
Abstract
Biofilm carriers play an important role in attached growth systems for wastewater treatment processes. In this study, to establish a more robust and efficient anammox system, corncobs were used as biofilm carriers in the anammox process to tackle nitrite inhibition and enhance nitrogen removal. Results indicated that the nitrite- and nitrate-denitrifying activities of the corncob biofilm were 32% and 26%, respectively, higher than that of bulk sludge when adopting synthetic wastewater as influent that only contained NH4+-N and NO2−-N. The higher nitrite-denitrifying activity of the corncob biofilm had a positive effect on alleviation of nitrite inhibition. Furthermore, the corncob anammox system reached ∼92.5% total nitrogen (TN) removal stably, and the anammox pathway played a dominant role by carrying out more than 90% of TN removal. Anammox bacteria coexisted stably with the denitrifiers in the corncob biofilm, in which the anammox activity and denitrifying activity were 42.46 ± 0.58 mg NH4+-N d−1 and 10.28 ± 0.02 mg NO3−-N d−1, respectively. Therefore, employing corncobs as biofilm carriers was a useful and applicable supplement for enhancing the performance robustness of the anammox process.
Introduction
Ammonia nitrogen is one of the most widespread pollutants and nutrients in domestic sewage and industrial wastewater (Kumar and Lin 2010; Chen et al., 2017b). Anaerobic ammonium oxidation (anammox) technology is a promising alternative to the conventional nitrification–denitrification process for removing ammonia from sewage and wastewater (Mulder et al., 1995; Du et al., 2016; Kraiem et al., 2019), in which ammonium and nitrite are combined to generate nitrogen gas (Castrobarros et al., 2017; Shen et al., 2017). The anammox process can reduce the oxygen demand for nitrification and organic carbon consumption for denitrification (Ma et al., 2017; Wang et al., 2017a, 2018c).
However, some crucial factors influence the performance robustness of the anammox process. For instance, the inhibition effect of nitrite (both a substance and inhibitor) on anammox activity has been widely reported, but the optimum concentration for nitrite has not been widely accepted, which greatly threatens the performance robustness of the anammox process (Strous et al., 1999; Raudkivi et al., 2017). Furthermore, at the end of the anammox process, 11% of nitrate remains in the effluent, which lowers the nitrogen removal efficiency of anammox (Kumar and Lin 2010; Wang et al., 2017b). Therefore, promoting the efficiency and stability of the anammox process is necessary.
Anammox reactors with biofilm carriers or granular biomass have been reported to have higher IC50 values (concentration for inhibition of 50% activity of nitrite) than flocculated anammox reactors (Bettazzi et al., 2010; Raudkivi et al., 2017). Furthermore, granulation of anammox sludge is generally considered an effective method for enhancing the stability of the anammox process, while the flocculent anammox sludge usually requires a longer period to granulate due to the slow growth rate of anammox bacteria (Chen et al., 2017a). The biofilm technology, which can efficiently retain sufficient amounts of anammox bacteria in reactors, is another important means of improving the performance of the anammox process (Xie et al., 2017). However, one of the main drawbacks of biofilm technology is the requirement of ideal carriers for rapid anammox biofilm formation (Ding et al., 2018). Additionally, since nitrate is produced during the anammox process, it is also a crucial factor that would influence total nitrogen (TN) removal in the anammox system (Wang et al., 2015, 2017b).
Corncob is an essential, renewable agricultural residue and lignocellulosic crop worldwide (Xing et al., 2016; Menezes et al., 2017). It has been used as a biofilm carrier in bioreactors for denitrification and has been found to exhibit excellent performance for stable nitrogen removal (Li et al., 2012). Furthermore, it has been reported that the corncob mainly consists of cellulose, hemicellulose, and lignin (Yang et al., 2015; Cao et al., 2016b). Yang et al. (2015) demonstrated that the corncob possessed a relatively larger surface area, which is favorable for multiplication of bacteria with a low growth rate. Additionally, pore structures of corncobs provide more space for bacterial attachment and enrichment (Tseng and Tseng, 2005). In this study, to establish a more robust and efficient anammox reactor, a corncob was used as a biofilm carrier in the anammox process. However, the performance and stability of the corncob–anammox system remained unclear. Therefore, objectives of this study were to investigate the performance robustness of corncob-dosed anammox reactors, which use corncobs as biofilm carriers in the anammox process to tackle nitrite inhibition and enhance nitrogen removal by coupling anammox with denitrification. Furthermore, roles of corncobs and the microbial community structure in the reactor were discussed.
