
Abstract
Abstract
In this study, anaerobic digestion was employed to degrade the refractory, high-salinity PO/MTBE (propylene oxide/methyl tertiary butyl ether) manufacturing wastewater (P/MMW) in sequencing batch mode. H2SO4 was added into the system to neutralize the alkalinity produced by the hydrolysis of sodium formate in P/MMW. The pH value in reactor A was only adjusted at the beginning of each cycle, and controlled in the range of 7.51–8.26. Meanwhile, the pH value in reactor B was maintained at 7.2 by regulating multiple times throughout the acclimation cycles. After five cycles of acclimation, both reactors exhibited stable removal of chemical oxygen demand (COD), while COD removal in system B was significantly improved in later stages (cycle III–V). More sulfate was reduced to sulfide alongside the COD removal process in reactor B, indicating more organic pollutants were removed by sulfate-reducing bacteria. High-throughput sequencing results illustrated that Firmicutes and other microorganisms capable of degrading the main pollutants in P/MMW were enriched to a greater degree in reactor B. Microbial communities in reactors A and B were different from seed sludge and evolved into two distinct groups, with sulfate and sulfide concentrations being main contributing factors for microbial community selection.
Introduction
Propylene oxide (PO) is the third derivative of propylene and a typical petrochemical. The propylene oxide/t-butyl alcohol (PO/TBA) process is widely employed for the selective production of PO, utilizing a complex production process and generating high yields of the coproduct methyl tertiary butyl ether (MTBE). MTBE is a fuel additive used almost exclusively in motor gasoline to raise the octane number and reduce carbon monoxide release in vehicle exhaust gases (Levchuk et al., 2014). Approximately 25 million tons of MTBE are produced worldwide annually (Roslev et al., 2015), and accordingly a large amount of PO/MTBE manufacturing wastewater (P/MMW) is generated during PO/MTBE production.
P/MMW contains many refractory organic compounds, such as t-butyl-formate (TBF), TBA, t-butyl isopropyl ether, and dichloropropane (Mitani et al., 2002; Wang et al., 2017), potentially contaminating groundwater and surface water sources. Furthermore, TBF can hydrolyze to formic acid [Eq. (1)] when NaOH is added in the etherification reaction system to neutralize the acidity. This resulted in the formation of sodium formate [Eq. (2)] and high salinity in P/MMW. While the hydrolysis of sodium formate would produce a large amount of OH−, it is, therefore, in urgent need of pH adjustment, which brings some challenges to the P/MMW treatment.
Previous studies have mainly focused on removing specific pollutants, such as degrading PO using activated sludge (Wang et al., 2017) or MTBE by advanced oxidation (Vaferi et al., 2014). Few reports had been devoted to the integrated treatment of P/MWW. Anaerobic digestion (AD) process is one of the most extensively used technologies for wastewater treatment, which has the advantage of being energy efficient, cost effective, and environmentally friendly (Kim et al., 2013; Li et al., 2015). But the refractory, high-salinity characteristics of P/MWW pose many challenges for AD process. Moreover, the anaerobic process is extremely vulnerable to the variation of pH value (Latif et al., 2017), and therefore it is important to investigate the pH-regulation mode.
The pH value in anaerobic reactors is always adjusted within a specified range and continuously monitored. Two different pH-regulation modes were commonly used during practical production processes: adding certain amount of acid or alkali once at the beginning of every cycle or multiple times throughout each cycle. The former is easy to operate, but the system pH fluctuates inevitably. In contrast, the later can always maintain the pH value within the desired range, but it has a higher requirement for operation control. This study aims to investigate the AD processing of P/MMW in the two pH-regulation modes mentioned above, further compare their performance, and explore the reasons. The dynamic evolutions of microbial communities were revealed using high-throughput sequencing technology. The relationships among pH-regulation mode, microbial community structure, and system performance were fully investigated.
Materials and Methods
Characteristics of P/MMW
P/MMW was obtained from a PO/MTBE-manufacturing factory (Nanjing Jinling Huntsman New Material Co., Ltd., Jiangsu Province, China). P/MMW was transported to the laboratory and stored at 4°C before use, and the physicochemical characteristics are presented in Table 1.
Physicochemical Characteristics of Propylene Oxide/Methyl Tertiary Butyl Ether Manufacturing Wastewater
BOD, biochemical oxygen demand; COD, chemical oxygen demand; ORP, oxidation reduction potential; TDS, total dissolved solids; TN, total nitrogen; TOC, total organic carbon; TP, total phosphorus; VFA, volatile fatty acid.
