce energy consumption and sludge volume during the treatment process, so that the remaining wastewater contains a significantly reduced amount of organic matter (Gong et al., 2015; Jafari et al., 2015).
After organic matter has been removed, the remaining wastewater often contains a low C/N ratio (total organic carbon [TOC]/total nitrogen [TN], often lower than 8.0), and nitrogen removal is key to cleansing this wastewater (Sun et al., 2010). At present, the commonly used methods for biological nitrogen removal involve successive nitrification and denitrification, which are carried out by different bacterial groups. During the nitrification process, autotrophic ammonia-oxidizing bacteria (AOB) and nitrite-oxidizing bacteria are responsible for the aerobic oxidation of ammonium to nitrite and nitrate (Kim and Kim, 2006). The resulting nitrate and nitrite are then converted into nitrogen gas by heterotrophic bacteria, in the presence of an organic substrate during the denitrification process (Wiesmann, 1994).
When treating wastewater with a low C/N ratio, nitrogen removal is often limited by a lack of organic carbon. Higher recirculation rates and the addition of external organic carbon (such as acetic acid and methanol) are required to achieve complete denitrification, which results in instability and a high cost and energy consumption of the CAS system (Sun et al., 2010). Therefore, a widely used predenitrification process could effectively solve this problem by utilizing organic matter from the inflow of low C/N ratio wastewater as a carbon source. However, it is difficult to retain enough slow-growing nitrifying bacteria, whose highest growth rates are 0.014–0.064 h−1 in a CAS reactor (Prosser, 1989).
Membrane bioreactor (MBR) technology is a combination of the activated sludge process and membrane separation (Marrot et al., 2004). The MBR process offers some advantages over the CAS system, including lower cost, better effluent quality, a lower food/microorganism (F/M) ratio, less sludge production, longer sludge retention time (SRT), and a smaller footprint (Visvanathan et al., 2000; Marrot et al., 2004). Many studies have reported good nitrification performance for low C/N ratio wastewater using MBR processing, due to the large biomass of nitrifying bacteria and longer SRTs (Bracklow et al., 2007; Tan and Ng, 2008; Xia et al., 2008).
Nitrification performance depends on the balance of different microorganisms responding to environmental conditions in a given system. An understanding of bacterial communities in specific systems is beneficial for improving their performance. To date, most studies have focused on differences in the operational performance between CAS and MBR systems under different conditions. However, there have been few reports investigating and comparing the diversity of microbial communities between the two systems (Silva et al., 2010; Wan et al., 2011; Chiellini et al., 2013; Saunders et al., 2013). To our knowledge, information on microbial communities about the treatment of wastewater with a low C/N ratio is even more limited, with only several published reports (Ma et al., 2013). In light of these findings, the goal of the current study was to evaluate and compare the performance and bacterial communities involved in nitrogen removal, in MBR and CAS systems fed with low C/N ratio wastewater, which simulates sewage after the recovery of organic matter.
Materials and Methods
Operation of CAS and MBR systems
Laboratory-scale anoxic/oxic CAS and MBR systems were constructed, each with a predenitrification reactor and a nitrification reactor. Activated sludge from the aeration section of the Hangkonggan Sewage Treatment Plant (Chengdu, China) was used as the inoculum to set up the two systems. The percentage of total solids (TS), volatile total solids (VTS), and mixed liquor suspended solids (MLSS) in the inoculated sludge were 3.1% and 2.6%, 5.3 g/L, respectively.
As shown in Fig. 1, the structures of the denitrification reactor in the CAS system (designated as CND) and denitrification reactor in the MBR system (designated as MND) were absolutely identical. They each consisted of a tower reactor made of acrylic resin, each with a working volume of 0.45 L. Approximately 0.45 L of sludge was inoculated into each reactor, which was then fed with low C/N ratio wastewater. The composition of the synthetic wastewater was as follows (mg/L): glucose, 65; NH4COOCH3, 88; NaHCO3, 35; CH3COONa, 10; beef extract, 2; and peptone, 4. The concentrations of TOC, COD, and NH4+ in the wastewater were 65.2, 116, and 20 mg/L, respectively. The liquid in each reactor was circulated at a constant flow rate of 0.1 L/min using a roller pump.
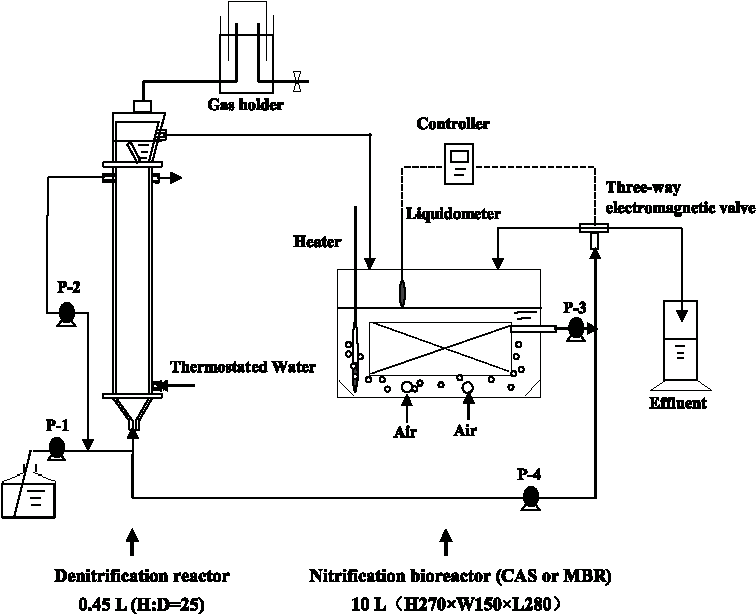
Schematic diagram of denitrification and nitrification processes. CAS, conventional activated sludge; MBR, membrane bioreactor.
The structures of the nitrification reactor in the CAS system (designated as CN) and denitrification reactor in the MBR system (designated as MN) were identical, except that 2 U of hollow fiber membrane (Sterapore PE, SURM0334L; Mitsubishi Rayon, Tokyo, Japan) was submerged in parallel in the MN reactor. Both reactors were made of acrylic resin, each with a total volume of 10 L and a working volume of 6 L. Six liters of sludge was inoculated into each reactor. Air was supplied to the MN and CN reactors through a blower at an aeration rate of 1.0 L/min and the air was dispersed through four glass ball filters set into the bottom of each reactor. In the MN reactor, the four glass ball filters were located beneath the membrane module.