Materials and Methods
Reactor setup and operation
Four parallel sequencing batch reactors (SBRs) were set up for the anammox process with a working volume of 0.2 L, using corncobs as biofilm carriers. The corncobs were cut into small pieces (5–10 mm diameter) and dried for use. Next, different weights (10.0, 5, 2.5, and 0 g/L) of corncobs were prepared for the experimental reactors R1, R2, R3, and the blank reactor R0, respectively. Before initiation of these reactors, corncobs were immersed in the SBRs for 1 week to stabilize the releasing rate of carbon source, expressed as soluble chemical oxygen demand (COD). Subsequently, the four SBRs were inoculated with anammox sludge, which was collected from an anammox reactor in the laboratory (stable operation over 2 years) (Wang et al., 2018a). The mixed liquor volatile suspended solid (MLVSS) of SBRs, after inoculation, was 2.60 g VSS/L. Operating parameters of the four reactors are shown in Table 1. The hydraulic retention time of the four reactors was 24 h during the 60 days of operation, corresponding to the nitrogen loading rate of 0.232 kg N/(m3·d) in phase 1 and 0.116 kg N/(m3·d) in phase 2, respectively. The four SBRs were completely covered with a black polyvinyl chloride (PVC) material (3-mm thickness) to prevent light penetration (Du et al., 2016).
Operating Parameters of the Reactors
Inf., influent.
Synthetic wastewater
NH4Cl and NaNO2 were added into the feed synthetic wastewater with influent NO2−-N/NH4+-N ratio of 1.32. NH4+-N and NO2−-N concentrations were about 100 and 132 mg/L in phase 1 and decreased to 50 and 66 mg/L in phase 2, respectively. The composition of the medium solution was (per liter) 10 mg KH2PO4, 500 mg KHCO3, 300 mg MgSO4·7H2O, 6.4 mg CaCl2·2H2O, and 1.0 mL trace element solution. The trace element solution contained (per liter) 15 g EDTA, 18 g FeSO4·7H2O, 0.43 g ZnSO4·7H2O, 0.25 g CuSO4·5H2O, 0.19 g NiCl ·6H2O, 0.24 g CoCl ·6H2O, 0.014 g H3BO4, 0.99 g MnCl2·4H2O, 0.22 g NaMoO4·2H2O, and 0.21 g NaSeO4·10H2O (Van de Graaf et al., 1996).
Bacterial activity tests
To investigate the activity of anammox and denitrifying bacteria, in-site batch tests were carried out on day 6 (phase 1) and day 60 (phase 2). In batch test A, 50 mg/L NH4+-N and 66 mg/L NO2−-N were added into the reactors (R1, R2, R3, and R0) to determine the activity of anammox bacteria. In batch test B, 66 mg/L NO2−-N was added to measure the specific nitrite reduction rate. In batch test C, 15 mg/L NO3−-N was added into the reactors to assay the specific nitrate reduction rate. All tests were conducted in a shake incubator at a temperature of 35°C and a speed of 150 rpm. The batch tests were run for 300 min and about 1 mL of supernatants was taken out every 60 min (Ma et al., 2017). Furthermore, the activity of bulk sludge and corncob biofilm in R1 was determined to clarify the role of corncob carriers.
Sampling and analytical methods
Liquid samples were collected, on a daily basis, from the influent and effluent of the reactors. Samples were filtered through 0.45-μm filters and used for analyses of NH4+-N, NO2−-N, NO3−-N, and COD, and sludge samples were collected to measure the mixed liquor suspended solids (MLSSs) and MLVSSs according to standard methods (APHA, 2005).