Experimental set-up
Batch experiments were performed in two identical 1-L sealed flasks (working volume of 700 mL) to investigate the influence of different pH-regulation modes on AD treatment of P/MMW. Before use, P/MMW was diluted with deionized water at a 1:1 (v/v) volume ratio as the influent. The v/v ratio of influent to granular seed sludge in both flasks was 1:1. The seed granular sludge was gathered from a starch wastewater treatment plant operating using upflow anaerobic sludge blanket technology. The average particle diameter of the granular sludge was greater than 1 mm, while the average mixed liquor suspended solid (MLSS) and mixed liquor volatile suspended solid (MLVSS) were 37.3 and 31.7 g/L, respectively.
Both reactors were sparged with N2 gas to maintain anaerobic conditions, then incubated on a rotary shaker with rotation speed of 110 rpm at 38°C. Chemical oxygen demand (COD) was taken as the indicator for acclimation process. Once the system COD dropped to the bottom, an acclimation cycle was considered to be completed. After settling the sludge for 30 min, the supernatant was completely decanted and fresh P/MMW was infused. Thus, a new acclimation cycle was started. Five acclimation cycles were carried out in both reactors (denoted as cycles I–V), and lasted for 43 days in total. The hydraulic retention time in the two reactors was 5–9 days and the organic load rates were 1.2–2.2 kg COD/(m3·d).
Two different pH-regulation modes were used in the two flasks, respectively, as follows: reactor A: 9.5 mL H2SO4 (10%, vol%) was added to the system at the beginning of each cycle (one-time regulation); reactor B: the pH was adjusted every 3–6 h to around 7.2 by H2SO4 (10%, vol%) with the dosage as needed (multiple-times regulation).
Analytical methods
Effluent samples were collected from both reactors at regular intervals and centrifuged at 8,000 rpm for 5 min before analysis. COD, biochemical oxygen demand, NH3-N, MLSS, MLVSS, volatile fatty acid (VFA), total dissolved solids, and total phosphorus concentrations were analyzed according to the Standard Methods for Water and Wastewater Examination (APHA, 2012). The pH and oxidation reduction potential were measured using a pH meter (HQ30d; HACH, America). Total organic carbon (TOC) and total nitrogen concentrations were determined using a TOC analyzer (TMM-1; Shimadzu, Japan). The sulfate concentration was analyzed using the barium chromate spectrophotometric method and dissolved sulfide (Sun et al., 2018) was measured using the methylene blue spectrophotometric method (APHA, 2012). All measurements were performed in triplicate unless otherwise indicated. All statistical analyses were performed using Statistical Product and Service Solutions (version 18.0).
Microbial community analysis
Seed sludge (S0) and sludge samples collected at the end of cycle I, III, and V from reactor A (A1, A3, and A5) and B (B1, B3, and B5), were sent for microbial community structure analysis using the same method in Wan et al. (2018). Primers 341F and 805R were used to amplify the V3–V4 region of bacterial 16S rRNA gene sequences. The raw sequences were deposited into the NCBI short reads' archive database under accession number of SRP135950. The raw 16S rRNA gene sequences were separated by barcode for quality control, using Flash software (version 1.2.7) to merge the paired-end reads. Low-quality sequences were removed using Prinseq software and UCHIME was used to remove chimeric sequences from the tags.
After quality filtering, 22,385–31,392 high-quality reads were retained from the 7 samples, with an average length of 414 bp. The sequence read sets were aligned using the SILVA 16S rRNA gene reference database and clustered into operational taxonomic units (OTUs) with 97% similarity. The number of OTU obtained from the 7 samples varied from 1,612 to 2,727. Phylogenetic affiliation of each 16S rRNA gene sequence was performed using the Ribosomal Database Project (RDP) classifier. Alpha diversity indices and Good's coverage were calculated and analyzed using MOTHUR. Nonmetric multidimensional scaling (NMDS) was performed using R software (version 3.2) to illustrate the relationship between samples. Canonical correspondence analysis (CCA) was also performed using R software, to analyze the impact of physicochemical attributes on community structure dynamics.
Results and Discussion
Operational performance of anaerobic reactors
Both of the acclimation processes were repeated for five cycles, with the variations in COD, COD removal efficiency, and pH values monitored under different pH-regulation modes (Fig. 1). The initial pH values of reactors A and B were both 5.95. Due to the alkalinity produced by the hydrolysis of sodium formate, the pH in reactor A increased to the maximum observed level (8.26) in cycle I, followed by fluctuation between 7.51 and 7.89 during the subsequent cycles. Conversely, the pH value in reactor B was always maintained at a constant level of 7.2.
