
Abstract
Abstract
Effluents from textile and dyeing industries are typical hypersaline wastewaters, which also contain high concentration of dyes (most are azo types). To achieve effective and environment-friendly treatment, a laboratory-scale activated sludge-based membrane bioreactor (MBR), which was bioaugmented by a halotolerant yeast Pichia occidentalis G1, was constructed and assessed through a long-term operation. Dynamics of microbial community during the operation were analyzed by high-throughput sequencing method. The results demonstrated that the bioaugmented MBR was more stable and efficient for decolorization, chemical oxygen demand removal, and detoxification, as well as stronger tolerance to shock loading than the nonaugmented control. Sludge concentration in the yeast-augmented MBR was also higher than the other two controls, which might ensure its more stable and higher treatment efficiency. MiSeq sequencing results suggested that the most abundant fungal genus in the bioaugmented MBR were identified as Pichia, indicating the possibly successful colonization of strain G1 or some other species belonging to the same genus with high metabolic activity. Fungal species belonging to Candida were also enriched with bioaugmentation. Rhodanobacter was the most dominant bacterial genera in the bioaugmented system, which might share acidic conditions with fungi belonging to Pichia. This method was potentially effective in bioremediation of hypersaline industrial wastewaters, which contained azo dyes.
Introduction
Textile wastewaters, which always consist of considerable amounts of contaminants such as dyes, suspended solids (SS), additives, detergents, and surfactants, may cause serious pollution due to their discharge without effective purification. Among these components, synthetic dyes are the major group of organic pollutants, which can cause high chroma and low biodegradability (Prigione et al., 2008). Azo dyes are the most widely used types, which contain one or more azo bonds (-N = N-), as well as aromatic rings and sulfonic groups in their chemical structures (Arun Prasad et al., 2013; Wang et al., 2013). As reported, more than 10% of azo dyes are lost during dyeing processes and then released into industrial effluents (Pearce et al., 2003).
Azo compounds may destroy the water ecosystem, since they drastically decrease oxygen concentration and light penetration (Durruty et al., 2015). Meanwhile, the breakdown products from degradation of azo dyes are even more harmful to organisms due to their toxicity, teratogenicity, and carcinogenicity (Lim et al., 2013). Furthermore, wastewater streams from dye fixation processes generally contain a high load of salts, which can cause more damage to the environment and biological treatments (Silva et al., 2009). Therefore, effective treatment of textile wastewaters before discharge to the environment is significantly important.
Many chemical and physical techniques were used to remove azo dyes. For instance, activated carbon was widely used for removing azo dyes from simulated or real industrial wastewaters through physical adsorption (Al-Degs et al., 2000; Pal and Deb, 2014). In addition, advanced oxidation processes such as Electro-Fenton method were also efficient for removing azo dyes through chemical degradation (Jiang et al., 2016; Özcan and Özcan, 2018). However, the shortcomings such as high energy consumption and sludge production, and formation of secondary pollutions limit the wide application of these methods (Tamboli et al., 2010). By contrast, biological techniques are widely used due to their environmentally friendly and cost-effective characteristics (Du et al., 2011).
Many microorganisms, mainly including bacteria and fungi, are capable of removing azo dyes from wastewaters through adsorption or biodegradation (Banat et al., 1996). According to the previous reports, bacteria could reductively cleave azo bonds by azoreductases, most of which are specific enzymes (Keck et al., 2002; Kumari et al., 2016). By contrast, fungi can degrade or even mineralize many recalcitrant organics depending on their nonspecific and nonstereoselective ligninolytic enzymes, including laccase, manganese peroxidase, and lignin peroxidase (Gomi et al., 2011).
For instance, white rot fungi such as Phanerochaete chrysosporium and Trametes versicolor have been confirmed as efficient degraders of azo dyes (Solís et al., 2012), some ascomycota fungi (yeasts) such as Candida tropicalis and Magnusiomyces ingens could also be capable of degrading and even detoxifying azo dyes efficiently (Tan et al., 2013, 2014). Furthermore, it was indicated that co-cultures of fungi and bacteria displayed higher azo dye decolorization efficiency than either of them (Su et al., 2009; Gou et al., 2009; Qu et al., 2010).
Some researches even found that the fungal-bacterial coculture could effectively mineralize some of the azo dyes, which is significantly important for removing recalcitrant organics by biological processes. For instance, over 50% of 100 mg/L Reactive Brilliant Red X-3B could be mineralized within 12 h through a series of anaerobic-aerobic process by the coculture of Penicillium sp. QQ and Exiguobacterium sp. TL (Shi et al., 2014). The possible mechanism was described as, in fungi-bacteria cocultures, fungi could decompose azo dyes and their aromatic breakdown byproducts into small-molecule substances to improve the biodegradability of byproducts, then bacteria further degraded or even mineralized most of the residual organics (Li et al., 2015; He et al., 2017).