Fluorescence excitation–emission matrix measurements
Fluorescence excitation–emission matrix (EEM) spectra were measured using a luminescence spectrometer (Hitachi, Tokyo, Japan) to contrast the variation of dissolved organic matter of wastewater samples. The emission wavelengths (Em) and excitation wavelengths (Ex) of EEM were increased incrementally from 200 to 600 nm at 5-nm intervals with a scan speed of 1,200 nm/min (Ly et al., 2018; Yang et al., 2018).
Calculations
The contributions of anammox and denitrification pathways to TN removal were evaluated based on depletion of NH4+-N, NO2−-N, and NO3−-N. The concentration of TN was calculated by summing concentrations of NH4+-N, NO2−-N, and NO3−-N. Nitrogen loss due to microorganism assimilation was neglected (Du et al., 2017).
Percentage of anammox contribution to TN removal
Anammox percentage = [(Inf.NH4+-N − Eff.NH4+-N) +1.32 (Inf.NH4+-N − Eff.NH4+-N) −0.26 (Inf.NH4+-N − Eff.NH4+-N)] × 100%/(Inf.TN − Eff.TN) where Inf.NH4+-N and Eff.NH4+-N are the influent and effluent concentrations of NH4+-N during operation, respectively, and Inf.TN and Eff.TN are TN concentrations of the influent and effluent, respectively (Chen et al., 2016).
Percentage of denitrification contribution to TN removal
Denitrification percentage = [(0.26 (Inf.NH4+-N − Eff.NH4+-N) − Eff.NO3−-N) + (Inf.NO2−-N − Eff.NO2−-N) −1.32 (Inf.NH4+-N − Eff.NH4+-N)] × 100%/(Inf.TN − Eff.TN)
where Eff.NO3−-N, Inf.NO2−-N, and Eff.NO2−-N are the effluent concentration of NO3−-N and influent and effluent concentrations of NO2−-N during operation, respectively (Du et al., 2017).
Microbial community analyses
The inoculated anammox sludge and sludge samples (bulk sludge and biofilm) collected from R1 on day 60 were used for analysis of the microbial community through the Illumina MiSeq platform (Illumina, Inc., San Diego, CA). The DNA from sludge samples was extracted using the extraction kit (E.Z.N.A™ Mag-Bind Soil DNA Kit; OMEGA) following the manufacturer's instructions. Bacterial 16S rDNA genes were PCR amplified with primers 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) for the V4 region (Wang et al., 2018a). All analyses were conducted by Sangon Biotech Co., Ltd. (Sangon, China) (Wang et al., 2018b).
Results and Discussion
Performance recovery of the anammox process after nitrite inhibition
In phase 1 (1–6 day), the reactors containing corncobs (10.0, 5.0, and 2.5 g/L) were operated as experimental groups (R1, R2, and R3) and the one without corncobs was operated as the control group (R0), with the influent NH4+-N and NO2−-N concentrations of 100 and 132 mg/L, respectively. Results showed that the effect of nitrite inhibition on the anammox performance in the control reactor (R0) was significant. On day 6, the removal efficiencies of NH4+-N, NO2−-N, and TN in R0 rapidly declined to 31.0%, 55.4%, and 38.4%, respectively (Fig. 1a, b, d). Furthermore, the inhibitory effect was also observed in the experimental groups (R1, R2, and R3) with respect to decrease in nitrogen removal. However, the effect of inhibition was lower in the experimental reactors than in the control reactor (Fig. 1). Subsequently, the influent concentrations of NH4+-N and NO2−-N were decreased to 50 and 66 mg/L, respectively, in phase 2 (7–65 days). The performance recovery of the control reactor (R0) was very slow and the TN removal efficiency only reached 78% by day 46. In contrast, the experimental groups (R1, R2, and R3) showed a faster performance recovery rate after nitrite inhibition, especially reactors R1 and R2 (Fig. 1). It is worth mentioning that the maximum dosage of corncobs used (R1, 10 g/L) greatly shortened the performance recovery time. On day 12, the removal efficiency of TN of R1 had increased to around 85%, and it was observed that R1 had better stability and higher nitrogen removal efficiency compared with other reactors (Fig. 1d). During the stable period in R1, the removal efficiency of TN was found to be at around 92.5% and the residual nitrate in the effluent was much lower than in the other reactors (Fig. 1c). Evidently, corncobs provided organics for denitrification in bulk sludge and carrier biofilm, which accelerated the recovery of anammox activity after nitrite inhibition and further improved TN removal in the reactor.
