
Abstract
Abstract
Selecting a desirable remediation technology is an emergency task for drought and infertile loess with serious dispersal of petroleum pollution in the region of Yanchang Oilfield, Yan'an, China. In this research, batch experiments were performed to investigate natural attenuation (NA), bioaugmentation (BA), and biostimulation combined with bioaugmentation (BSBA) effect on the degradation of petroleum hydrocarbons and indigenous microbial communities under low temperature (10°C). Results showed that compared to NA, (1) BA with JY (mixed strains cultured in laboratory) inoculation lead to symplastic growth of specific low temperature tolerant oil degraders Variovorax, Methanobrevibacter, and Comamonas and thus a higher degradation efficiency of aromatics (62.34%) and polar components (63.64%); and (2) BSBA showed negligible community shift since the indigenous microbe competing ability was enhanced by NH4Cl addition, which contributes to an adding effect of JY and indigenous petroleum degraders for best performance in total petroleum hydrocarbon (34.76%), saturate (27.62%), and aromatic (70.13%) removal. This study demonstrated that BSBA had the best removal efficiency and BA was more suitable for purpose of some special fraction removal.
Introduction
Total Petroleum Hydrocarbons (TPHs), mainly composed of carbon and hydrogen, are a group of organic contaminants derived from a mixture of fuels and crude oil byproducts released into the environment through mining, manufacturing procedures, and accidental spills (Aiyoub et al., 2016). Contamination and accumulation of TPHs in ecosystem can cause serious carcinogenesis, teratogenesis, and gene mutations (Chen et al., 2017; Wu et al., 2017).
Yanchang Oilfield is located in the gully region of the Loess Plateau in Yan'an city, China. In the past 20 years, petroleum production from this field has increased from 1 million tons to more than 12 million tons per year. Unfortunately, environmental pollution has accompanied with petroleum exploitation. Since drought and infertility are the key characters of the local soil, large area soil erosion has caused serious dispersal of petroleum pollution in soil, river, and river sediments in this area (Wang et al., 2018). Therefore, selecting a desirable remediation technology is an emergency task for this region.
According to previous reports, bioremediation, based on the metabolic capabilities of microorganisms, was a cost-effective and environment-friendly technology for the treatment of petroleum contaminants in soils and sediments (Chang et al., 2010; Aiyoub et al., 2016; Wu et al., 2016). However, Yanchang Oilfield is located in cold area with an average annual temperature of 10.4°C, and low temperature was supposed as an adverse factor on petroleum hydrocarbon biodegradation, since the low temperature inhibited the production of some oil degrading related enzymes (Walworth et al., 2001; Delille et al., 2007; Ferguson et al., 2008). Even so, several studies found that cold adapted microorganisms could change metabolic mechanism and pathway when temperature decreased and kept the potential for bioremediation of petroleum hydrocarbons in soil under cold environment (Chang et al., 2010; Jeong et al., 2015; Wang et al., 2015).
Nowadays, many cold-adapted bacteria, such as the Gram-negative bacteria Pseudomonas, Flavobacterium, Alcaligenes, Aeromonas, Xanthomonas, and Serratia and the Gram-positive bacteria Arthrobacterium, Bacillus, and Micrococcus, have been confirmed to be able to degrade petroleum at low temperature (Margesin and Schinner., 2001; Margesin and Miteva., 2011). Based on cold adapted microorganisms, various bioremediation technologies have been employed to effectively degrade petroleum in contaminated soil under cold weather conditions (Aislabie et al., 2006; Chang et al., 2011; Kauppi et al., 2011; Sanscartier et al., 2011).
Research on bioremediation confirmed the feasibility of different strategies, such as biostimulation (BS) (with nutrient amendment) and bioaugmentation (BA) (with inoculation of petroleum-degrading microorganisms), to degrade petroleum hydrocarbons under cold weather conditions (Francisco and Majid, 2013). However, these bioremediation techniques will disturb the indigenous microbial community (Wu et al., 2016), in which diversity and activity are important in ensuring the effective bioremediation of petroleum-contaminated soil in local (Kauppi et al., 2011; Liu et al., 2012a; Taccari et al., 2012). However, little research has been conducted to investigate the effect of these bioremediation methods on indigenous microbial community, which contribute to the natural attenuation (NA) of petroleum contaminants (Taccari et al., 2012; Aiyoub et al., 2016; Wu et al., 2016).
To provide a feasible bioremediation method to cope with the stressing petroleum contamination in the cold region, batch experiments were performed to investigate NA, BA (supplement with JY, a mixed petroleum-degrading culture), and biostimulation combined with bioaugmentation (BSBA) (supplement with nitrogen and JY) effect on the degradation of petroleum hydrocarbons and indigenous microbial communities under low temperature (10°C). This study aimed at providing a theoretical support and suitable technical strategy for regional petroleum hydrocarbon pollution restoration.
Materials and Methods
Crude oil, soil, and petroleum-degrading bacterial flora
Crude oil
Crude oil was taken from Yanchang oil field in the city of Yan'an, located in loess plateau in northern-west of China. The crude oil was firstly dissolved in petroleum ether with boiling range of 60–90°C. Then the solvent was filtrated with Na2SO4 and the petroleum ether was volatilized by a 60°C water bath to obtain the pure oil. Finally, 10 g of the pure oil was dissolved in 1,000 mL petroleum ether with boiling range of 30–60°C to get 10 mg/mL standard stock oil solvent and was stored at 4°C for further utilization.
Soil
The soil used in this study was collected from loess plateau; the sampling site was far from the oil field. Before experiments, impurities such as stones, grass roots, and worms were removed by hands, the soil was dried by natural weathering for 2 days and ground, and screened by 2.0 mm sieve to obtain the uncontaminated soil (UCS).
Added a certain amount of standard stock oil solvent into UCS and mixed homogeneously. Placing the mixture in ventilation hood in order to volatilized the petroleum ether. Then the petroleum contaminated soil with the concentration of 30g/kg was obtained. The physiochemical properties of the UCS and contaminated soil are measured and listed in Table 1.
Physiochemical Properties of Uncontaminated Soil and Initial
TOC, total organic carbon; UCS, uncontaminated soil.
Petroleum-degrading bacteria flora
A group of petroleum-degrading bacteria isolated from Alpine Meadow soil was used as inoculating source in this study. The bacteria flora was named as JY, which can utilize petroleum as the sole carbon source at low temperature (10°C). The flora includes Arthrobacter sp. XD-B1, Arthrobacter sp. XD-R1, Arthrobacter sp. XD-H3, Pseudomonas sp. XD-S, Rhodococcus sp. XD-R2, Stenotrophomonas sp. XD-H1, and Sphingobacterium sp. XD-H2.