The synergistic effects of fungi and bacteria during biodegradation lead to high treatment capability and stability, thus suggesting a potentially effective alternative for field application. However, most of the related researches were done under low-salt conditions, which could not reflect the effectiveness of the method under practical conditions.
As mentioned above, textile wastewaters always contain high load of salts, which are extensively used during dye fixation processes (Peyton et al., 2002). High concentration of salts can reduce or even inhibit the metabolic activity of nonextremophilic microbes, thus causing poor efficiency of biological treatments. To resolve this issue, halotolerant or halophilic microorganisms are chosen for bioprocesses since they can maintain relatively high metabolic activity in hypersaline environments (Castillo-Carvajal et al., 2014). Some halotolerant bacteria have been utilized for treatment of dye-contaminated wastewater under hypersaline conditions (Tan et al., 2009).
For example, Psychrobacter alimentarius KS23 and Staphylococcus equorum KS26, which were isolated from seawater sediment, could decolorize three reactive dyes efficiently under a wide range of salt concentrations (Khalid et al., 2012). Shewanella aquimarina had also been confirmed as an efficient halotolerant bacterium coping with dye-polluted wastewaters (Meng et al., 2012). On the other hand, some halotolerant fungi have also been exploited for treating hypersaline wastewaters. For instance, Tan et al. (2016) and Song et al. (2017) isolated two halotolerant yeasts Scheffersomyces spartinae TLHS-SF1 and Pichia occidentalis G1, both of which could efficiently degrade and detoxified various azo dyes under hypersaline conditions.
In addition, it was also reported that a Candida strain JS3, which was isolated from industrial activated sludge (AS), could degrade phenol under saline conditions (Jiang et al., 2015). Therefore, these halotolerant fungi can be used for constructing fungal-bacterial cocultures coping with the purification of hypersaline recalcitrant industrial wastewaters. Furthermore, effectiveness of fungal-bacterial cocultures should be examined in at least laboratory-scale reactors during a long-term operation.
In this study, long-term effectiveness of a fungal-bacterial coculturing system, which was composed of a novel isolated halotolerant yeast P. occidentalis G1 and AS, was assessed in a laboratory-scale submerged membrane bioreactor (MBR). Another two MBRs, which were inoculated with pure culture of the yeast G1 and only AS, respectively, were simultaneously operated for control. Furthermore, MiSeq sequencing analysis was used to analyze the dynamics of microbial consortia in three MBRs, to monitor the shifts of the yeast G1 as well as other dominant microorganisms. The corresponding results were expected to investigate the bioaugmentation effectiveness of the halotolerant yeast during a long-term operation and provide useful clues for the bioremediation of other recalcitrant organic pollutants.
Materials and Methods
Reagents
The target dye Acid Red B (ARB) with higher than 99% purity and biochemical reagents with electrophoretical purity were purchased from Sangon Biotech (Shanghai) Co., Ltd. Other chemical reagents used in this study are of analytical grade.
Pretreatment of microbes
A previously isolated yeast strain P. occidentalis G1, which was capable of aerobically degrading various azo dyes under hypersaline conditions, was used in this study (Song et al., 2017). The cells of the yeast G1 were enriched and washed with 0.02 mol/L phosphate buffer (pH = 7.2) three times before inoculation as described by He et al. (2017). AS was purchased from Chunliu River municipal sewage treatment plant (Dalian, China) and was acclimatized for 30 days before inoculation with the medium containing (g/L) glucose 2.0, (NH4)2SO4 0.6, MgSO4·7H2O 0.5, NaCl 30.0, and ARB 0.05–0.2 (initial pH was 6.0).
Operation of pre-experiment in flasks and MBRs
A group of pre-experiment was performed in flasks under the conditions of 25°C (temperature) and 160 r/min (rotation speed) for determining the inoculation ratio between AS and the yeast G1 in the bioaugmented MBR. It was performed in the form of sequencing batch tests in five flasks, which was inoculated with AS (the concentrations were all 4,000 mg/LSS) and the yeast G1 (0, 500, 1,000, 1,500, and 2,000 mg/L SS), respectively. The whole operation contained four batches with initial ARB concentration of 200 mg/L (during the first two batches) and 300 mg/L (during the next two batches), corresponding to the reaction time of 24 and 32 h for each batch, respectively. Centrifugation (10,000 g for 10 min) was used for liquid-solid separation at the end of each batch instead of membrane separation.