The temperature in all reactors was maintained at 27°C. In each system, the denitrification and nitrification reactors were connected and the effluent from the nitrification reactor was recycled into the denitrification reactor. Using a recirculation ratio of 1, the effects of loading rate (from 5 to 12.5 L/day) on effluent quality were investigated. The recirculation ratio was then increased to 2, at a loading rate of 12.5 L/day. The MBR systems were operated for ∼5 months without any sludge purge.
Chemical analysis
Samples from the four reactors were collected periodically for analysis of pH, TS, VTS, suspended solids (SS), volatile suspended solids (VSS), MLSS, TOC, inorganic carbon (IC), NH4+, and NO3−. The pH was measured using a pH meter (HM-25G; TOADKK, Tokyo, Japan). TS, VTS, SS, and VSS concentrations were analyzed using standard methods (APHA, 1998). MLSS measurements were performed gravimetrically. Samples for TOC, NH4+, and NO3− analysis were membrane filtered before analysis (0.45 μm). TOC was measured using a TOC analyzer (TOC-V CSH; Shimadzu, Kyoto, Japan). The concentration of NH4+ was measured using an ion chromatography system (ICS-1500; Dionex, Sunnyvale, CA) equipped with a CS-12A column and a CG-12A guard column. NO3− concentration was measured using an ion chromatography system (ICS-1100; Dionex) equipped with an AS-12A column and an AG-12A guard column.
Construction of 16S rRNA gene clone libraries
To investigate the bacterial communities involved in denitrification and nitrification in the MBR and CAS systems, sludge samples were collected at a loading rate of 10 L/day. These samples were used to construct four 16S rRNA gene libraries: CDN, CN, MDN, and MN. DNA was extracted from samples as previously described (Shigematsu et al., 2006a). Bacterial 16S rRNA genes were PCR amplified with the universal primers, Eu27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1490R (5′-GGTTACCTTGTTACGACTT-3′). PCR conditions were as follows: 95°C for 5 min; followed by 20 cycles at 94°C for 1 min, 50°C for 45 s, and 72°C for 90 s. The amplified 16S rRNA gene fragments were then cloned into the pMD19 T-vector plasmid (TaKaRa, Kusatsu, Japan) using a DNA Ligation Kit (TaKaRa), according to the manufacturer's protocol.
The recombined plasmids were then transformed into Escherichia coli DH5α cells (TaKaRa) and plated on Luria-Bertani agar plates containing 100 μg/mL ampicillin, 40 μg/mL X-gal (TaKaRa), and 24 μg/mL isopropyl β-D-1-thiogalactopyranoside (TaKaRa). One hundred white clones from each library were randomly selected for plasmid extraction and the presence of the 16S rRNA gene was verified by PCR. The positive clones were classified into different phenotypic groups, according to the pattern of restriction fragment length polymorphism digestion with RsaI and MspI enzymes (New England BioLabs, Ipswich, MA). Representative clones from each group were then sequenced.
Sequence analysis of libraries
A chimera check was performed using Bellerophon webservices, as described by Huber et al., and those sequences showing >97% similarity were grouped into one operational taxonomic unit (OTU) using Mothur software (Huber et al., 2004; Schloss et al., 2009). A similarity search was performed using the GenBank database and the BLAST search program. OTU sequences were then aligned using the Clustal_X program (Thompson et al., 1997) and phylogenetic trees were constructed based on the neighbor-joining method using Molecular Evolutionary Genetics Analysis software (Tamura et al., 2007). The robustness of the phylogeny was tested by bootstrap analysis with 1,000 iterations.
Statistical analysis of libraries
Statistical analysis of each library was performed using Mothur software (Schloss et al., 2009). Chao 1 and Abundance-Based Coverage Estimator (ACE) estimators for species richness estimation and the Shannon and Simpson indices for diversity estimation were calculated with 95% confidence intervals, with a sequence similarity cutoff set at 3%. Good's coverage index, which describes the extent to which the sampled sequences in a given library represent the total population, was calculated. The Boneh index was calculated when the sample size was set at 50.
Nucleotide sequence accession numbers
The GenBank accession numbers for the bacterial OTU sequences obtained in this study are KP054129 to KP054242.
Results
Performance of CAS and MBR systems in response to different operational strategies
The CAS and MBR systems fed with low C/N ratio wastewater were operated in parallel to investigate and compare their denitrification and nitrification performance at different loading rates.
Performance of denitrification process
Figure 2A shows the effects of loading rate on the denitrification process of two systems, with a recirculation ratio of 1. The loading rate was increased from 5 to 12.5 L/day, which corresponded to hydraulic retention time (HRT) values increasing from 1.08 to 0.432 h. The MDN reactor showed a higher TOC removal efficiency (70.4–76.7%) than the CDN reactor (50.1–57.7%) and the NO3−N-removal efficiency was maintained at greater than 97% for all loading rates studied. NO3−N-removal efficiency in the CDN reactor slightly improved from 86.3% to 89.7% (with an effluent concentration increase from 2.2 to 1.53 mg/L) when the loading rate increased from 5 to 12.5 L/day. The MLSS concentration in both reactors increased when loading rates increased from 5 to 7.5 L/day, but then began to decrease when the loading rate increased up to and above 10 L/day. Less sludge was produced in the MDN reactor than in the CDN reactor and this was most apparent at a loading rate of 10 L/day (2.7 and 4.7 g/L MLSS, respectively).

Performance of denitrification
Performance of nitrification process
Figure 2B shows the effects of loading rate on the nitrification process of two systems, with a recirculation ratio of 1. HRT increased from 14.4 to 5.76 h, corresponding to a loading rate increase from 5 to 12.5 L/day. The MLSS concentration in the CN reactor exhibited a trend of first increasing (from 5 to 7.5 L/day) and then decreasing (from 7.5 to 12.5 L/day). In contrast, the MN reactor was able to keep a continuous growth of sludge and maintain a high MLSS concentration, without sludge discharge, over a 5-month period, even at a loading rate of 12.5 L/day. TOC removal was lower in the MN reactor (59.8–78.8%) than in the CN reactor (71.4–81.2%). Almost all of the NH4+-N was removed from the MN reactor at all loading rates, whereas more than 92% of the NH4+-N was removed from the CN reactor. Moreover, a higher removal efficiency was observed at lower loading rates in the CN reactor.