Bioremediation assays
The bioremediation was investigated in BA and BSBA assays. NA was conducted as control. All the bioassays were conducted in plastic boxes (3.5 L) with 2 kg soil and incubated at 10°C for 91 days. BA assays were conducted with 2 kg contaminated soil (30 g/kg) and by the addition of 108 cell/g bacteria flora JY. BSBA assays were conducted with 2 kg contaminated soil (30 g/kg) and supplement of 108 cell/g bacteria flora JY and 1.6 g NH4Cl (to maintain the C: N: p = 100: 10: 1). NA assays were conducted with only 2 kg contaminated soil (30 g/kg). Each treatment was set up in triplicate. The soil was stirred periodically to ensure oxygen supply, and distilled water was added regularly to keep the soil moisture around 15%. Another two controls, UCS and Initial, were also conducted. UCS was the soil collected from the study area with no oil pollution and Initial was the sample with oil addition at the study time of t = 0.
Analysis methods
Physiochemical properties of soil
The pH values of the samples were analyzed using a pH meter (Ohaus ST2100/3C Pro-B; America) in a soil and deionized water suspension (1:2.5 soil weight/water volume). Total organic carbon (TOC) was measured using a Total Organic Carbon Analyzer (ET1020A, Shanghai Continental Technology Co. Ltd. China). Total nitrogen (TN) was analyzed using the Kjeldahl method (Wang et al., 2018). Total phosphorous (TP) was analyzed using an UV-vis spectrophotometer after wet digestion with HClO4-H2SO4 (Wang et al., 2018). Moisture was measured using the oven drying method (Koponen et al., 2006).
TPH, saturates, aromatics, and polar components
After 91 days of incubation, 5 g of soil samples from each pot were extracted to determine the TPH contents by the gravimetric method (Wang et al., 2018). The saturates and aromatic fractions were separated by column chromatography using a silica gel alumina column (30 cm × 10 mm i.d). Briefly, extracted TPH was dissolved in n-hexane and loaded on the column. A successive elution was performed using 30 mL of n-hexane and 40 mL of n-hexane/DCM (1:2, vol: vol) to obtain the saturates and aromatic fractions, respectively. These two fractions were then determined gravimetrically after the solvent evaporation. The polar hydrocarbon fractions were the amount of TPH minus that of saturates and aromatics. Afterward, all extracted samples were stored at 4°C for next analysis. Each treatment was conducted in triplicates.
GC-MS for n-alkanes and 16 USEPA PAH analysis
The separated elution of saturates and aromatics was each concentrated to 0.5 mL under a gentle N2 stream. An aliquot of 1,500 ng of hexamethyl benzene (AccuStandard, Inc.) was added as an internal standard before analysis. The hexamethyl benzene was dissolved in n-hexane to the concentration of 1,000 mg/L, and then an aliquot of 1.5 μL of this solution was injected to the elution using a 5 μL microliter syringe (Hamilton, Switzerland) as the internal standard.
N-alkenes were analyzed using gas chromatography-mass spectroscopy (GC-MS) (Agilent 6890 GC with an Agilent 5973 mass spectrometer equipped with a 30 m × 0.25 mm HP-5 fused silica capillary column).
The GC oven temperature was programmed as follows; first, the ion source was maintained at 230°C; the inlet temperature and the transfer line temperature were maintained at 280°C. The temperature program was held for 1 min at 70°C; the temperature was then increased to 300°C at a rate of 4°C per min. This was followed by a temperature hold for 5 min at 300°C. The injection volume was 1 μL and the carrier gas was helium (1 mL/min). The mass spectrometer was operated at electron ionization energy of 70 eV. Aliphatic hydrocarbons were determined at a level of 57 m/z. Peak levels of individual n-alkanes were used to determine their respective concentrations, using external hydrocarbon mixture standards ranging from n-C8 to n-C40 (AccuStandard, Inc.).
The 16 Polycyclic Aromatic Hydrocarbons (PAHs) were analyzed using the same GC-MS operating in the selected ion-monitoring mode. The GC oven temperature for PAH analysis was programmed as follows; the temperature was held at 60°C for 2 min. This was followed by an increase in the temperature to 290°C at a rate of 3°C per min. The temperature was then held at 290°C for 5 min. The standard PAH curve was established using 16 PAH mixture standards at a concentration of 200 ppm (Accu Standard, Inc.).
DNA extraction, PCR, and Illumina MiSeq sequencing
Total genomic DNA was extracted directly from soil samples using E.Z.N.A.® Soil DNA Kit (Omega Biotech, Norcross, GA) following the manufacturer's protocol. The DNA concentration was then determined using a Qubit 4.0 Fluorometer Pub.No. MAN0017210 (Invitrogen, Carlsbad, CA). Agarose gel electrophoresis was conducted to examine the completeness of extracted DNA. The DNAs extracted from three duplicated samples were pooled together.
The V3–V4 region of 16S rRNA gene was amplified using the primer sets in MiSeq platform,
341F(CCCTACACGACGCTCTTCCGATCTGCCTACGGGNGGCWGCAG) and 805R(GACTGGAGTTCCTTGGCACCCGAGAATTCCAGACTACHVGGGTATCTAATCC). PCRs were carried out according to the methods described by Sutton et al. (2016). The PCR products were sequenced on Illumina MiSeq platform and the data processing was performed by Sangon Biotech Co., Ltd. (China). Then FLASH v1.2.7 and PRINSEQ-lite 0.19.5 were used to control the sequence quality. The operational taxonomic units (OTUs) were classified using uclust v1.1.579 (www.drive5.com/uclust/downloads1_1_579.html) at 97% sequence similarity after correcting the wrong sequence using precluster and using uchime to cut the chimera. Species diversity indices were calculated through the Mothur program (www.mothur.org). The statistical analyses were performed in the R environment (version 3.2.1; www.r-project.org). The heat map was generated by R ggplot software package.
Statistical analysis
Tukey's test for multiple comparisons was performed using SPSS software.
Results and Discussion
Degradation of petroleum hydrocarbons
Degradation of TPH and its different fractions
Degradation of TPH and its different fractions showed discrepancies in different bioremediation treatments (Fig. 1). After 91 days of remediation, the residual concentrations of TPH were detected at 25,700, 28,300, and 24,400 mg/kg in NA, BA, and BSBA, respectively. In comparison with initial TPH concentration of 37,400 mg/kg, removal percentages of 31.28%, 24.33%, and 34.76% were achieved in NA, BA, and BSBA, respectively (Supplementary Table S1). These three treatments showed significant effect in TPH removal (p < 0.05). Furthermore, BSBA contributed to a slight enhancement in TPH removal compared with NA (p > 0.05), while BA has an inhibition effect significantly (p < 0.05).