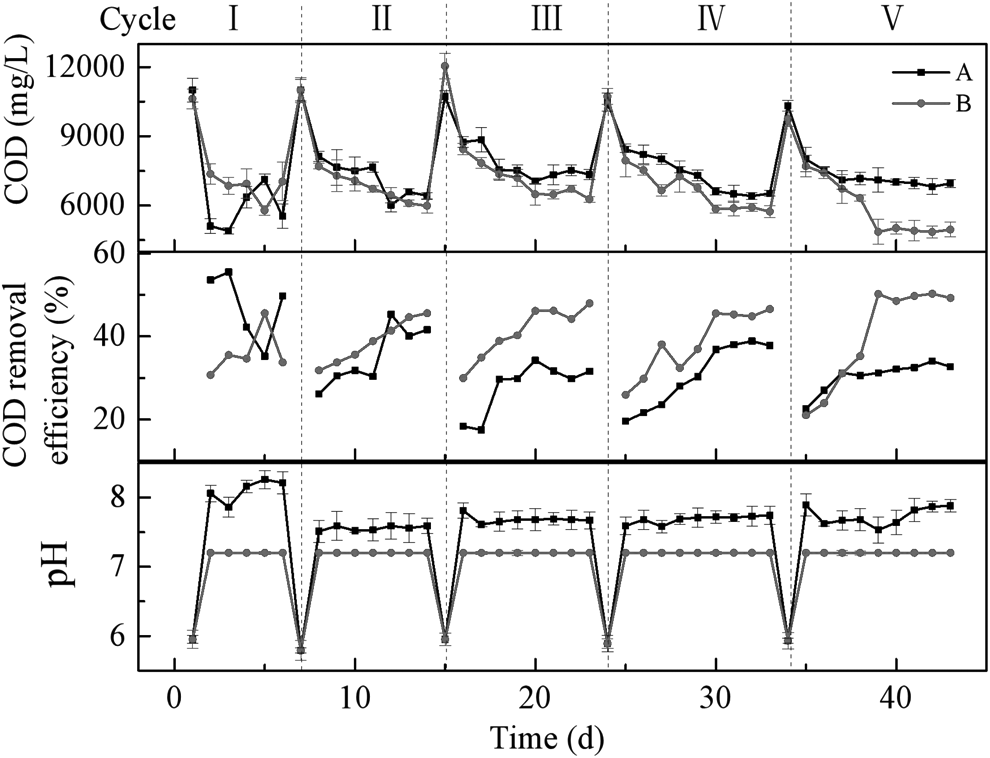
Variations of COD, COD removal efficiency, and pH in different acclimation cycles. COD, chemical oxygen demand.
In the early cycles, the COD levels in both reactors rapidly decreased, suggesting that AD processes could effectively remove organic pollutants. However, the COD removal efficiencies in both reactors were unstable, indicating that the living conditions were not favorable for sludge-derived microbial communities due to the refractory, high-salinity P/MMW. From cycle III onward, the organic matter removal efficiency reduced slightly in reactor A, while it increased progressively in reactor B and reached a maximum level of 50% at the end of cycle V. Meanwhile, the time required for COD levels to drop from their maximum to minimum values, reduced gradually (6 days in cycle III and 5 days in cycle V), indicating that seed sludge was well adapted to the P/MMW environment. In cycles III–V, the COD removal efficiency of reactor B was significantly higher than that of reactor A (p < 0.05).
Different dosages of H2SO4 were added into reactor A and B according to their respective pH-regulation modes. From Fig. 2, the initial influent sulfate concentrations of A and B were 2,408 and 4,013 mg/L, respectively. During the treatment period, the effluent sulfate concentrations reduced appreciably, whereas the sulfide concentrations increased in both reactors. This phenomenon indicates the existence of sulfate-reducing bacteria (SRB), which are able to reduce sulfate to sulfide using organic compounds (such as carbohydrates, organic acids, alcohols, aliphatic hydrocarbons, and aromatic hydrocarbons) or hydrogen as electron donors (Chen et al., 2013; Huan et al., 2013). Furthermore, both the consumption of sulfate and the decomposition of organic matter can cause an increase in carbonate alkalinity (Baumgartner et al., 2006), resulting in an increase of pH value. In particular, the sulfate content in reactor A reduced from 2,408 to 165 mg/L in cycle I, which may explain the peak in pH value observed in this cycle.

Variations of sulfate and sulfide concentrations in different acclimation cycles.