Performance of whole denitrification/nitrification process
Figure 2C shows the effects of loading rate on the whole process of two systems, with a recirculation ratio of 1. There were no clear differences in TOC or NH4+-N removal between the CAS and MBR systems and low NH4+-N levels were detected in the effluents, with less than 1 mg/L at each loading rate. However, the removal efficiency of TN in the MBR system was 59.7–61%, indicating that it performed better than the CAS system, which showed a TN removal efficiency of 52.5–53.9%.
To improve the TN removal efficiency, the recirculation ratio of the two systems was increased from 1 to 2. In the MBR system, the total TOC removal efficiency subsequently increased and remained above 93% and the TN removal efficiency increased to 70%. However, in the CAS system, the MLSS in the CN reactor sharply decreased to 600 mg/L over 2 days and a stable treatment process could not be maintained at a recirculation ratio of 2 (data not shown). These results showed that, in the MBR system, nitrogen removal could be performed at a much lower HRTs of 0.288 and 3.84 h for denitrification and nitrification, respectively. These differences in performance may be due to different responses of the microbial community in each system to the higher recirculation ratio, in that the microbial community in the MBR system may have a better adaptive capacity.
Statistical analysis of 16S rRNA gene libraries from CAS and MBR
To identify differences in the microbial content of the CAS and MBR systems, the bacterial communities in each reactor, at a loading rate of 10 L/day, were investigated and compared using a 16S rRNA gene library. Two libraries from the CAS system (libraries CDN and CN) and two from the MBR system (libraries MDN and MN) were constructed. A total of 312 bacterial sequences were obtained from the four libraries (161 sequences from the CAS system and 151 from the MBR system). As shown in Supplementary Table S1, over 80% coverage for all four libraries indicated that the sequences obtained in each library were representative of the whole population. The Boneh indices predicted that three to four new OTUs would be identified if 50 or more bacterial clones were analyzed from each of these libraries. Library MN had much greater bacterial richness and diversity than the other libraries, as indicated by the Shannon index, Chao 1, and ACE values.
Sequence analysis revealed 22 OTUs from 73 clones (library CDN), 22 OTUs from 72 clones (library MDN), 25 OTUs from 88 clones (library CN), and 34 OTUs from 79 clones (library MN) in the different libraries (Supplementary Table S1). Most of the OTUs from all four libraries were identified as already known species or were affiliated with as-yet uncultivated species in public databases with ≥97% sequence similarity (Supplementary Table S1). A greater number of uncultured OTUs were obtained from library MN than from the other libraries. In addition, some novel OTUs, which had not been described in previous analyses of either CAS or MBR communities, were present in each library. Moreover, a greater number of novel OTUs were identified in the MBR system (libraries MDN and MN) than in the CAS system.
Composition of the bacterial communities at the phylum level
As shown in Supplementary Table S2 and Figs. 3–7, the composition of bacterial communities at the phylum level differed between libraries from the CAS and MBR systems, although the phylum, Proteobacteria, was predominant in all libraries.
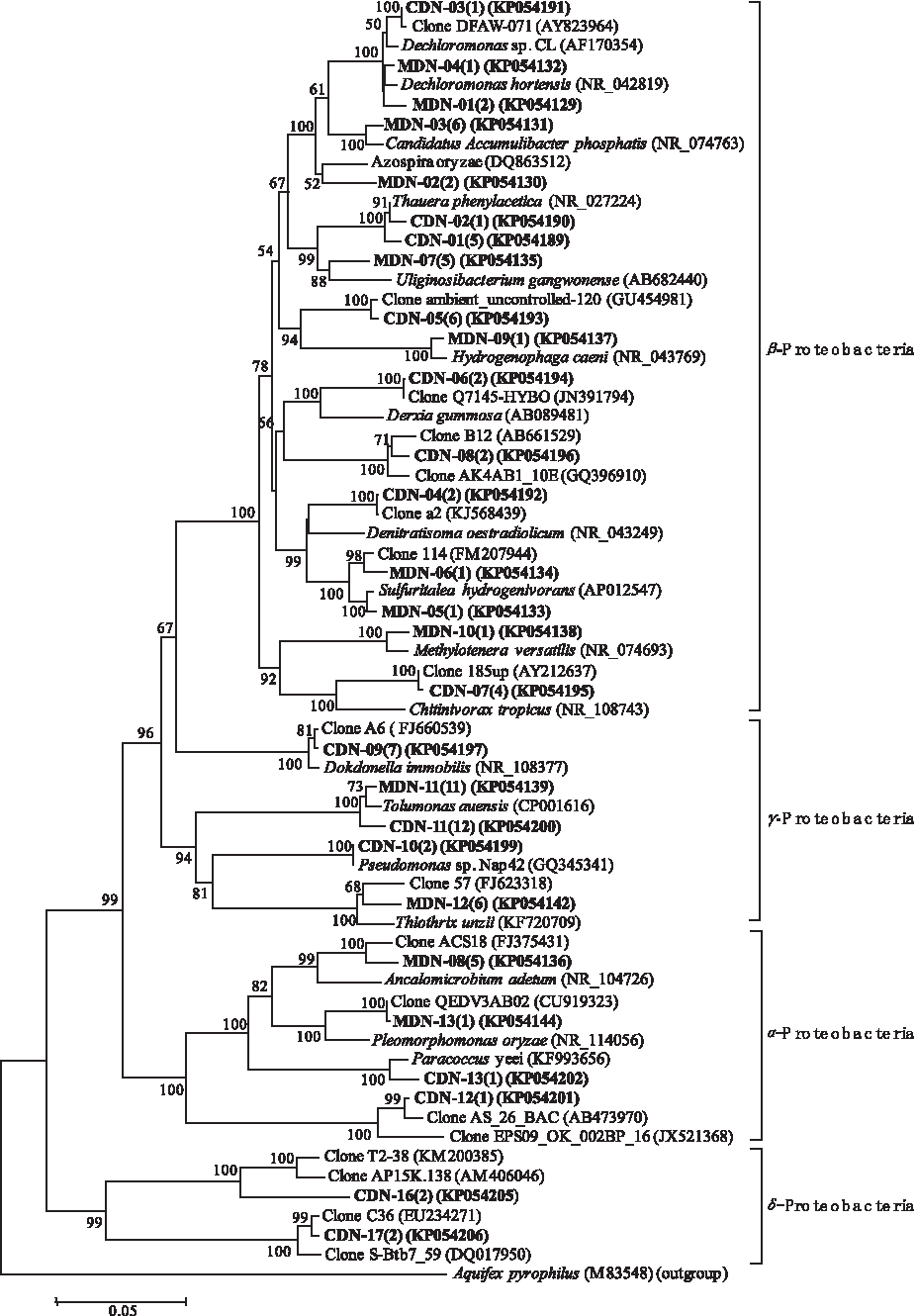
Phylogenetic affiliation of denitrifying bacteria belonging to Proteobacteria phylum retrieved from the MBR and CAS systems. Evolutionary distance was constructed using the neighbor-joining method. The number of clones with identical sequences is shown in parentheses. Clones obtained in this study are in boldface.