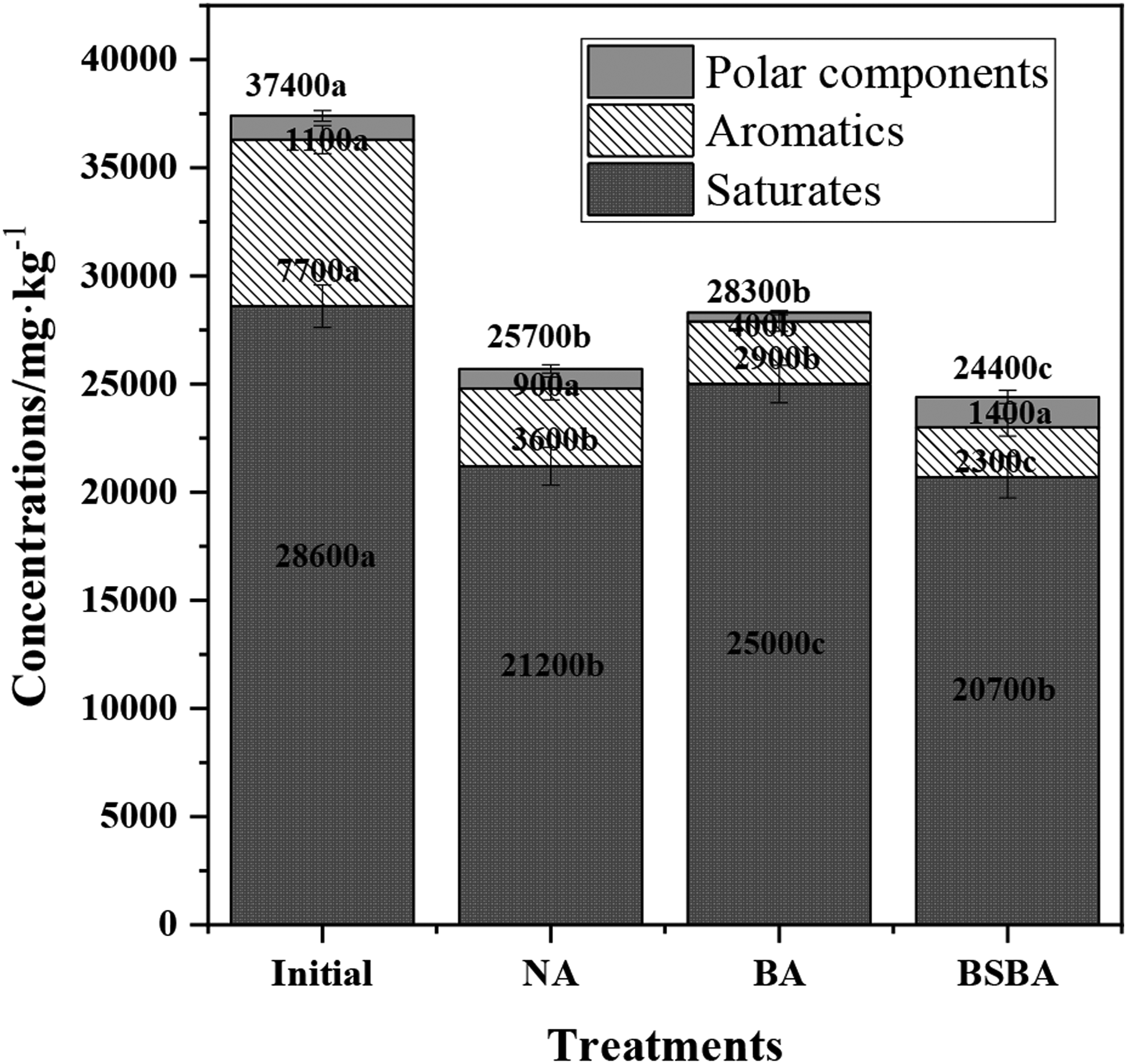
Concentrations of TPH and its different fractions (saturates, aromatics, and polar components) before and after 91 days of biodegradation. Bars represent the average concentration of three replicate incubations and error bars represent the standard deviation. Different letters in the same petroleum hydrocarbon fractions represent a significant difference at p ≤ 0.05. TPH, total petroleum hydrocarbons.
According to the initial composition of TPH's fraction, saturates (28,600 mg/kg) were apparently the main component, accounting for 76.47% of the TPH (37,400 mg/kg), and followed by aromatics (7,700 mg/kg, 20.59%) and polar components (1,100 mg/kg, 2.94%). After 91 days of remediation, the residual concentrations of saturates were 21,200, 25,000, and 20,700 mg/kg with the removal percentages of 12.59%, 5.87%, and 27.62% in NA, BA, and BSBA, respectively. BSBA showed a slight enhancement (p > 0.05), while BA showed an inhibition (p < 0.05) compared to NA.
In terms of aromatics, the removal percentages were 53.25%, 62.34%, and 70.13%, corresponding to the residual concentrations of 3,600, 2,900, and 2,300 mg/kg in NA, BA, and BSBA, respectively (Supplementary Table S1). Obviously, BA showed a slight enhancement (p > 0.05), but BSBA has a significant acceleration (p < 0.05) on aromatic removal compared to NA. However, for the polar components, 900, 400, and 1,400 mg/kg were detected in the residual soil of NA, BA, and BSBA treatments, which showed a significant positive effect in BA (p < 0.05), a slight enhancement in NA (p > 0.05), but an opposite removal efficiency in BSBA (p < 0.05).
Compared to NA, BA showed a less performance in TPH removal, while for the removal of aromatics and polar components, it showed quite a good performance. The intermediate metabolites produced in BA may contribute to the relative high concentration of saturates in this assay. Al-Daher et al. (1998) reported a TPH removal percentage of 60% by nature attenuation, but the BA had no effect on TPH removal. Also, Mancera-Lopez et al. (2007) proved that BA could improve the bioremediation efficiency, especially for special matters that could be metabolized by the added bacterial consortium, which might be the reason of our observation, as JY addition contributes to large amount of aromatics' removal.