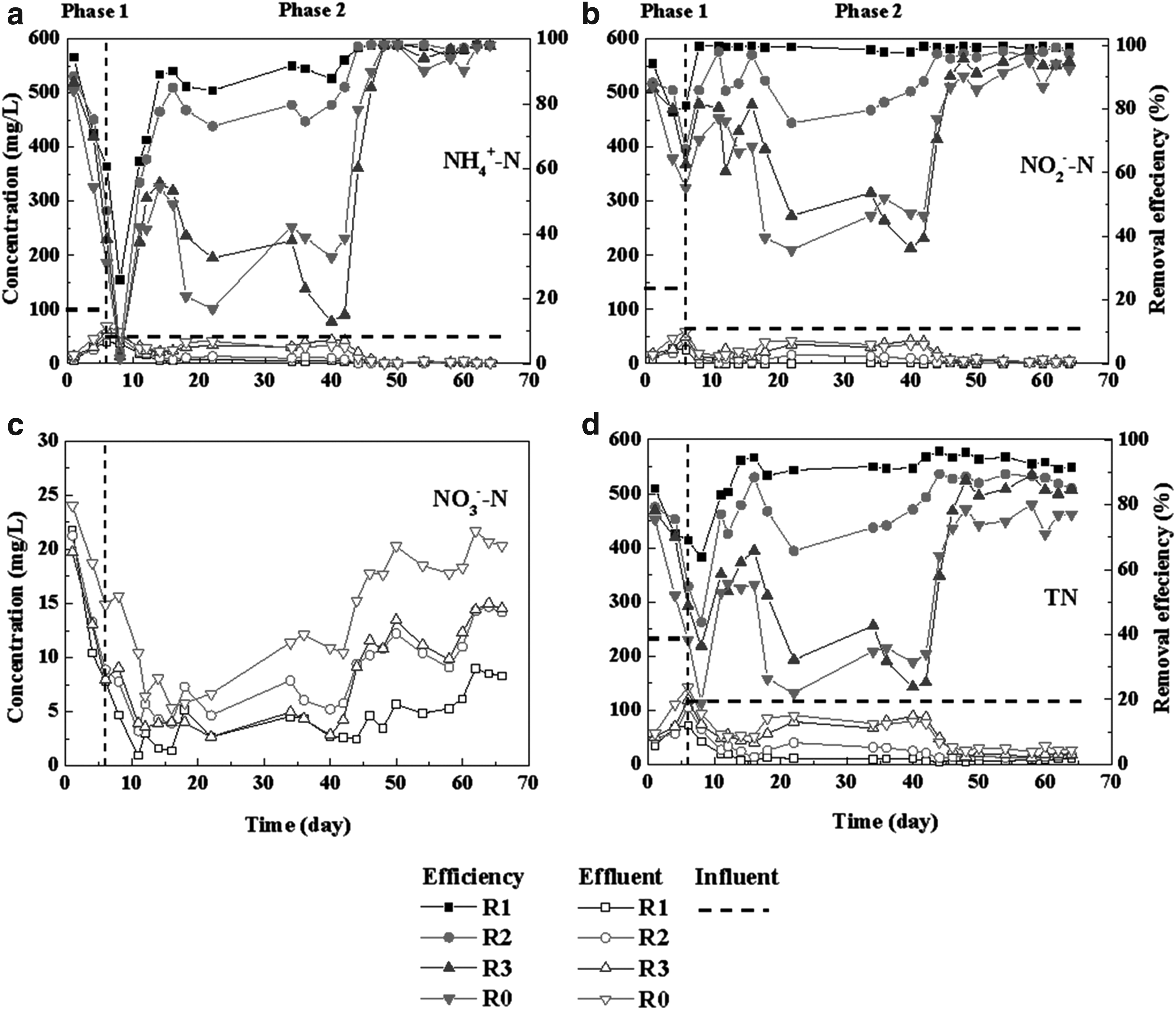
Nitrogen removal performance of the reactors.