Basing on the pre-experiment, three laboratory-scale cubic MBRs with a working volume of 2 L, which were 160 mm in length, 80 mm in width, and 190 mm in height, were constructed and operated to estimate the long-term effectiveness of the AS system bioaugmented with the yeast G1. The flow chart is shown in Supplementary Fig. S1. The nonwoven fabric membrane module was 62 mm in length and 42 mm in width with the effective area of 50 mm × 30 mm × 2 surfaces and the membrane pore size of 0.22 μm. Three MBRs were named as 1#, 2#, and 3#, which were inoculated with 5,000 mg/L AS, 5,000 mg/L the yeast G1, and both of them (4,000 mg/L AS and 10,00 mg/L the yeast G1), respectively, which were determined through pre-experiments in flasks.
The simulated wastewater contained (g/L) glucose 2.0, (NH4)2SO4 0.6, MgSO4·7H2O 0.5, NaCl 30.0, and ARB 0.2–0.3. The MBRs were operated at room temperature (about 25–28°C) and the whole process was divided into two stages. In stage I (0–30 days), the initial concentrations of ARB were 200 mg/L and the hydraulic retention time (HRT) was 24 h; then, in stage II (31–60 days), the influent concentration of ARB was increased to 300 mg/L with the same HRT. Besides, the dissolved oxygen (DO) was controlled at 2.5–3.0 mg/L by air pumps with gas disperser and flow meter.
Concentrations of ARB and chemical oxygen demand (COD) in influent and effluent were monitored once a day for estimating the treatment effectiveness of MBRs. Besides, acute toxicity of influent and effluent at the end of each operation stage was also analyzed to estimate the detoxification effectiveness. Microbes in three MBRs were gathered at the end of each operation stage for analyzing the dynamics of microbial (including bacterial and fungal) communities.
Analytical methods
Concentration of ARB was measured with an ultraviolet (UV)-Vis spectrophotometer and decolorization percentages were calculated according to Tan et al. (2013). COD was analyzed by the standard method “microwave digestion method” as described by Shi et al. (2015). Concentration of mixed liquor suspended solids (MLSS) was analyzed gravimetrically according to Xu et al. (2015). In addition, the acute toxicity of the influent and effluent was analyzed by the stand method “Microtox bioassays” (GB/T 15441-1995), which were performed with the luminescent bacteria Vibrio fischeri (NRRL B-11177) according to Li et al. (2015).
High-throughput sequencing
Genomic DNA of the samples was extracted and determined according to the previous study (Li et al., 2015). Then the DNA was amplified by polymerase chain reaction (PCR) with the primers of 515F/907R (GTGCCAGCMGCCGCGG/CCGTCAATTCMTTTRAGTTT) and 1737F/2043R (GGAAGTAAAAGTCGTAACAAGG/GCTGCGTTCTTCATCGATGC) for V4–V5 region of 16S ribosomal RNA (rRNA) gene (bacterial community) and ITS1-ITS2 region of 18S rRNA gene (fungal community), respectively (He et al., 2017). PCR amplification of each sample was performed in triplicate and pooled, and PCR products were purified before sequencing by the method described by Shi et al. (2015). High-throughput sequencing was performed by Sangon Biotech (Shanghai) Co., Ltd. with an Illumina MiSeq platform (Illumina).
Statistical and bioinformatics analysis of sequencing results were performed on the basis of previous procedures.
Statistical analysis
All data were analyzed in triplicate and statistically analyzed by one-way analysis of variance (ANOVA) with Microsoft Excel 2010 software.
Results and Discussion
All the experimental values for plotting (Figs. 1–3) are in the form of “mean value of three experiments ± standard error of mean.” The p-value of <0.001 suggested that all the experimental data were significantly different from control by one-way ANOVA with Tukey-Kramer comparison test.