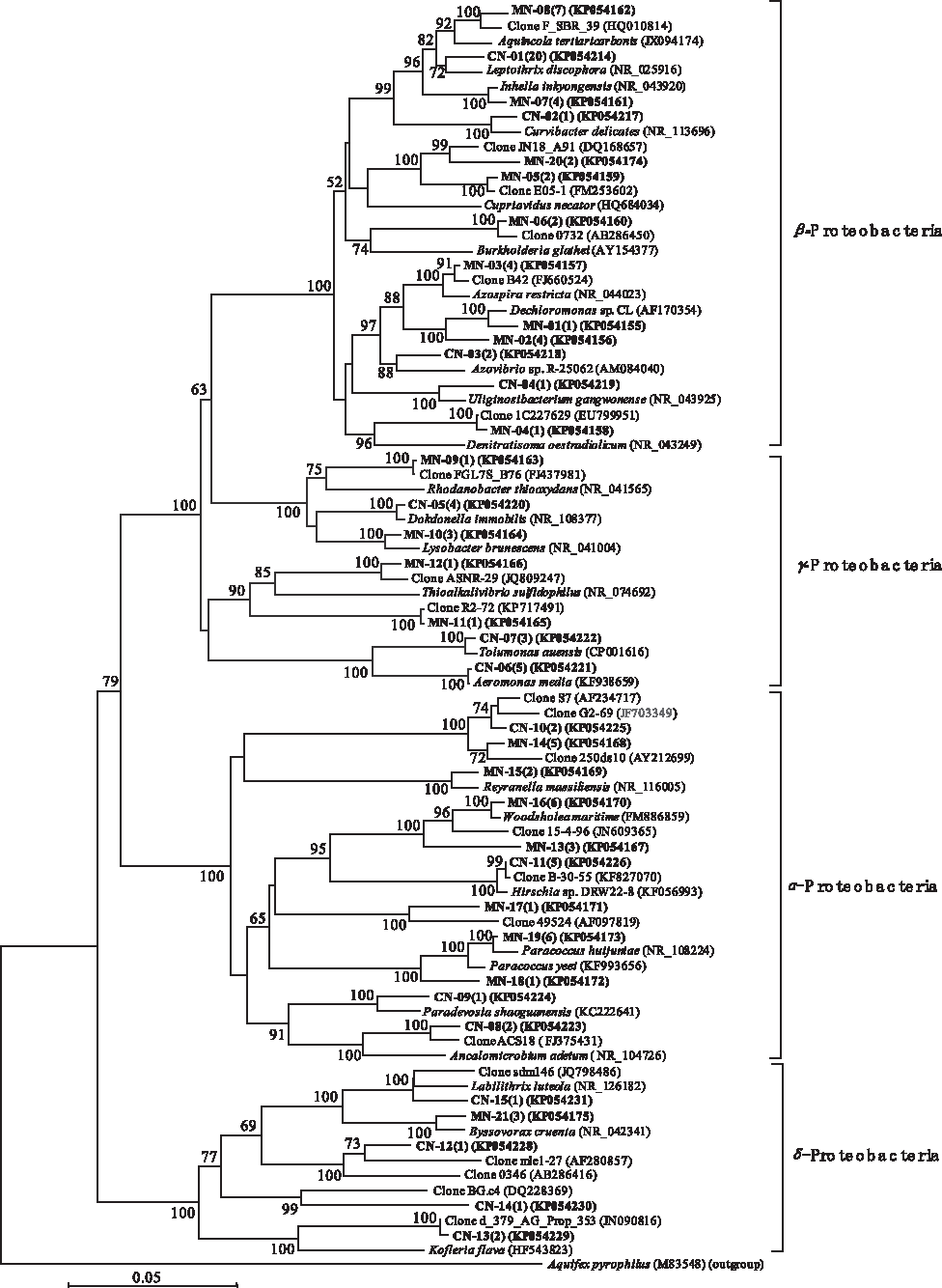
Phylogenetic affiliation of nitrifying bacteria belonging to Proteobacteria phylum retrieved from the MBR and CAS systems. Evolutionary distance was constructed using the neighbor-joining method. The number of clones with identical sequences is shown in parentheses. Clones obtained in this study are in boldface.