A high level of sulfate reduction was observed while simultaneous sulfide generation was comparatively low, which was in accordance with previous studies (Dudeney and Sbai, 1993; Kaufman et al., 1997; Salo et al., 2018). The reasons for this phenomenon were speculated as follows: first, sulfide may volatilize easily after sampling, which is linked with low sulfide production compared with sulfate reduction. Second, the P/MMW used in this study is real industrial wastewater and contains many complex organic compounds, such as metal ions, which can form precipitation with the sulfide. Richards and Pallud (2016) demonstrated that sulfide precipitation in sediment is a key sink for the sulfide generated through sulfate reduction. Furthermore, it is likely that the dissolved sulfide may be further oxidized to elemental sulfur by autotrophic sulfide-oxidizing bacteria (Zhang et al., 2018).
High-throughput sequencing analysis
Richness and diversity of microbial communities
The high-throughput sequencing primers applied in this study mainly targeted bacteria and therefore, the sequencing results may not fully reflect the archaeal community. The coverage values of all seven samples ranged from 94.81% to 97.37% (Supplementary Table S1), suggesting the sequence libraries constructed in this study adequately accounted for the bacterial community. The Venn diagrams (Supplementary Fig. S1) illustrate that only 205 OTUs were shared by all the 7 samples, revealing that the microbial community structure altered appreciably. Although over 22,000 reads were obtained from all 7 samples, the rarefaction curves did not reach a plateau (Supplementary Fig. S2), indicating the samples contained highly diverse bacterial communities and numerous rare species.
Chao1 and ACE indices are both positively related to microbial richness and are commonly used to estimate the total number of species in a community. Shannon and Simpson indices are mainly used to characterize the diversity of microbial communities. The microbial diversity increases with the increasing in Shannon index and the decreasing in Simpson index. In cycle I, the microbial richness reduced in reactor A and increased in reactor B, while the microbial diversity increased slightly in both A and B reactors. The fluctuations in microbial community structure indicate that microbial adaptation occurred in an attempt to survive in the highly osmotic oligotrophic environment caused by P/MMW. From cycle III onward, the diversity and richness in both reactors reduced, as under the constant selective pressure posed by P/MMW, and microbial populations that could not adapt were eliminated.
Taxonomic complexity of bacterial community
To get an insight into microbial community dynamics and evolution during the treatment of P/MMW, the sequences from the dominant microbial communities over time in each sample are presented at the phylum and genus levels. Figure 3a shows that the seven samples were dominated by Chloroflexi, Proteobacteria, Firmicutes, Euryarchaeota, and Bacteroidetes, which altogether accounted for more than 60% of all sequences. Previous studies have reported similar results, with these phyla being widespread in anaerobic reactors treating food waste (Goux et al., 2016), municipal sewage (Shu et al., 2015; Beale et al., 2016), and pulp and paper wastewater (Fykse et al., 2016). Compared with S0, the proportion of Firmicutes increased in both reactors during the treatment of P/MMW, with a prominent continuous upward trend particularly apparent in reactor B. Firmicutes can break up large macromolecules, such as proteins, complex fats, and polycarbohydrates into amino acids, long-chain fatty acids, and monosugars, among other products (Li et al., 2013). Guisado et al. (2015) isolated three bacterial strains affiliated to Firmicutes with the capacity to degrade MTBE (30–100 mg/L). Firmicutes was also a dominant phylum in PO saponification wastewater residual sludge, accounting for about 10.58% of all sequences (Wang et al., 2017).

Taxonomic classification of microbial communities at phylum
The genus assignment results (Fig. 3b) demonstrate that the microbial communities in both P/MMW treatment systems (A and B) were similar, while being dramatically distinct from the seeded sludge (S0). Sulfurovum, Levilinea, Longilinea, Syntrophobacter, Bellilinea, and Ornatiline were the primary genera in S0, and accounted for 60% of the whole sequences. Sulfurovum (16.68%) was almost eliminated in both reactors (<0.01%) during the acclimation process, while the later five genera still accounted for more proportions in both reactors. Sulfurovum is an obligatory aerobic thiosulfate or sulfur-oxidizing bacterium (Inagaki et al., 2004), it may be not suitable for growth in P/MMW. Syntrophobacter can oxidize hydrogen and formate with sulfate as an electron acceptor (Hu et al., 2015), with the proportion of this SRB increasing from 5.24% (S0) to 10.44% (A1) and 8.49% (B1), as formate is one of the main substances in P/MMW. However, the abundance of Syntrophobacter in both reactors reduced rapidly to <4.5% as the sulfide concentration in both reactors gradually accumulated (>450 mg/L) from cycle III to V. Lopes et al. (2010) reported a similar phenomenon, proposing that high sulfide concentrations (100 mg/L) have a highly inhibitory effect on SRB.