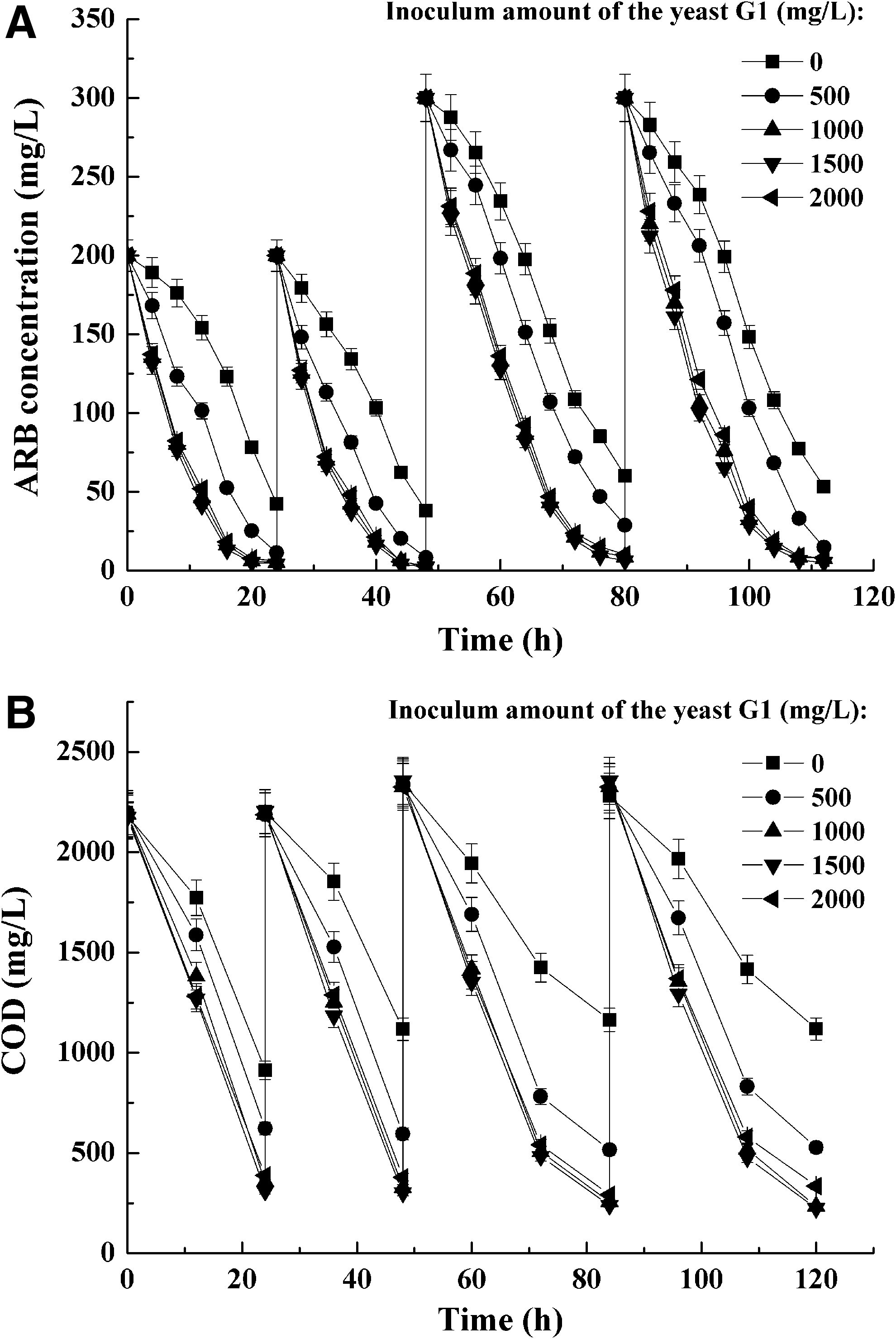
Pre-experiments in the form of batch tests for determining optimal inoculum amount of the yeast G1.

Performance of three MBRs.

The acute toxicity of the influent and effluents at the end of stages I
Pre-experiment for determining inoculum amount of the yeast G1
Fig. 1 displayed the results of the pre-experiment performed in flasks. It was shown in Fig. 1A that about 60.9% of 200 mg/L ARB was decolorized by the nonaugmented AS system during the first batch of treatment (20 h); meanwhile, about 87.3% decolorization percentage was achieved in the AS system bioaugmented by 500 mg/L of yeast G1. By contrast, more than 96.5% of 200 mg/L ARB was decolorized in the AS systems bioaugmented with 1,000–2,000 mg/L of yeast G1. It was suggested that decolorization efficiency by the AS system was improved through bioaugmentation and the highest decolorization efficiency was achieved with at least 1,000 mg/L of yeast G1.
During the following three batches of treatment (with initial ARB concentration of 200, 300, and 300 mg/L, respectively), ARB decolorization percentages of the nonaugmented AS system were still lower than those of the yeast-augmented ones, which suggested that bioaugmentation with the yeast G1 obviously promotes ARB decolorization. Comparatively, the optimal decolorization percentages were achieved with at least 1,000 mg/L of yeast G1. On the other hand, changes of COD during the corresponding processes were also monitored and the result is shown in Fig. 1B. Similar to decolorization processes, the nonaugmented AS system displayed lower COD removal efficiency than the bioaugmented ones.
The results also showed that the optimal COD removal performances were achieved with at least 1,000 mg/L of yeast G1 in the bioaugmented AS system. Therefore, the final inoculum amount of yeast G1 was determined as 1,000 mg/L in 4,000 mg/L AS.