Besides, BSBA showed as the best strategy on saturate and aromatic removal, but an accumulation of polar components. Some studies have found that saturated and aromatic hydrocarbons were relatively easy to be degraded. In contrast, the character of stability made polar compounds recalcitrant toward degradation (Atlas and Hazen, 2011). Our results were consistent with these references, as higher saturated and aromatic hydrocarbon removal were observed in all treatments, but polar components persisted in soil samples, and even accumulated in BSBA group.
The accumulation of polar components was described by many other researchers as a result of partial hydrocarbon degradation (Dibble and Bartha, 1979; Chaineau et al., 2005; Lee et al., 2007; Yanto and Tachibana, 2014). Fedorak and Westlake (1981) noted that increase of polar compounds in soil may be caused by the generation of polar biological metabolites during biodegradation. Thus, both recalcitrant polar components in petroleum and polar biological metabolites from petroleum biodegradation may contribute to the accumulation of these compounds.
A better TPH degradation efficiency for BSBA compared to BA treatment might result from that, proper C: N: P ratio in soil stimulated the activity of indigenous petroleum hydrocarbon degraders in soil. Mair et al. (2013) proved that the addition of nutrients had a significantly stimulating effect on the biodegradation activity of indigenous microorganisms. Atlas and Hazen (2011) also proved that the addition of nitrogen nutrient could stimulate the growth and degrading activity of the indigenous hydrocarbon microorganisms. Suja et al. (2014) set up laboratory and field experiments to study the effects of local microbial BA and biostimulation on the bioremediation of TPH in crude oil-contaminated soil and concluded that BSBA was the best method and degraded TPH markedly.
The effects of petroleum hydrocarbon pollution in soil on the ecological environment were mainly contributed by saturates and aromatics. Importantly, many aromatics show carcinogenic, teratogenic, and mutagenic effects on people and animals. Aromatics are the most toxic components in TPH; thus, the main goal for remediation of petroleum hydrocarbon-contaminated soil is to remove aromatic hydrocarbons. Although aromatic content distribution in TPH is less than saturates and polar components, their environmental impact is much higher than other components.
This study showed that BA was an effective bioremediation method for the key toxic aromatics. BSBA showed a good strategy on both saturate and aromatic removal, but led to the accumulation of polar components. However, these polar residues are usually less bioavailable and inert to impact the ecology. From the point of environmental effects about different fractions of TPH, BSBA is more valuable than other treatments, while BA is also a good choice for relieving of the toxic aromatics.
Concentration of n-alkanes and 16 priority-controlled PAHs
Biodegradation of n-alkanes was varied among different remediation treatments (Fig. 2).

Concentrations of n-alkanes (C14–C36) in different treatments before and after 91 days of biodegradation. Bars represent the average concentration of three replicate incubations and error bars represent the standard deviation.
In general, after 91 days of bioremediation, obvious degradation of most n-alkanes was observed in all treatments. Comparatively, BSBA treatment had the highest removal percentages in most quantified n-alkanes here except C17, C30, C33, C34, and C36. The highest removal percentage (52.61%) of BSBA occurred in C35 (Supplementary Table S2). As to BA treatment, it performed better than BSBA in C17, C30, C33, and C36 degradation in this study. In terms of the degradation of n-alkanes, it indicated that low carbon number alkanes were easier to be degraded, which might be because long-chain lengths of n-alkanes were solid and had low solubility for degradation by microorganisms (Fan et al., 2014). Yan et al. (2015) also claimed that for the high carbon number alkanes, the relatively low biodegradability could be attributed to low water solubility.
Biodegradation of 16 priority-controlled PAHs was evaluated after different bioremediation (Fig. 3). For most PAHs, both BA and BSBA showed better degradation effect than NA. Furthermore, BA showed the best degradation effect on most PAHs, including phenanthrene, fluoranthene, pyrene, chrysene, benzo[k]fluoranthene, benzo[b]fluoranthene, benzo[a]pyrene, diphenyl[a,h]anthracene, indeno[1,2,3-cd]Pyrene, and benzo[g,h,i]perylene. It indicated that JY addition could promote the degradation of high ring number PAHs effectively.
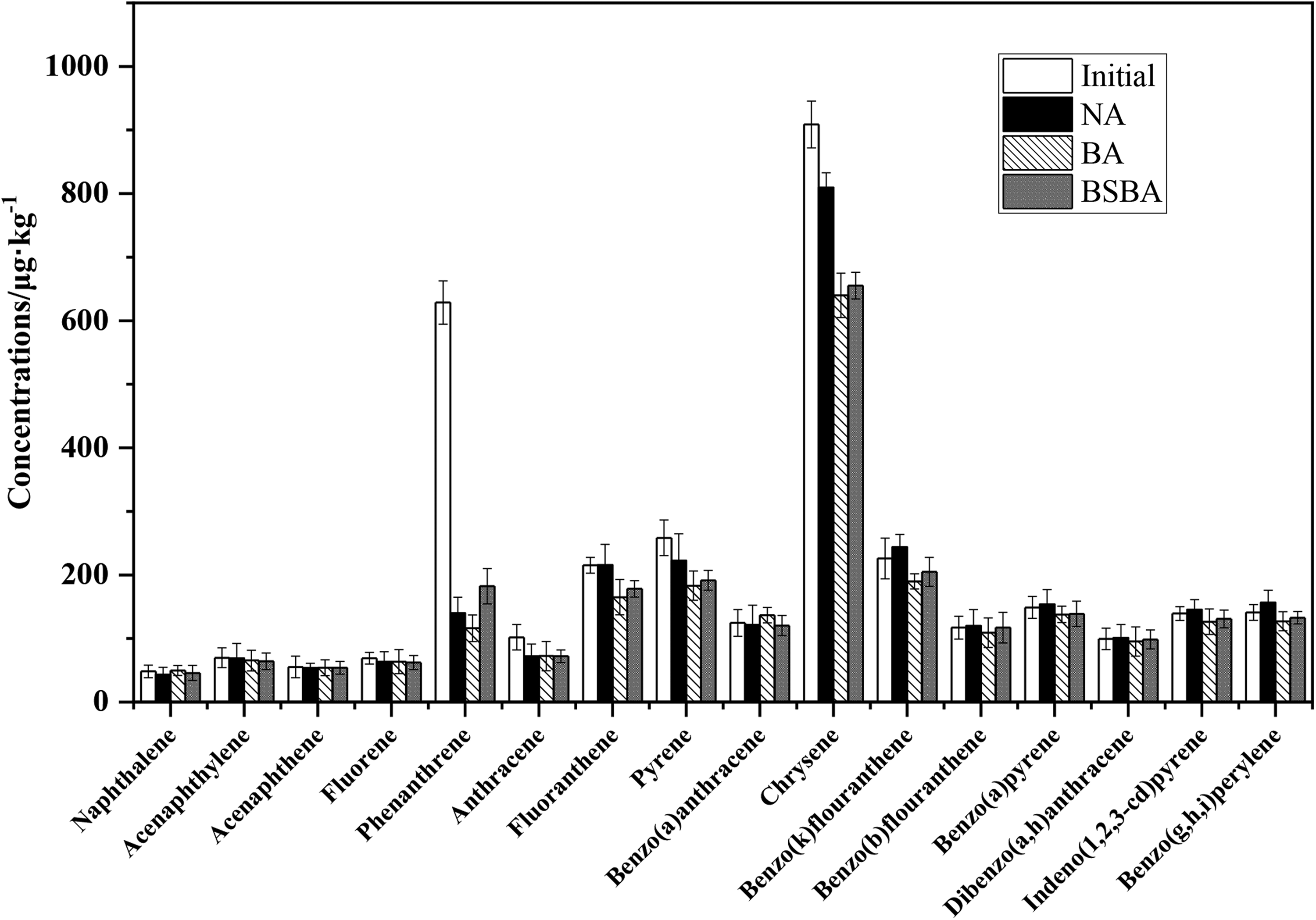
Concentrations of 16 USEPA PAHs in different treatments before and after 91 days of biodegradation. Bars represent average concentration of three replicate incubations and error bars represent standard deviation.