In comparison with S0, some functionally important populations, such as Brachymonas, Acetoanaerobium, Tissierella, Fusibacter, Acinetobacter, and Arcobacter were more abundant in reactors A and B. Brachymonas increased sharply in reactor A, while the remaining genera increased rapidly in reactor B. Brachymonas is capable of degrading alicyclic hydrocarbons, cyclohexane, cyclopentane, and their methyl derivatives (Rouviere and Chen, 2003). Fusibacter is a moderate salt-tolerant genus, and Serrano et al. (2017) had isolated a Fusibacter sp. 3D3 with the capacity of biodegrading hydrocarbon from a hypersaline salt flat in Northern Chile. Acinetobacter and Arcobacter are related to the metabolism of sulfur-containing compounds (Mente et al., 2006; Zhang et al., 2015), with these characteristics promoting their enrichment in reactors A and B accordingly. Moreover, 18.38–34.63% of the total indentified sequences could not be classified at the genus level for the seven samples. This revealed that studies on microorganism genome were still limited due to the great variety, extensive distribution, and complex metabolism of microorganisms.
Cluster analysis of microbial communities and impact factors on microbial community structure shift
NMDS was conducted to visualize the overall similarities and differences in microbial community structures among all seven samples. According to Fig. 4a, the microbial communities of the seven samples were clustered into three groups (S0, A1, and B1; A3 and A5; B3 and B5). NMDS1 appears to separate the samples based on the pH-regulation modes. A1 and B1 were located more closely with S0, because the community structure changes were not distinct enough within 5 days of acclimation (cycle I). With extended acclimation time, microbial populations in reactors A and B obviously evolved in two separate directions due to altered environmental conditions, potentially as a result of their specific pH-regulation modes. Ma et al. (2019) also reported that bacterial communities were influenced significantly by pH during AD of food waste, with the bacterial communities at pH of 4–8 and 9–11 being clearly separated into two groups, which differed with seed sludge.

Comparison of the microbial communities in all seven samples based on the NMDS analysis
CCA was adopted to evaluate potential effects of environmental factors on the dynamics of microbial populations in AD systems. As displayed in Fig. 4b, the microbial communities in reactors A and B deviated from those of seed sludge, in response to four physiochemical factors, such as pH, COD, sulfate, and sulfide contents. pH was positively associated with the microbial communities of B1, A1, A3, and A5, while negatively correlated with the rest of the microbial communities. Moreover, the obvious divergence between community structures in reactors A and B from cycle III–V, were mainly caused by the sulfate and sulfide concentrations.
Relationships among different pH-regulation modes, microbial community structure, and system performance
The AD performance of reactor B (multiple-times regulation) was significantly improved as compared with A (one-time regulation), especially in the later stages of acclimation (cycles III–V). Based on these experimental results, the reasons for this effect were speculated as follows:
First, reactor B provided a more ideal and stable pH (7.2) for methanogens, which are highly sensitive to pH conditions, as supported by the results of Lu et al. (2015), reporting that the optimum pH for AD production of methane was 6.8–7.2. Conversely, the pH in reactor A continually fluctuated and were slightly alkaline (>7.5), deviating from the optimal pH range. Second, multiple-times regulation caused more H2SO4 to be introduced to reactor B compared with reactor A. SRB can perform anaerobic respiration utilizing sulfate as a terminal electron acceptor and oxidize organic compounds (Huan et al., 2013). Increased sulfate consumption in reactor B may also contribute to the improved COD removal efficiency observed in this system.
Furthermore, Firmicutes and other microorganisms active in the degradation of pollutants in P/MMW were more abundant in reactor B, improving its efficiency when treating this kind of wastewater. Overall, although multiple-time regulation is less convenient to practically implement, it encourages faster acclimation and AD reactor stability, resulting in improved COD removal compared with one-time regulation.
Conclusions
AD process can effectively remove the refractory organic pollutants from P/MMW, with the COD removal efficiency reaching 50% in the later stages of acclimation (cycle III–V). The pH value in reactor A was only adjusted at the beginning of each cycle, but the pH value in reactor B was regulated multiple times. Provision of favorable pH in reactor B could enhance the methanogenesis and sulfate reduction, thus exhibiting better performance than reactor A. High-throughput sequencing results revealed that the microbial communities under different pH-regulation modes evolved into distinct groups, in which sulfate and sulfide concentrations were the main factor for this deviation.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (No. 51508517, 51208179). The authors gratefully acknowledge the technical assistance provided by ZhiHe Environmental Science and Technology Co., Ltd.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.