Performance of MBRs
Color removal
It was indicated that reductive cleavage of azo bonds was always the first step for biodegradation of azo dyes under either aerobic or anaerobic condition, which resulted in decolorization (Tan et al., 2009, 2013). Thus, color removal processes were monitored. Color removal performance of three MBRs is shown in Fig. 2A and B. At the beginning, 3 days, ARB concentration in effluents from 1#, 2# and 3# MBRs was about 45.2, 6.5, and 7.1 mg/L, corresponding to the decolorization percentages of about 77.4%, 96.8%, and 96.5%, respectively. However, ARB concentration in the effluents from all the three MBRs slightly increased from the 4th day. Then, decolorization efficiency of all MBRs started to recover during the following 6–10 days.
The excessive decolorization percentages at the beginning of operation were probably due to the adsorption of ARB on AS, according to the explanation in previous report (He et al., 2017). An extended experiment was performed in three flasks, which were inoculated with autoclaved AS, the yeast, and the coculture with the same MLSS corresponding to the three MBRs, respectively, for estimating color removal performance through adsorption. The result (data not shown) showed that about 17.3–21.6% color was removed through adsorption in 1# and 3# MBRs during the first 4 days; in contrast, color removal percentages by 2# MBR were only 3.5–5.7%. After operation for 5 days, decolorization percentages in all the three flasks approached to 0, suggesting that the adsorption processes saturated.
In comparison with 2# and 3# MBRs, the effluent ARB concentration of 1# MBR was obviously lower during stage I (shown in Fig. 2A). After the fluctuation of decolorization percentage curves (Fig. 2B) at the beginning of treatment, decolorization percentages of 1#, 2#, and 3# MBRs were finally stabilized at about 65.5%, 92.0%, and 94.5% at the end of stage I, respectively. Then influent ARB concentration was increased to 300 mg/L in stage II for estimating the resistance to organic shock loading of three MBRs. Among all the three MBRs, 1# MBR also showed the lowest decolorization percentages of about 70.5% at the end of stage II. By contrast, about 87.3% and 95.2% of ARB was decolorized in 2# and 3# MBRs at the end of stage II, respectively.
It was suggested that 3# MBR, which was bioaugmented by yeast G1, showed higher decolorization efficiency and resistance to shock loading than the other two inoculated with only AS or the yeast G1.
COD removal
Concentration of organic carbon, which was generally represented by COD or total organic carbon (TOC), was also important quality index of wastewaters (Bilal et al., 2016), thus organics (chose COD as target in this study) removal was also analyzed. COD removal performance of three MBRs is shown in Fig. 2C and D. COD in the effluent from 1# MBR ranged from 455.0 to 900.0 mg/L in stage I; by contrast, the corresponding ones from 2# and 3# MBRs were 670.0–1,050.0 mg/L and 160.0–445.0 mg/L, respectively (Fig. 2C). The corresponding COD removal percentages in 1#, 2#, and 3# MBRs were 58.6–79.1%, 48.5–68.2%, and 76.9–92.1%, respectively (Fig. 2D).
At the beginning of stage II, COD removal efficiency in all the three MBRs decreased due to the increase of influent ARB concentration from 200 to 300 mg/L. Then, from the 5th day of stage II, COD removal percentages in three MBRs started to increase again. The effluent COD at the end of stage II of 1#, 2#, and 3# MBRs was about 680.0, 982.8, and 286.3 mg/L, corresponding to the COD removal percentages of 69.1%, 55.4%, and 88.2%, respectively. These results suggested that the yeast-augmented 3# MBR also showed better performance for COD removal than the other two. In addition, 2# MBR was less efficient for COD removal than 1# MBR, although it showed higher decolorization efficiency.
The reason might be that, although the yeast G1 was efficient for decolorization and detoxification of ARB, it would be less efficient for mineralizing complex and mixed organic substrates in comparison with AS, which contained diverse microorganisms (Shi et al., 2015). On the other hand, it was indicated that pure cultures might not survive in long-term operating bioreactors because they could not adapt complex environmental conditions. Thus, microbial consortia are essential for removing most of organic carbon in practical application (Ruiz-Arias et al., 2010).
Sludge concentration (MLSS)
Variation of sludge concentration, which was represented by MLSS in three MBRs, was also monitored because higher treatment efficiency might depend on higher biomass. It was shown in Fig. 2E that MLSSs in all the three MBRs decreased at the beginning. After operating for 4–8 days, MLSSs increased again and those in 1#, 2#, and 3# were finally stabilized at 3,870–3,950, 4,330–4,450, and 4,590–4,810 mg/L at the end of stage I, respectively. Then at the beginning of stage II, MLSSs in all the three MBRs decreased again due to the increase of organic loading and the subsequently accumulated toxic decolorization intermediates (Li et al., 2015). After several days of operation, MLSSs in all the three MBRs rose again.