However, supplementation of NH4Cl could not apparently help JY and indigenous microbes in improving PAH removal. Among 16 priority-controlled PAHs, both phenanthrene and chrysene were much richer than other PAHs in the initial contamination soil, while the removal percentage of phenanthrene (77.73% in NA, 81.52% in BA, and 71.00% in BSBA) was much higher than chrysene (10.93% in NA, 29.60% in BA, and 27.90% in BSBA), which indicated low-ring PAHs were relatively easier to be degraded than high-ring PAHs (Supplementary Table S3). It was obvious that the degradation of phenanthrene was significant no matter in whichever treatments, which might be related to the special actively degradable molecular structure of phenanthrene.
What is more, the position of substituents in benzene ring affected the biodegradability of aromatics, which could be informed from the different biodegradation efficiency of benzo[k]fluoranthene and benzo[b]fluorathene at the same treatment strategy (Fan et al., 2014). Fan et al. (2014) also indicated that with the increase of benzene ring numbers and the complexity of molecular structures, the degradation of aromatics became more and more difficult, which was not reflected obviously in this study. Also, Kong et al. (2018) demonstrated that elimination of PAHs was significantly enhanced (increased by 54.45%) in the bioaugmented mesocosms and PAHs with four to six rings were more difficult to be degraded than lower rings PAHs.
Microbial structure and biodiversity
Microbial diversity indices
The Illumina sequencing found the highest reads of 39,538 in UCS, followed by 44,563 in BA, 10,442 in NA, and 9,197 in BSBA (Table 2). The number of OTUs in UCS was the highest of 9,417, but decreased sharply to 1,800 in NA, 4,253 in BA, and 1,728 in BSBA. Consistently, Shannon, ACE, and Chao1 indices showed the same variation trend (except BA) in the petroleum-contaminated soil. It suggests that oil contamination significantly inhibited the growth of partial soil organisms, which lead to a lower microbial richness and diversity in the contaminated soil samples. However, BA treatment has the lowest Shannon even and the highest Simpson indices, which means some single microbes have enriched to a large number.
Operational Taxonomic Unit Number and α-Diversity Indices in Five Samples
NA, natural attenuation; BA, bioaugmentation; BSBA, bioaugmentation combined with biostimulation; OUT, operational taxonomic unit.
The diverse indices are dependent both on richness and evenness of a community. Previous studies have found petroleum contaminants may result in either an increase (Peng et al., 2015) or a decrease (Sutton et al., 2013; Jiao et al., 2016) in microbial diversity. The biodiversity decrease in this study might be caused by: (i) the inhibitory effects of toxic petroleum and biodegradation metabolites (Jiao et al., 2016), and (ii) lack of electron acceptor for microbial community due to TPH mineralization (Zhou and Crawford, 1995).
Besides, we found that BA can change the microbial community greatly because of the higher richness, but lower evenness in BA. This phenomenon might be caused by JY inoculation. The added JY consortium compete the necessary nutrients for growth with the indigenous microbes. Expectedly, in BSBA, it was similar to NA extremely both on richness and evenness of the microbial community. Apparently, the NH4Cl addition alleviated the community shift resulted by JY inoculation. Many studies have proven that biostimulation can promote the growth of indigenous microbes through the addition of nutrients in soil (Taccari et al., 2012; Polyak et al., 2018)
Distribution of taxons at phylum level
In this study, twenty-eight bacterial and two archaeal phyla (Supplementary Table S4) were identified. The relative abundance of nine dominant phyla, including eight bacteria and one archaea, is shown in Fig. 4. Proteobacteria was the most dominant phylum in all treatments, with relative abundance of 46.35% in UCS, 41.76% in NA, 78.96% in BA, and 43.19% in BSBA. The relative abundance of some other phyla such as Firmicutes, Planctomycetes, and Gemmatimonadetes accounted for 6.33%, 2.35%, and 3.96% in UCS, but decreased to 3.32%, 0.49%, and 0.06% in NA.