Previously, Xu et al. (2009) reported that corncobs could be used as an effective and economical carbon source for denitrification. Li et al. (2012) also suggested that the corncob was a feasible carbon source and biofilm carrier for denitrifying microorganisms. Corncobs are rich in xylan and cellulose among other agricultural wastes (Yang et al. 2005), and consequentially could release the bioavailable carbon source sustainably in lixivium. The COD concentration, after the corncob was soaked in reactors, decreased and stabilized on day 6 in the soaking stage, which was similar to that reported by Yang et al. (2015). The EEM analysis results of R1 also indicated that organic matter released from the corncob was decomposed by bacteria in the reactor (Fig. 2). Subsequently, anammox sludge was inoculated into the reactors, and the COD concentrations in the effluent of R1, R2, and R3 were 26 ± 3, 23 ± 2, and 20 ± 2 mg/L, respectively, during the experimental stage (Fig. 3). Although there was an increasing trend of carbon source release with increase in corncob dosage, the organic matter in corncob biocarriers was released slowly and did not accumulate in the effluent.
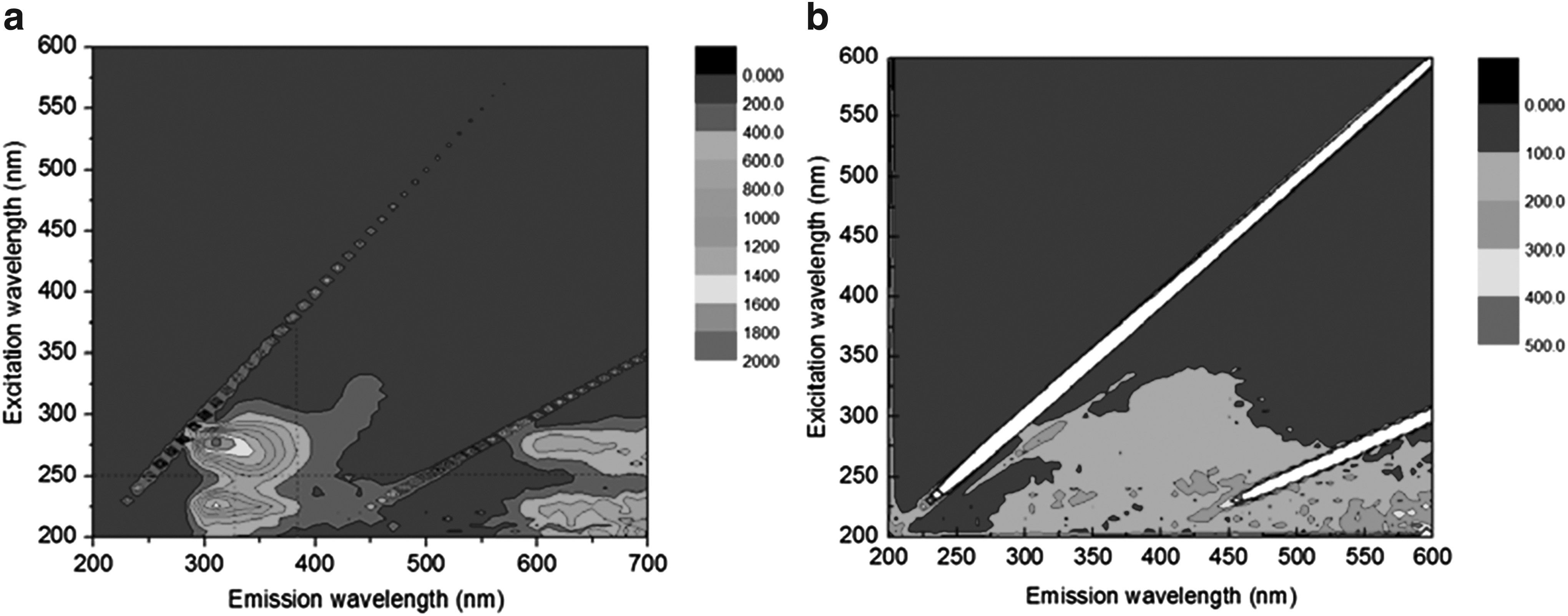
EEM fluorescence spectra of dissolved organic matter in the effluent of R1
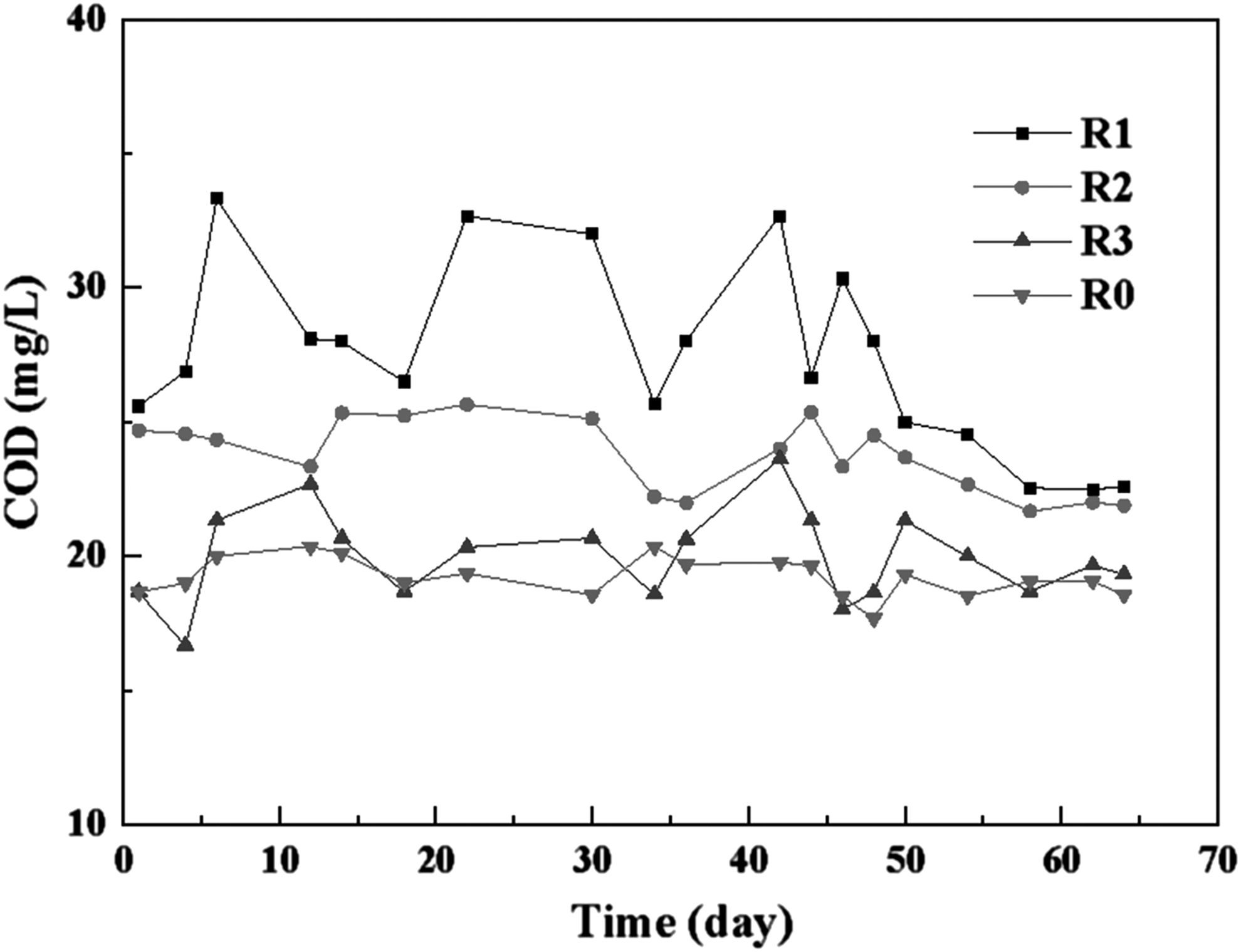
Variation of COD concentration in the leaching solution. COD, chemical oxygen demand.