The final MLSSs in 1#, 2#, and 3# MBRs were 3,650–3,720, 3,510–3,580, and 4,669–4,730 mg/L at the end of stage II, respectively. It was obvious that biomass concentration in all the three MBRs was lower than the initial MLSS of 5,000 mg/L after long-term operation, which suggested that some microbes were inhibited after being exposed to a new environment containing toxic compounds (Mrozik and Piotrowska-Seget, 2010). Among the three MBRs, the biomass concentration of 3# MBR was obviously higher and more stable than those of 1# and 2# MBRs throughout the whole operation processes. As discussed above, pure culture always displayed low adaptability to complex conditions during long-term operation of bioreactors. Although the MLSS of 2# MBR approached to that of 3# MBR during stage I, it sharply decreased at the beginning of stage II due to the increase of influent organic load.
On the other hand, the yeast G1 could detoxify ARB and its toxic decolorization intermediates, thus persevered some potentially useful microorganisms in AS from being inhibited (Li et al., 2015; He et al., 2017). By contrast, although ARB could be decolorized through reductive biodegradation by AS, most of the toxic intermediates (e.g., aromatic amines) could not be further degraded by AS. Thus, biomass concentration of 1# MBR obviously decreased during the long-term operation. In conclusion, extensive microorganisms in AS were probably preserved from being inhibited by toxic organics, which were detoxified by the yeast G1, thus resulting in higher MLSS of 3# MBR than the other two.
Toxicity assessment (the Microtox test)
To evaluate the detoxification effectiveness of three MBRs, acute toxicity of the influent and effluent at the end of stages I and II were analyzed through the Microtox test. The inhibition ratios (IRs) of about 91.5% and 94.7% shown in Fig. 3A and B suggested that the influents containing 200 mg/L and 300 mg/L ARB displayed high toxicity against V. fischeri. The IRs of about 94.5% and 96.7%, which corresponded to the effluents from 1# MBRs at the ends of stages I and II, respectively, suggest higher toxicity against V. fischeri than the corresponding influents. The possible reason might be that some more toxic breakdown intermediates of ARB were produced and accumulated (He et al., 2017).
By contrast, the effluents from 2# MBR at the ends of stages I and II showed moderate toxicity against V. fischeri, as indicated by the IRs of about 67.6% and 69.9%, respectively. Furthermore, the effluents from 3# MBR at the end of stages I and II showed much lower toxicity against V. fischeri than any of the others as indicated by the IRs of about 22.6% and 30.2%, respectively.
Many previous reports demonstrated that the coculture of fungi and bacteria displayed higher efficiency and stability for biodegradation of organic pollutants. For instance, it was previously found by many researchers that the cocultures of single fungal and bacterial strains showed higher azo dye decolorization efficiency than either of them (Gou et al., 2009; Qu et al., 2010; Su and Lin, 2013). Another study by Lade et al. (2012) indicated that the mixed community of a fungus Aspergillus ochraceus NCIM-1146 and a bacterium Pseudomonas sp. SUK1 showed obviously better performance for removal of COD, biological oxygen demand (BOD), and TOC of an actual textile effluent containing azo dyes than any of the single one, suggesting that the cocultures of fungus and bacterium might also display outstanding advantages for carbon mineralization in comparison with either of them.
Furthermore, this method had also been confirmed as a more effective and stable alternative in bioreactors (Li et al., 2015; He et al., 2017). Although many related studies had been done as mentioned above, few of them were performed under hypersaline conditions. In this study, the effectiveness of the coculture of AS and a halotolerant azo-degrading yeast G1 for treatment azo dye ARB under hypersaline conditions was estimated in MBR. The above results demonstrated that the yeast-augmented 3# MBR could efficiently decolorize high concentration (200–300 mg/L) of ARB and remove more than 88% of COD under hypersaline conditions (with 30 g/L NaCl). Furthermore, the result of toxicity assessment demonstrated that the yeast-augmented MBR also displayed high detoxification capability.
Dhouib et al. (2006) indicated that the AS system bioaugmented with P. chrysosporium or T. versicolor could obviously increase the BOD5/COD ratio of olive mill wastewaters, in contrast with the one inoculated with only AS. The yeast G1 had been confirmed that were capable of detoxifying ARB in the previous literature (Song et al., 2017). It could be speculated that the yeast G1 and other potentially effective fungi in 3# MBR played important roles in detoxification of ARB through further degrading the toxic decolorization intermediates into low toxic compounds, thus preventing other important microorganisms in AS from being inhibited.