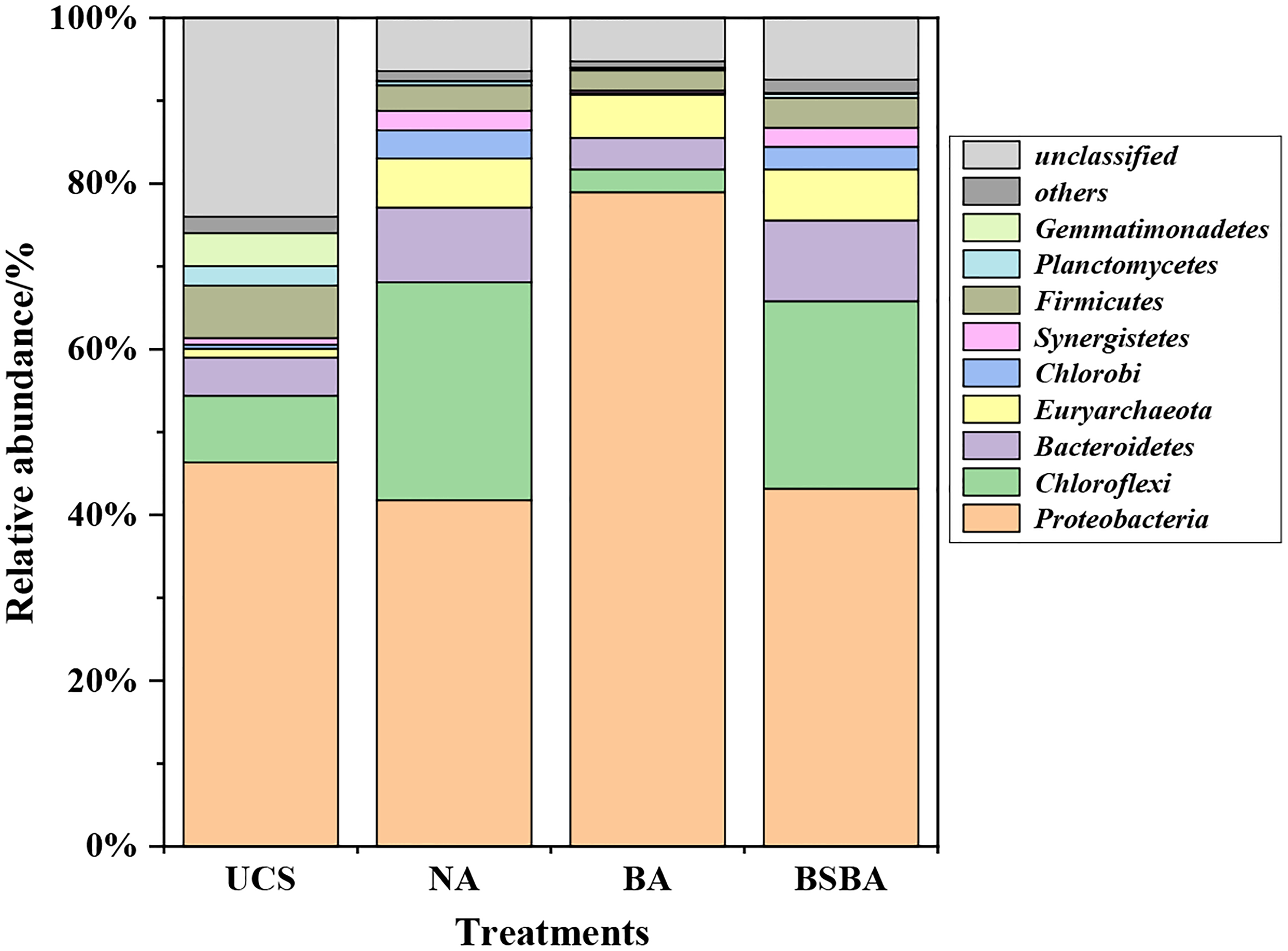
Relative abundance of nine most abundant phyla in different treatments. Relative abundance was displayed in terms of percentage in total effective sequences
On the contrary, a number of phylotypes in other phylum have enriched after oil addition. For instance, Chloroflexi, Bacteroidetes, Euryarchaeota, Chlorobi, and Synergistetes accounted for 8.03%, 4.62%, 1.07%, 0.51%, and 0.80% in UCS and were increased to 26.33%, 9.02%, 5.93%, 3.39%, and 2.36% in NA. Such variations of different microbial phylum indicated petroleum contamination has a selective effect on indigenous flora for stimulating the oil-degrading or tolerant microbes to grow (Sauret et al., 2014).
Furthermore, compared to NA, BA influenced the microbial phylum composition significantly. For BA treatment, a significantly different and inhomogeneous microbial community structure appeared with a large number of Proteobacteria (78.96%) and less than 22% for the total relative abundance of others, including Euryarchaeota (5.25%), Firmicutes (2.43%), Bacteroidetes (3.77%), and Chloroflexi (2.75%). Clearly, the bacterial communities were significantly affected by JY inoculation as shown in BA treatment, whereas nutrients addition balanced the indigenous bacterial community, which was confirmed by a similar microbial constitute observed in BSBA and NA.
All the phyla dominated in this study have been found in many other different oil pollution researches (Koo et al., 2015; Peng et al., 2015; Jiao et al., 2016). Proteobacteria and Firmicutes were widely found in low temperature environment (Perreault et al., 2007; Peeters et al., 2011). Proteobacteria has been proven to not only exhibit strong tolerance to the low temperature but also showed potential organic degradation capacity at low temperature (Xie et al., 2017). In this study, Proteobacteria was also the most dominant phylum in all soil samples at a low temperature of 10°C, and furthermore, JY inoculation enhanced Proteobacteria remarkably.
Distribution of taxons at genus level
The hierarchically clustered heatmap plot showed that NA and BSBA libraries grouped first together, and then they clustered with BA treatment by decreasing the order of similarity (Fig. 5). UCS has the longest clustering distance with other three oil pollution treatments. These results indicated that the microbial community structure was similar in oil-contaminated samples, but significantly different from the uncontaminated one.