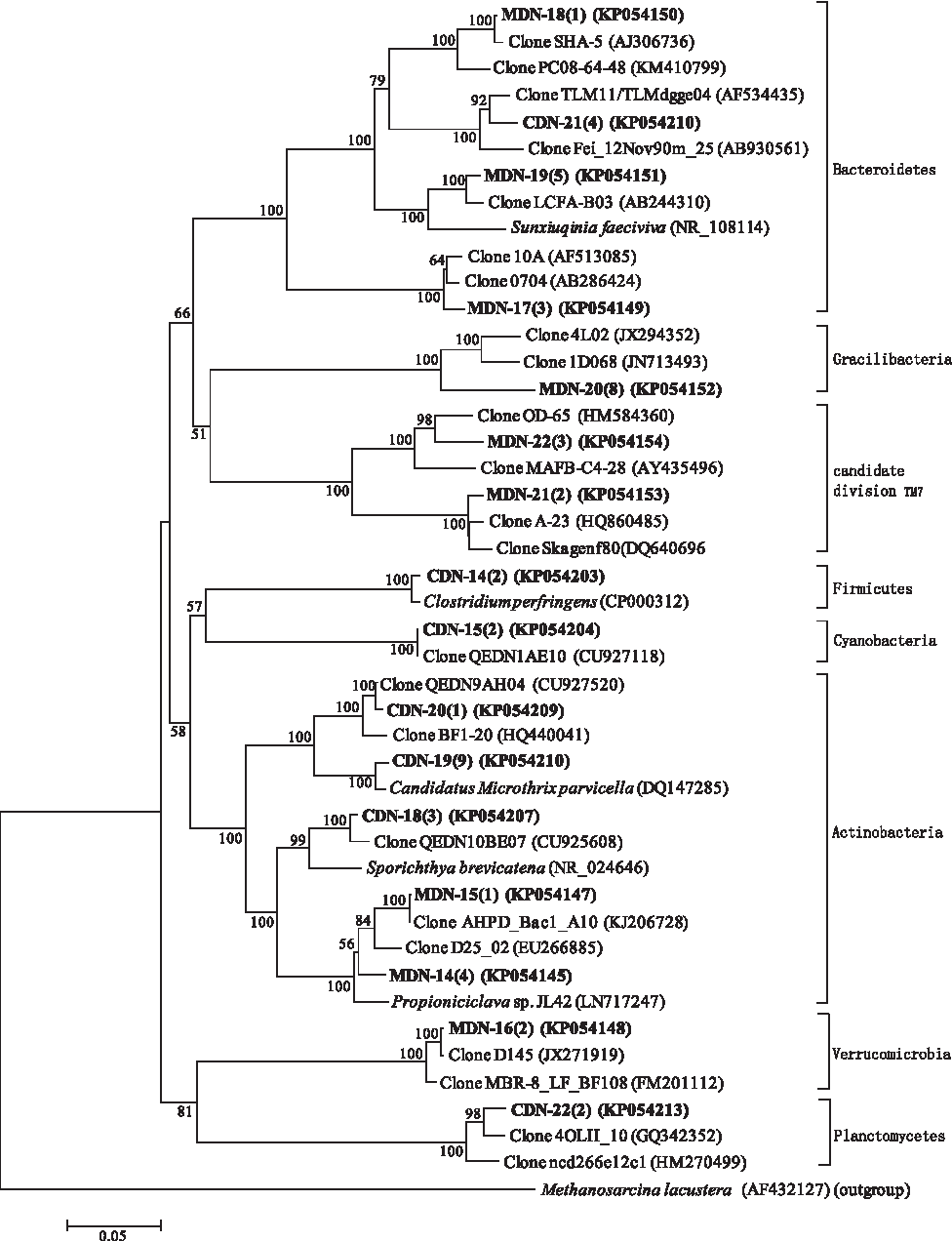
Phylogenetic affiliation of denitrifying bacteria belonging to other phyla retrieved from the MBR and CAS systems. Evolutionary distance was constructed using the neighbor-joining method. The number of clones with identical sequences is shown in parentheses. Clones obtained in this study are in boldface.
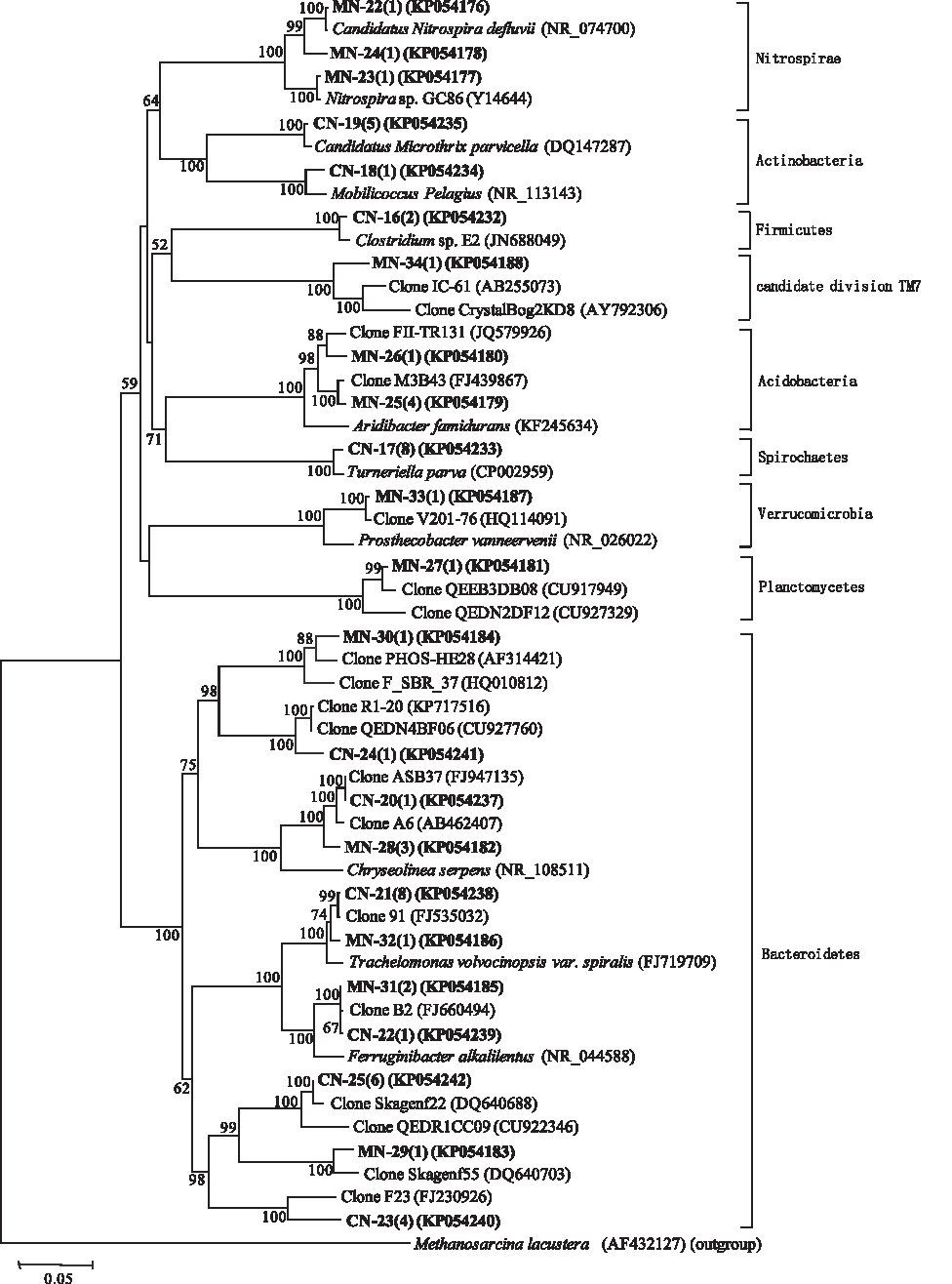
Phylogenetic affiliation of nitrifying bacteria belonging to other phyla retrieved from the MBR and CAS systems. Evolutionary distance was constructed using the neighbor-joining method. The number of clones with identical sequences are shown in parentheses. Clones obtained in this study are in boldface.