Microbial community analysis by Miseq sequencing
Richness and diversity of bacterial and fungal phylotypes
Success colonization of selected microorganisms in bioaugmentation systems was significantly important for persistently high treatment efficiency and stability (He et al., 2017). Therefore, dynamics of microbial community was analyzed for monitoring the dominance of the yeast G1 or relative species belonging to the same genus in 3# MBR, as well as other dominant bacteria and fungi with potential capability. Relationship between bioreactor efficiency and microbial ecology was thereby expected to be understood.
After preliminarily removing short and low-quality reads, 69,808 and 45,451 sequences for bacterial and fungal communities were yielded from the Miseq sequencing analysis, respectively. The read lengths of 16S rRNA and 18S rRNA sequences, which correspond to bacterial and fungal communities, were about 466.13 and 283.46 bp, respectively. Although tens of thousands of tags per sample were determined, rarefaction curves (shown in Supplementary Fig. S2) were still far from the plateau, suggesting that there were some undetermined tags either from real rare species or artificial sequences produced by PCR and sequencing mistakes (Wang et al., 2012).
Richness and evenness of bacterial and fungal communities could be represented by the Shannon's diversity index (shown in Supplementary Table S1), respectively (Ma et al., 2013). The Shannon index corresponding to the bacterial community in original AS was 2.89. After continuously treating the simulated wastewater containing 200 mg/L (for 30 days) and 300 mg/L (for the following 30 days) ARB in 1# MBR, the Shannon indexes decreased to 2.60 and 2.26, respectively. It was suggested that richness and evenness of bacterial community in AS decreased due to exposure to high concentration of ARB and salt. Although only the yeast G1 was inoculated in 2# MBR at the beginning, multiple bacterial species were also detected at the end of stages I and II due to the nonsterile operating conditions.
In addition, richness and evenness of the bacterial communities in both 2# and 3# MBRs also decreased from 1.77 and 1.86 to 0.42 and 0.44 with the increase of influent ARB concentration from 200 to 300 mg/L, respectively. Comparatively, the Shannon's diversity index of the bacterial communities in both 2# and 3# MBRs was lower than that in 1# MBR. These results suggested that the richness and evenness of the bacterial community in all the three MBRs was inhibited by high ARB concentration; furthermore, inoculation of the yeast G1 further decreased the richness and evenness of bacterial community. On the other hand, the Chao1 index, which represented richness of microbial consortia, suggested that bacterial richness in all the three MBRs was similar in stage I.
When the influent ARB concentration increased to 300 mg/L in stage II, richness of the bacterial communities in 1#, 2#, and 3# MBRs increased from 475, 465, and 475 to 738, 603, and 556, respectively. It was obvious that richness of the bacterial community in 1# MBR at the end of stage II was the highest, followed by those in 2# and 3# MBRs. It was also suggested that, although the richness of bacteria in 2# and 3# MBRs was relatively high at the end of stage II, the corresponding evenness of bacteria was probably relatively low, thus resulting in much lower Shannon's diversity index. The Shannon index corresponding to the fungal community of original AS was 2.62, which also decreased to 1.85 and 1.59 at the ends of stages I and II, respectively.
Similarly, richness and evenness of the fungal communities in both of 2# and 3# MBRs decreased from 1.40 and 1.69 to 1.10 and 0.68 with the increase of influent ARB concentration, respectively. Furthermore, comparison of the Shannon's diversity index between different MBRs also indicated that fungal richness and evenness in 1# MBR was the highest; however, those in 3# MBR was the lowest.
On the other hand, the Chao1 index indicated that fungal richness in 2# MBR was the highest at the end of stage I, probably due to the initial inoculation of the yeast G1. However, the Chao1 index of the fungal community in 2# MBR decreased to lower than that in 3# MBR at the end of stage II, suggesting that the richness of fungal community in 2# MBR decreased after a long-term operation under nonsterile operation conditions; by contrast, those in 3# MBR were promoted due to the synergistic effect with other microorganisms in AS.
Taxonomic complexity of bacterial and fungal communities
Six dominant phyla or candidate divisions of bacteria were detected in all samples (results shown in Supplementary Fig. S3A). It was demonstrated that Proteobacteria (66.17%) was the dominant phylum in the original AS, followed by Bacteroidetes (19.04%) and Actinobacteria (1.52%). In the other six samples obtained from MBRs at the end of stages I and II, Proteobacteria was also one of the dominant phyla; however, Bacteroidetes almost disappeared. In 1# MBR, Firmicutes was another dominant phylum with the stable relative abundance of about 39.36%. In 2# and 3# MBRs, the relative abundance of phylum Firmicutes was 7.22% and 10.93% at the end of stage I; however, it decreased to 1.49% and 1.76% at the end of stage II, respectively.