The stable and efficient performance of R1 was mainly ascribed to the contribution of denitrification and anammox pathway to TN removal (Fig. 4). During phase 1, the percentage contribution of the anammox pathway to TN removal decreased to about 36% due to inhibition of nitrite, while the percentage of denitrification gradually increased to around 64%. Subsequently, the anammox pathway occupied a dominant position with contribution to TN removal having increased to above 90% and contribution of denitrification maintained at below 10% during phase 2. Although anammox bacteria were inhibited by nitrite, the anammox pathway remained predominant in the R1 system. It indicated that successful cooperation between denitrification and anammox had been achieved at the COD/TN ratio of around 0.2, and denitrification played a key role in alleviating nitrite inhibition in phase 1, which greatly accelerated anammox recovery. Lotti et al. (2012) found that the anammox activity inhibited by NO2−-N could be fully recovered after removal of nitrite. Furthermore, coordination of denitrification and anammox enhanced the TN removal performance at a COD/TN ratio of 0.8 (Ding et al., 2018). Therefore, autotrophic anammox bacteria could steadily coexist with heterotrophic denitrification in the corncob–anammox system, which was similar to the result that anammox bacteria and denitrifiers could share the same environment (Chen et al., 2016).
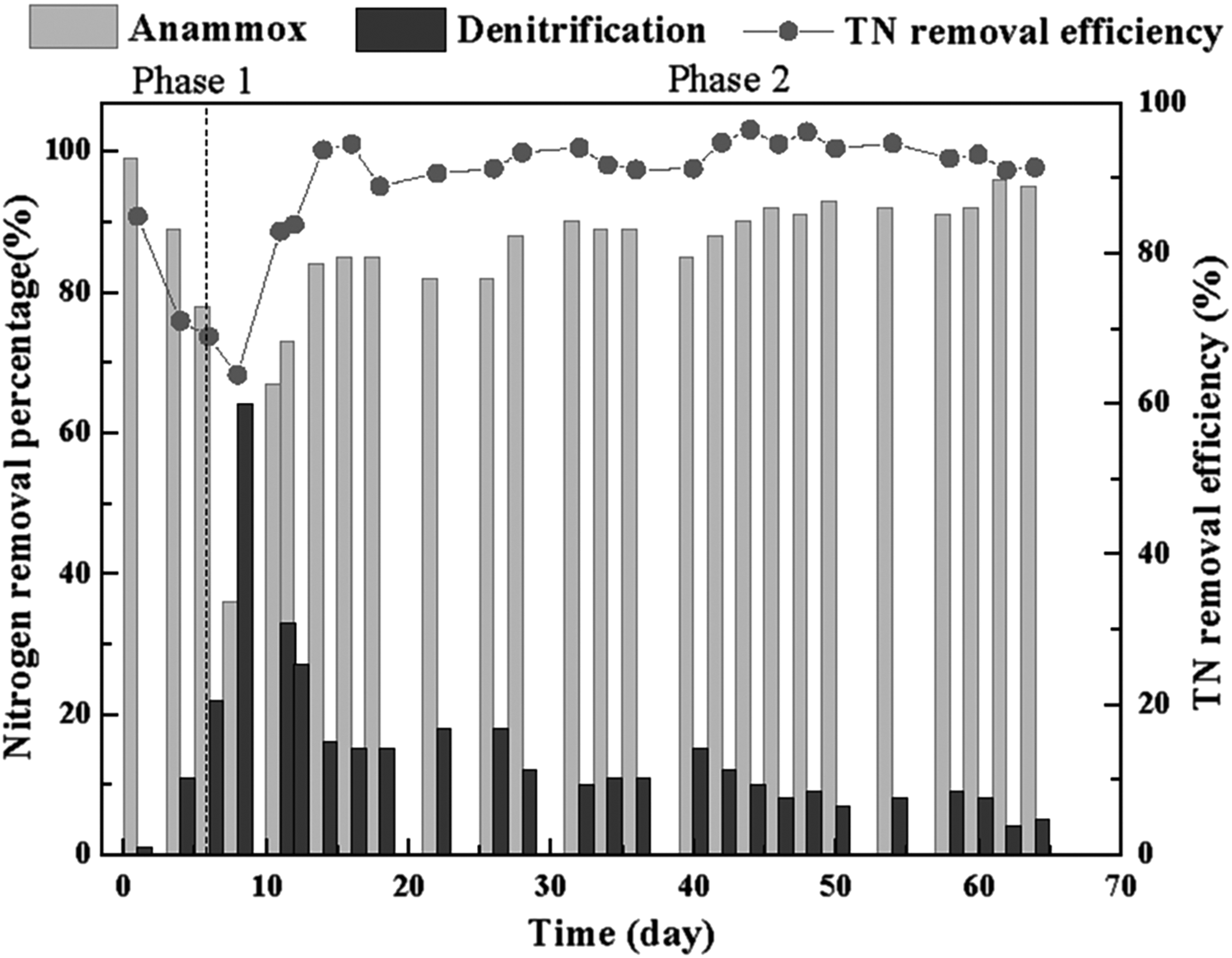
The relative contribution to TN removal by anammox and the denitrification pathway and the TN removal efficiency.