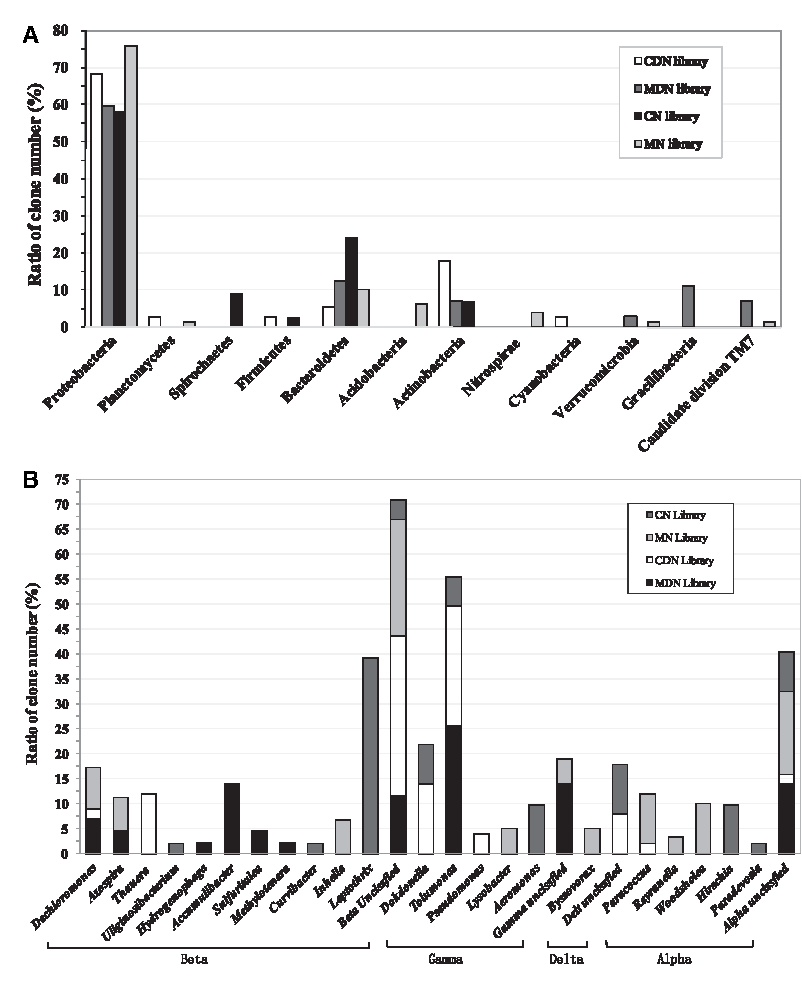
Proportion of 16S rRNA gene clones from the MBR and CAS processes within different phyla
For CDN and MDN, six major taxa (22 OTUs) were identified in both libraries. In library CDN, 68.5% of sequences were related to the phylum, Proteobacteria, and most clones were identified as belonging to either class β-Proteobacteria (31.5%) or γ-Proteobacteria (28.8%). The remaining sequences were affiliated with the phyla, Actinobacteria (17.8%), Bacteroidetes (5.5%), Planctomycetes (2.7%), Firmicutes (2.7%), and Cyanobacteria (2.7%). In library MDN, 59.7% of clones were identified as Proteobacteria species. The most abundant clones were classified as belonging to classes β- and γ-Proteobacteria, which accounted for 27.8% and 23.6% of clones, respectively. No δ-Proteobacterial clones were present in library MDN, whereas a few were found in library CDN. The other phyla in library MDN were identified as Bacteroidetes (12.5%), Gracilibacteria (12.5%), Actinobacteria (6.9%), Candidate division TM7 (6.9%), and Verrucomicrobia (2.8%). In conclusion, there were only three common phyla present in both libraries (Proteobacteria, Actinobacteria, and Bacteroidetes) and they were present at different clone ratios.
For the nitrification process, library CN had less bacterial diversity than library MN. Five phyla (25 OTUs) and seven phyla (34 OTUs) were identified in libraries CN and MN, respectively. In library CN, the clones were affiliated with the phyla, Proteobacteria (58%), Bacteroidetes (23.9%), Spirochaetes (9.1%), Actinobacteria (6.8%), and Firmicutes (2.3%). Class α-, β-, γ-, and δ-Proteobacteria accounted for 11.4%, 27.2%, 13.6%, and 5.7% of the total clones, respectively. In library MN, Proteobacterial clones were predominant (75.9%) and most were identified as α-Proteobacteria (30.4%) or β-Proteobacteria (34.2%). The remaining sequences were affiliated with the phyla, Bacteroidetes (10.1%), Acidobacteria (6.3%), Nitrospirae (3.8%), Candidate division TM7 (1.3%), Planctomycetes (1.3%), and Verrucomicrobia (1.3%). Proteobacteria and Bacteroidetes species were common to both libraries.
Composition of bacterial communities at genus level
Larger differences were detected when the genera belonging to Proteobacteria were compared between the libraries (Figs. 3, 4, and 7B). For the denitrification process, only the genera, Tolumonas and Dechloromonas appeared in both libraries. In library CDN, three γ-Proteobacterial genera (Tolumonas, Dokdonella, and Pseudomonas), two β-Proteobacterial genera (Dechloromonas and Thauera), and one α-Proteobacterial genus (Paracoccus) were identified and the clones were predominantly affiliated with Tolumonas auensis. T. auensis was also identified as the dominant bacteria in library MDN. Other genera from the class, β-Proteobacteria, were found, including Dechloromonas, Azospira, Hydrogenophaga, Accumulibacter, Sulfuritalea, and Methylotenera. In addition, many uncultured clones were obtained from both libraries, most of which were affiliated with β-Proteobacteria in library CDN, whereas a similar clone ratio of α-, β-, and γ-Proteobacteria was present in library MDN. The phylogenetic analysis presented in Fig. 3 shows that there was low sequence similarity between the uncultured clones from library CDN and library MDN.