As previously reported, bacteria belonging to Proteobacteria and Firmicutes were always detected from azo dyes treatment systems (Forss et al., 2013), which was consistent with the result of this study. Actinobacteria was also dominant in 1# MBR at the end of stage I with the relative abundance of 27.09%; however, only 0.64% was detected at the end of stage II. In addition, some bacteria belonging to unclassified phyla (30.19%) were dominant in 3# MBR at the end of stage I, and then they almost disappeared at the end of stage II. At genus level (as shown in Supplementary Fig. S3B), some bacteria were only dominant at the end of stage I, such as Humibacter and Alicyclobacillus in 1# MBR and Cryobacterium in 2# MBR.
When influent ARB concentration increased in stage II, the relative abundance of these genera sharply decreased, suggesting that they were inhibited. By contrast, the relative abundance of some other genera increased when exposed to higher organic loading, such as Leifsonia and Exiguobacterium in 1# MBR, and Metallibacterium in 2# and 3# MBRs. It was notable that genus Rhodanobacter, which was one of the original dominant bacterial genus in AS, became absolutely dominant in 2# and 3# MBRs after the long-term operation. This result suggested that Rhodanobacter might be related to the yeast G1. Many species belonging to the genus Rhodanobacter were found as efficient degraders of recalcitrant organics such as azoxystrobin, raw oil, and polycyclic aromatic hydrocarbons (Howell et al., 2014; Song et al., 2016; Xue et al., 2016).
In addition, some of them were identified as acid-tolerant bacteria (Koh et al., 2015; Dahal and Kim, 2017). In this study, the relative abundance of Rhodanobacter was higher in 2# and 3# MBRs, both of which were inoculated with the yeast G1. It was reported that the yeast G1 showed the optimal growing and metabolic performances under weak acidic conditions (Song et al., 2017). Therefore, the dominance of Rhodanobacter in 2# and 3# MBRs was possibly due to the synergistic degradation effects or the similar optimal pH conditions with the yeast G1.
On the other hand, the original AS was dominated by Ascomycota and Basidiomycota at the phylum level for fungal community. In the other six samples from three MBRs, Ascomycota was the absolutely dominant phylum, which was represented by the relative abundance of higher than 97.30% (shown in Supplementary Fig. S3C), which was also consistent with the results of related researches (Li et al., 2015; He et al., 2017). At the genus level (shown in Supplementary Fig. S3D), distribution and shifts of Candida and Pichia were notable. It was obvious that both Candida and Pichia were original dominant genera in AS with the relative abundance of 10.06% and 13.21%, respectively.
In 1# MBR, the relative abundance of Candida and Pichia increased to 29.64% and 19.75% at the end of stage I, and then decreased again to 14.73% and 13.18% at the end of stage II, suggesting that they might be inhibited by higher concentration of ARB under nonaugmented conditions. In contrast, the sum of the relative abundance of Candida and Pichia in 3# MBR exceeded 73.00% and 95.90% at the end of stages I and II, respectively. As reported, fungi belonging to Candida and Pichia were widely used for degradation of organic pollutants. For instance, Tan et al. (2013) and Qu et al. (2012) isolated fungal strains belonging to Candida and Pichia, respectively, both of which were capable of degrading azo dyes. Some of them were even identified as halotolerant ones in the studies reported by Jiang et al. (2015) and Song et al. (2017).
Furthermore, 44.9% and 81.3% were attributed by Pichia in 3# MBR at the end of stages I and II, respectively. This result suggested the possible colonization of the yeast G1 and/or some other potentially effective fungi affiliated to Pichia in 3# MBR during a long period of operation, thus enhancing the processes of ARB decolorization, COD removal, and detoxification.
Conclusions
Performance of AS-based MBR, which was bioaugmented by a halotolerant yeast P. occidentalis G1 for treating hypersaline ARB wastewater, was estimated through long-term operation. Dynamics of bacterial and fungal communities in MBRs were analyzed with MiSeq sequencing method. Compared with the MBRs inoculated with only AS or the yeast G1, the bioaugmented MBR showed higher treatment stability, efficiency, and tolerance to shock loading and sludge concentration during continuous treatment of hypersaline and high-concentration ARB wastewater.
Obvious differences were observed among the microbial communities from different MBRs during the long-term operation. Bacteria and fungi such as Rhodanobacter sp. and Candida sp. were enriched in the bioaugmented MBR. Furthermore, Pichia was identified as the most dominant fungal genus in 3# MBR, which suggested that the yeast G1 might colonize in the system, or some other potentially effective fungi affiliated to the same genus were enriched, thus enhancing the biodecolorization and detoxification efficiency.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The work was financially supported by the National Natural Science Foundation of China (Nos. 51878324, 51108223, and 51508259) and the General Project of Scientific Research of Liaoning Provincial Education Department (No. L201783644).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.