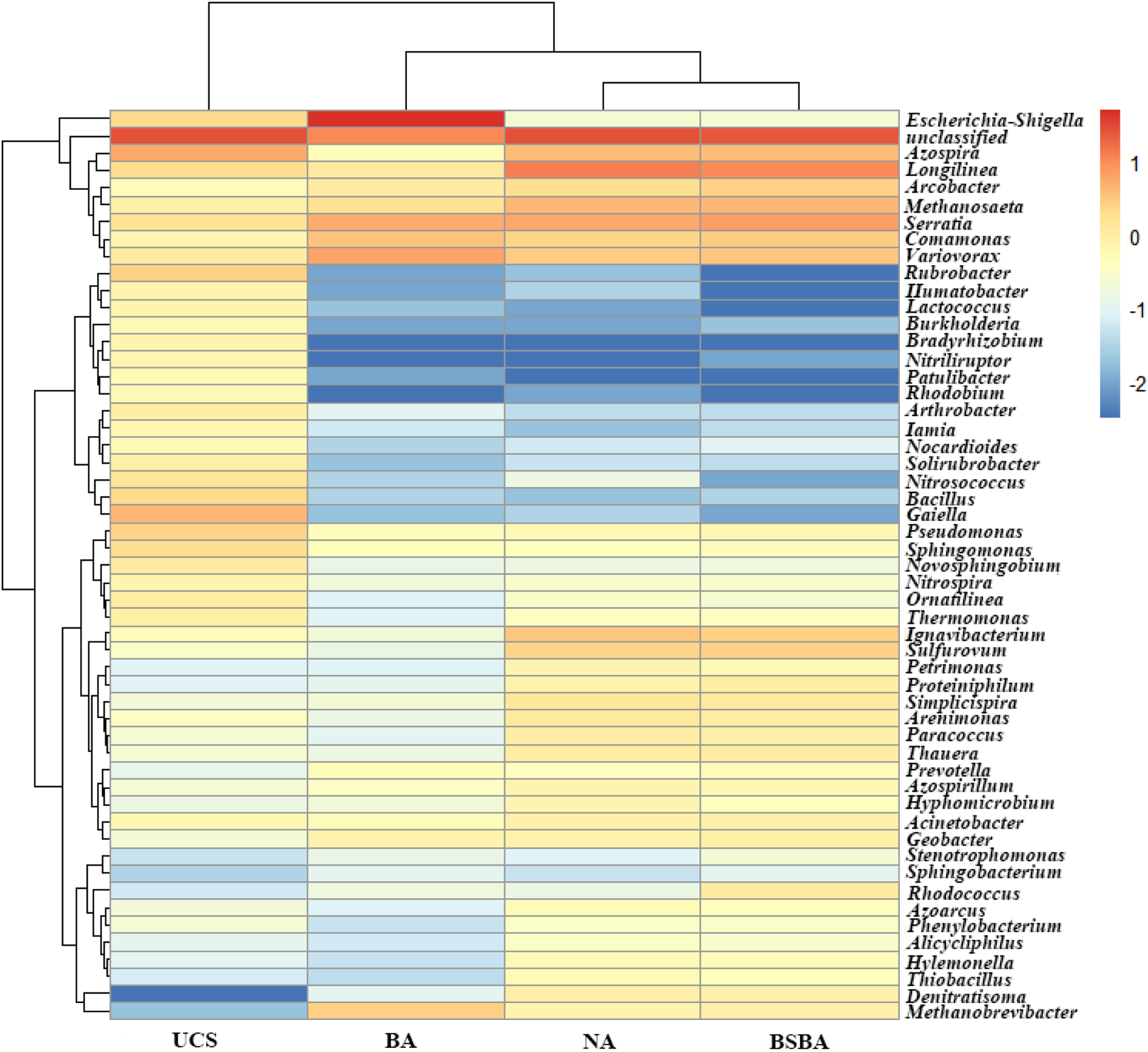
Hierarchically clustered heat map of the distribution of taxons at the genus level with the relative percentage higher than 0.5% in the five samples. Row represents the relative percentage of each genus, and column stands for different samples. The relative abundance for each genus was depicted by color intensity with the legend indicated in the right of the figure.
Microbial distribution in NA
Among all these identified genus, some phylotypes have enriched greatly after petroleum contamination. For instance, Longilinea, Serratia, Methanosaeta, Ignavibacterium, and Variovorax were the dominant genus in NA with the relative abundances of 13.31%, 5.63%, 4.56%, 3.37%, and 3.04%, but accounted for 2.11%, 1.63%, 0.94%, and 1.16% in uncontaminated sample, respectively Supplementary Table S5. Notably, other six microbes of Comamonas, Arcobacter, Methanobrevibacter, Sulfurovum, Arenimonas, and Thauera have the relative abundances lower than 0.68% in UCS, but increased to a range of 0.72%–2.51% in NA treatment. Many of these microbes have been proven to be oil-tolerant or oil-degrading microorganisms.
The predominance of strictly anaerobic bacteria Longilinea within Chloroflexi, Ignavibacterium within Chlorobi, and archaea Methanosaeta within Euryarchaeota indicated that NA has occurred under anoxic conditions, while the predominance of facultatively anaerobic bacteria Serratia within Gammaproteobacteria and aerobic bacteria Variovorax within Betaproteobacteria suggested that aerobic petroleum degradation has also happened. All these microbes have been found to participate in oil degradation process in previous studies.
Methanosaeta has previously been observed in oil sands (Penner and Foght, 2010) and has been observed in consortiums able to perform anaerobic mineralization of long alkane chains (Zengler et al., 1999; Lanoil et al., 2001). Longilinea affiliated to class Anaerolineaceae has been found to syntrophically cooperate with Methanosaeta archaea involved in the process of methanogenic degradation of n-alkanes in the last period during long-term degradation (Liang et al., 2015). Ignavibacterium album was found to have terminal oxidases, which were widely reported in alkane biodegradation (Liu et al., 2012b). Reports about Serratia function in environment was hardly found, but the species Serratia marcescens has been reported to degrade aromatic aldehydes, which were the metabolites of aromatic hydrocarbon biodegradation (Delafuente et al., 1991).
Variovorax, a versatile genus containing various alkane- and aromatic hydrocarbon-degrading species under aerobic conditions, has been found by many studies. Jones et al. (2011) proved that species of Variovorax can degrade naphthalene, anthracene, and Benzo[a]pyrene in soil by stable isotope probing DNA technology, Martirani-Von Abercron et al. (2017) found Variovorax enriched to 54% when the oxygen was high in a PAH bioremediation treatment, Posman et al. (2017) found Variovorax MAK3 can express aromatic ring-hydroxylating dioxygenase to be involved in benzene degradation, and Han et al. (2011) found that species of Variovorax paradoxus S110 has diverse metabolic capabilities of biodegradation of both biogenic compounds and anthropogenic contaminants, and it also engages in mutually beneficial interactions with both bacteria and plants.
Moreover, there were many petroleum-degrading microbes among other microorganism's flora enriched in NA. Comamonas sp. has been reported to degrade phenanthrene and anthracene in soil (Widada et al., 2002), and Comamonas testosterone GZ42 metabolizes phenanthrene in soil and sediment (Cebron et al., 2008; Edlund and Jansson, 2008). The genus Arenimonas belongs to Gammaproteobacteria, which is a well-known oil-degrading bacterium; an example includes Arenimonas malthae sp. nov isolated from an oil-contaminated site in Taiwan (Young et al., 2007). In addition, Arenimonas was found to be enriched highly in contaminated soil in oil refineries and was positively associated with TN (Jiao et al., 2016).
Arcobacter is a genus of Gram-negative, spiral-shaped bacteria in the Epsilonproteobacteria class. Species of the genus Arcobacter currently consists of five species: A. butzleri, A. cryaerophilus, A. skirrowii, A. nitrofigilis, and A. sulfidicus, although several other potential novel species have recently been described from varying environments (Fera et al., 2004; Donachie et al., 2005). Three of these five known species are pathogenic, while other two are nonpathogenic strains. A. nitrofigilis is a nitrogen-fixing bacterium isolated from the roots of the salt marsh plant Spartina alterniflora (Fera et al., 2004). A. sulfidicus is an obligate microaerophile that oxidizes sulfides and is an autotrophic producer of filamentous sulfur (Fera et al., 2004).