For the nitrification process, no known genus belonging to Proteobacteria coexisted in the two libraries (Fig. 4). In library CN, the most dominant sequence was from Leptothrix spp., affiliated with β-Proteobacteria, accounting for 39.2% of the total Proteobacterial clones. Low populations of other genera were found, including Uliginosibacterium and Curvibacter, belonging to the class, β-Proteobacteria; Dokdonella, Tolumonas, and Aeromonas, belonging to the class, γ-Proteobacteria; and Hirschia and Paradevosia, belonging to the class, α-Proteobacteria. In library MN, three α-Proteobacterial genera (Paracoccus, Reyranella, and Woodsholea), three β-Proteobacterial genera (Dechloromonas, Azospira, and Inhella), one γ-Proteobacterial genus (Lysobacter), and one δ-Proteobacterial genus (Byssovorax) were identified. In addition, a greater number of uncultured clones were found in library MN (40% of the Proteobacterial clones) than library CN and these were affiliated with α- and β-Proteobacteria. Only 20.5% of the uncultured clones present in library CN were Proteobacterial clones.
Figures 5 and 6 show the phylogenetic relationship of the bacterial communities belonging to other phyla. There was one genus (Propioniciclava belonging to the phylum, Actinobacteria) and two genera (Clostridium belonging to the phylum, Firmicutes and Microthrix belonging to the phylum, Actinobacteria) identified in MDN and CDN libraries, respectively. The remaining denitrifying bacteria identified in the two libraries were uncultured clones. In library CN, two Actinobacterial genera (Microthrix and Mobilicoccus), one Firmicutes genus (Clostridium), one Spirochaetes genus (Turneriella), and one Bacteroidetes genus (Trachelomonas) were identified. In library MN, one Bacteroidetes genus (Trachelomonas), and two Nitrospirae genera (Nitrospira) were found. In addition, most of the uncultured clones identified in the two libraries were affiliated to the phylum, Bacteroidetes.
Discussion
Laboratory-scale CAS and MBR systems fed with low C/N ratio wastewater were operated in parallel to compare their performance and the bacterial communities they harbor, during denitrification and nitrification processes. The results showed that the MBR system was more efficient at TN removal than the CAS system when they were operated under different loading rates (from 5 to 12.5 L/day), at a recirculation ratio of 1. However, there was no clear difference in the efficiencies of TOC and NH4+ removal between the two systems.
The CAS system was not able to remain stable and washout occurred when the recirculation ratio was increased to 2, in an attempt to improve TN removal. In addition, the MBR in this study was continuously operated for 5 months without any sludge discharge, which could greatly reduce the operational costs of this system. The improved performance of the MBR system over the CAS system may be attributed to the membrane separation step used in the MBR, which may prevent slow-growing nitrifiers from being washed out and thus, allow a microbial community to form that is completely different from the CAS system, resulting in a longer SRT and a lower F/M ratio (Wagner et al., 2002).
The 16S rRNA gene library analysis showed that there were distinctly different bacterial communities in the CAS and MBR systems, at a loading rate of 10 L/day and a recirculation ratio of 1. Higher bacterial diversity and a greater number of novel OTUs were identified in the MDN and MN reactors than the other reactors (Supplementary Table S1). Proteobacteria were most abundant in all reactors, followed by Bacteroidetes and Actinobacteria, which is similar to the bacterial communities reported previously (Loy et al., 2002; Wagner et al., 2002). This suggested that these phyla may be the major denitrifying and nitrifying bacteria. However, the abundance of clones representing the different classes of Proteobacteria and other phyla was different in each library. These discrepancies may be due to differences in the SRT and F/M ratios between the two systems. According to the resource ratio theory, R-strategist bacteria (with low substrate affinities and high growth rates) can be competitive in the CAS system, whereas K-strategist bacteria (with high substrate affinities and low growth rates) would be favored in the MBR system (Curtis et al., 2003).
At the genus level in the phylum, Proteobacteria, significant differences were found between the two nitrification reactors. Only the genera, Tolumonas and Dechloromonas, appeared in both libraries. T. auensis, the dominant bacterium in the CDN and MDN libraries, has been reported to have the ability to produce toluene from phenylalanine and other phenyl precursors, as well as phenol from tyrosine (Fischer-Romero et al., 1996). It was first identified in a denitrification system, but its function is still unknown. Microbes belonging to the genus, Dechloromonas, can reduce nitrate/nitrite in anoxic conditions using organic matter as energy and electron donors (Coates et al., 2001).
In the CDN reactor, the genus, Dokdonella, was the one of abundant genera. Kim et al. were the first to report the function of Dokdonella spp. in nitrogen and/or phosphate removal in the A2O process (Kim et al., 2013). Some clones detected in the CDN library were closely related to the genera, Thauera, Pseudomonas, and Paracoccus, which have been found to have roles in hydrogen-oxidizing autotrophic denitrification (Liessens et al., 1992; Vasiliadou et al., 2006; Mao et al., 2013). These bacteria are able to use nitrate/nitrite as an electron acceptor, hydrogen as an electron donor, and IC as a carbon source, in the absence of oxygen, to remove nitrogen from water with limited amounts of organic matter, such as groundwater and drinking water (Matějů et al., 1992). In addition, Thauera spp. have been reported to be heterotrophic denitrifying bacteria, using acetate as a carbon source (Mao et al., 2013). These results suggest that heterotrophic and hydrogen-based autotrophic denitrification are both occurring in the CDN reactor.
In the MDN reactor, Candidatus Accumulibacter phosphatis, Hydrogenophaga caeni, Sulfuritalea hydrogenivorans, and Azospira oryzae were detected. These species have been reported to have roles in phosphate accumulation, phosphorus removal, sulfur oxidation, and nitrogen removal, respectively (Blackall et al., 2002; Chung et al., 2007; Lin et al., 2014; Watanabe et al., 2014). One clone from the MDN reactor was affiliated with Methylotenera versatilis, a novel restricted facultative methylotroph isolated from lake sediment (Kalyuzhnaya et al., 2012).
Uncultured clones in the MDN library were almost always α-, β-, and γ-Proteobacteria, whereas most uncultured clones in the CDN library were identified as β-Proteobacteria. The diversity at the genus level shows that the MDN reactor permits bacteria with more diverse functions to coexist, ensuring a more stable system and improved denitrification performance. In the CDN library, OTUs CDN-06 (two clones) and CDN-08 (two clones) belonged to β-Proteobacteria and may be responsible for nitrogen removal. They shared the highest sequence similarity to clones Q7145-HYBO (JN391794) and B12 (AB661529), which were identified as denitrifying bacteria in a hybrid biofilm-activated sludge system (unpublished data) and during methane oxidation, respectively (Kojima et al., 2012). In the MDN library, OTU MDN-06 (one clone, β-Proteobacteria) showed 98% similarity with clone 114 (FM207944), a denitrifying bacterium that grows anaerobically with p-xylene as the sole organic carbon source and electron donor (Rotaru et al., 2010). The bacterium most similar to OTU MDN-12 (six clones, γ-Proteobacteria) was Thiothrix unzii (KF720709, only 96%), which is a filamentous sulfur bacteria able to covert nitrate into nitrite under anaerobic conditions (Trubitsyn et al., 2014).
Differences at the genus level of the phylum, Proteobacteria, were also observed in both nitrification reactors, but there was no common genus between the two reactors. In the CN reactor, the most abundant Proteobacterial clones were related to Leptothrix discophora, a producer of biogenic manganese oxides (Nelson et al., 1999). Leptothrix spp. have been identified as some of the filamentous bacteria observed in bulking sludge and some filamentous bacteria are known to have a role in nitrite oxidization (Martins et al., 2004). Their morphological features give them the advantage to preferentially access the substrate under substrate-limited concentrations. However, the presence of these filamentous bacteria at such a high ratio (40% of Proteobacterial clones) in the CN reactor may be a potential problem, in that it may induce sludge bulking. Clones affiliated with Hirschia sp. and Aeromonas media were the second most abundant bacteria in library CN. The latter were identified as biofouling bacteria forming a biofilm in the MBR (Lade et al., 2014). In the MN reactor, most Proteobacteria were identified as the genera, Woodsholea, and the denitrifiers, Dechloromonas, Paracoccus, and Azospira, with a few clones affiliated with the genera, Inhella, Lysobacter, and Reyranella. The most similar strains, Woodsholea maritime and Inhella inkyongensis, were novel species isolated from different environments and their functions are still unknown (Abraham et al., 2004; Song et al., 2009).
In addition, some of the uncultured clones, including OTU CN-10 and CN-13 from library CN and OTU MN-08 and MN-12 from library MN, may be involved in the nitrification process. These clones have the highest similarity to clone S7 (AF234717), which is present in nitrifying/denitrifying activated sludge (Juretschko et al., 2002); clone d_379_AG_Prop_353 (JN090816), from aerobic granules for phosphorus removal (Gonzalez-Gil and Holliger, 2011); clone F_SBR_39 (HQ010814), from sequencing batch reactors that use sludge fermentation liquid as a carbon source for nitrogen and phosphorus removal (Ji and Chen, 2010); and clone ASNR-29 (JQ809247), from the autotrophic nitrogen removal process in granular sludge bed reactors (Wang et al., 2012), respectively. However, a greater number of unclassified Proteobacteria were observed in the MN reactor than the CDN reactor, suggesting that these unknown species may be involved in the nitrification process in the MBR system.
Actinobacteria and Bacteroidetes were the second most abundant bacteria in CDN and MDN libraries, respectively. Most Actinobacteria (OTU CDN-19, nine clones) were identified as Candidatus Microthrix parvicella (DQ147285), a new species of filamentous bacteria isolated from industrial activated sludge wastewater treatment plants (Levantesi et al., 2006). This bacterium, together with Leptothrix sp., may contribute to the instability of the CAS system at a higher recirculation ratio. All Bacteroidetes clones (OTUs MDN-17, MDN-18, and MDN-19) in library MDN were closely related to the uncultured clones. OTU MDN-19 (five clones) had the highest similarity to clone LCFA-B03 (AB244310) for long-chain fatty acid degradation (Shigematsu et al., 2006b).
Clones classified as Bacteroidetes were the second most abundant clones in both CN and MN libraries and all of them were closely related to the uncultured clones. The Bacteroidetes species represented by these clones, including OTU CN-25 in CN library and OTUs MN-28, MN-29, and MN-30 in library MN, may be responsible for nitrogen removal. OTU CN-25 and OTU MN-29 have the highest similarity to clone Skagenf22 (DQ640688) and clone Skagenf55 (DQ640703), respectively, and both of them were identified in a full-scale enhanced biological phosphorus removal plant (Kong et al., 2007). OTU MN-28 and OTU MN-30 shared the highest similarity with clone A6 (AB462407), from an anaerobic upflow granular bed anammox reactor for nitrogen removal (Cho et al., 2010) and clone F_SBR_37 (HQ010812), from a sequencing batch reactor for nitrogen and phosphorus removal (Ji and Chen, 2010), respectively.
It has been widely accepted that Nitrosomonas and Nitrobacter are the most frequently detected bacteria in wastewater treatment plants, as the typical ammonium and nitrite oxidizers, respectively. However, no Nitrosomonas spp. were detected in the two systems, which suggests that other AOBs, such as unclassified clones with unknown function, were likely to be responsible for the nitrification. One study reported that operational parameters and wastewater type could greatly affect the AOB community (Park and Noguera, 2004). Similarly, clones of Nitrobacter were not identified in either CN or MN libraries. However, a few clones from another nitrite oxidizer, Nitrospira spp., were detected in the MN reactor. It is reasonable to conclude that they were more competitive in the MN reactor with a lower concentration of nitrite, because bacteria in the genus, Nitrospira, are K-strategist bacteria (Kim and Kim, 2006). Washout of slow-growing nitrifiers may explain the absence of Nitrospira. spp. in the CN reactor.
The remaining phyla in the CAS system were Planctomycetes, Firmicutes, and Cyanobacteria, along with Acidobacteria, Verrucomicrobia, Gracilibacteria, and Candidate division TM7, which showed low abundance in the MBR system. Representatives of the phylum, Planctomycetes, have previously been detected at a low percentage in both CAS and MBRs, where looser sludge was shown to allow the enrichment of Planctomycetes (Morgan-Sagastume et al., 2008; Chiellini et al., 2013). Members of these phyla have been poorly studied in MBRs and they are represented mostly by uncultivated organisms with unknown function. Therefore, their role in nitrogen removal requires further investigation.
Conclusion
In the current study, anoxic/oxic CAS and MBR systems, fed with low C/N ratio wastewater, were compared on performance and microbial communities involved in nitrogen removal. The MBR system was found to be more resistant to a high recirculation ratio than the CAS system. Different bacterial communities in the two systems, under a 10 L/day loading rate and a recirculation ratio of 1, were identified using 16S rRNA gene library analysis. Proteobacteria were the most abundant clones in all reactors, but there were clear differences in the abundance of clones and genera distribution in each Proteobacterial class. Higher bacterial diversity and a greater number of novel OTUs were found in the MBR system reactors than in the CAS system reactors. These differences in performance and bacterial communities may help when selecting a specific microorganism. This work increases our understanding of the relationship between bacterial structure and reactor performance for nitrogen removal.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.