Genus of Thauera within Betaproteobacteria has been found with a major characteristic of recognized species, so far with the ability of utilization of aromatic or terpenoid hydrocarbons under denitrifying conditions (Bradley et al., 2009). These microbes with various metabolite abilities of nitrogen, sulfur, and hydrocarbon cooperated together to degrade petroleum and while forming the microbial community structure in the soil.
Microbial community shift in BA and BSBA
Furthermore, BA has influenced the distribution of some special microbes. For instance, Variovorax (6.69%), Methanobrevibacter (2.79%), and Comamonas (3.61%) enriched in the BA treatment compared to NA. However, in BSBA, these microbes have the similar relative abundances with NA. This result demonstrated that JY inoculation promoted the growth of these microbes.
The JY bacterial consortia contain three strains of Arthrobacter and one strain of Sphingobacterium, Stenotrophomonas, Rhodococcus, and Pseudomonas, respectively. These strains were all well-known TPH degraders, and have good performance, especially in PAH metabolism under different conditions.
Rhodococcus has been reported with the ability of degrading both alkanes and aromatics in the temperature range between 4°C and 25°C and Pseudomonas was proven to degrade PAHs only at low temperatures of 4°C and 10°C (Hamamura et al., 2008). Stenotrophomonas maltophilia has been found can degrade crude oil, Benz[a]anthracene, Benzo[a]pyrene, Chrysene, Coronene, Dibenz[a,h]anthracene, Fluoranthene, and Phenanthrene (Obuekwe et al., 2009; Varjani et al., 2015; Varjani., 2017a). Arthrobacter gandavensis has been found to metabolize pyrene and Arthrobacter sp. P1-1 can degrade phenanthrene and naphthalene (Seo et al., 2006). Sphingobacterium spiritivorum AS43 has been reported to produce biosurfactants to enhance the biodegradation of used lubricating oil (Noparat et al., 2014).
The possible reason for enrichment of Variovorax (6.69%), Methanobrevibacter (2.79%), and Comamonas (3.61%) in BA were as follows: (i) the intermediate metabolite of JY bacterial consortia has provided the growth substrates for these microbes; (ii) the JY consortia have produced some substances that enhanced the bioavailability of petroleum in soil, especially for the high molecular aromatics and polar components. As mentioned above, a bacteria strain Sphingobacterium in JY can produce biosurfactants that can enhance the solubility of hydrocarbon fractions to enhance the biodegradation of used lubricating oil (Lawniczak et al., 2013; Noparat et al., 2014). Moreover, the high degradation efficiency of PAHs and polar components in BA may be correlated with the high enrichment of Variovorax and Comamonas.
However, there were some microbes, such as Azospira (0.47%), Arcobacter (1.17%), Arenimonas (0.14%), Ignavibacterium (0.2%), Longilinea (1.23%), Methanosaeta (1.81%), Sulfurovum (0.13%), and Thauera (0.15%) in BA, lower than their relative abundances in NA, which were much higher than 1.0%. However, for BSBA, compared to BA, these microbes have increased in different levels and at the same time showed slightly higher relative abundance than NA.
These results might imply an inhibition effect of JY inoculation in BA and further a positive stimulation of NH4Cl addition in BSBA on this microbial growth. It may be caused by the competitive inhibitory effect of JY inoculation for the growth of inoculant strains competing nutrients in the environment with indigenous flora (Wu et al., 2016). However, in BSBA, NH4Cl was added to supply more nitrogen nutrition for the growth of these microbes, and therefore an increase of these microbes occurred.
In addition, an outbreak of Escherichia-Shigella occurred in BA treatment with the relative abundance of 52.53%, but was lower than 2.17% in other samples. According to the knowledge of Escherichia-Shigella so far, species of Escherichia-Shigella were all pathogenic bacteria (Mwaikono et al., 2016). Also, the reason of this phenomenon still needs to be further studied.
Some eliminated microbes by petroleum contamination
There are some sequences affiliated to Bacillus, Gaiella, Nitrosococcus, Novosphingobium, Pseudomonas, Rubrobacter, Sphingomonas, and Ornatilinea higher than 1.0% in uncontaminated sample, but have a great decrement or even did not exist after remediation in NA, BA, and BSBA treatments (Fig. 5 and Supplementary Table S5), especially for Gaiella and Rubrobacter, which accounted for 4.26% and 2.76% in uncontaminated sample, but lower than 0.03% in other three treatments. Also, some microbes with the relative abundance lower than 1.0% in UCS, including Ilumatobacter, Lactococcus, Burkholderia, Bradyrhizobium, Nitrilruptor, Patulibacyer, Rhodobacter, Arthrobacter, Iamia, Nocardioides, and Solirubrobacter, were decreased significantly in the oil pollution and remediation treatments.
These enormous decrements of such a large number of microbes might have illustrated the marked decrease of microbial richness and diversity in petroleum-contaminated samples. What is more, previous researchers have proven that petroleum input can cause significant microbial community shift for selecting the specialized hydrocarbonoclastic bacteria suitable for local conditions (Bargiela et al., 2015).
Conclusion
This study demonstrated that BSBA and BA can accelerate petroleum degradation. BSBA showed the best on removal of TPH, saturates, and aromatics, but BA showed good performance on removal of higher molecular PAHs and even some polar components. Petroleum contamination has a selective effect on indigenous flora for stimulating growth of the oil-degrading and low temperature-tolerant microbes, including Comamonas, Arcobacter, Methanobrevibacter, Sulfurovum, Arenimonas, and Thauera, which lead to petroleum degradation in NA. Thus, the contaminating loess is a potential pool for petroleum-degrading strain isolation.
BA resulted in an apparent community shift compared to NA. This phenomenon might be caused by the competition of necessary nutrients between JY and indigenous microbes. The activity of some other low temperature-tolerant oil degraders were stimulated in this process, such as Variovorax, Methanobrevibacter, and Comamonas. And then resulted in a high degradation efficiency of PAHs and polar components. While BSBA showed a similar community structure with NA, which indicated NH4Cl addition alleviated the community shift resulted by JY inoculation, BSBA showed as the best strategy for TPH bioremediation.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Grant No. 31670510), Funding of State Key Laboratory of Loess and Quaternary Geology in China (Grant No. SKLLQG), and the Fundamental Research Funds for the Central Universities in China.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.