As observed in Fig. 4, nitrogen removal by anammox was always maintained as the dominant pathway during phase 2, which guaranteed efficient and stable nitrogen removal performance, suggesting that using corncobs as biocarriers was a serviceable and controllable supplement to the anammox process.
Analysis of anammox activity and denitrifying activity of corncob biofilm and bulk sludge
Successful coupling between denitrification and anammox in corncob-dosed reactors led to high-efficiency nitrogen removal, which could be testified by the anammox and denitrification activity assays (Fig. 5). The specific anammox activity (SAA) of the inoculum was 189.96 ± 0.36 mg NH4+-N d−1. In phase 1, after exposure to nitrite at the concentration of 132 mg/L, the activity of anammox was inhibited significantly. In the control reactor (R0), the SAA decreased to 22.62 ± 0.26 mg NH4+-N d−1 on day 6. Meanwhile, in the experimental reactors (R1, R2, and R3), the SAA of sludge decreased to 83.43 ± 0.28, 42.56 ± 0.22, and 35.78 ± 0.30 mg NH4+-N d−1, respectively (Fig. 5A). Hence, addition of corncobs had a positive effect on reduction of nitrite inhibition, which mitigated the damage to anammox microorganisms due to high concentration of nitrite. As revealed by the nitrite-denitrifying activity tests (test B), the reactors containing corncobs during phase 1 showed a higher consumption rate of nitrite than those in phase 2 (Fig. 5B). Subsequently, the SAA of anammox bacteria recovered to 157.53 ± 0.34, 136.94 ± 0.22, 124.85 ± 0.49, and 112.03 ± 0.41 mg NH4+-N d−1 in R1, R2, R3, and R0, respectively, on day 60. Therefore, addition of corncobs (R1, R2, and R3) contributed to a faster performance recovery rate and higher anammox activity compared with the control reactor (R0) (as shown in Figs. 1 and 5A).
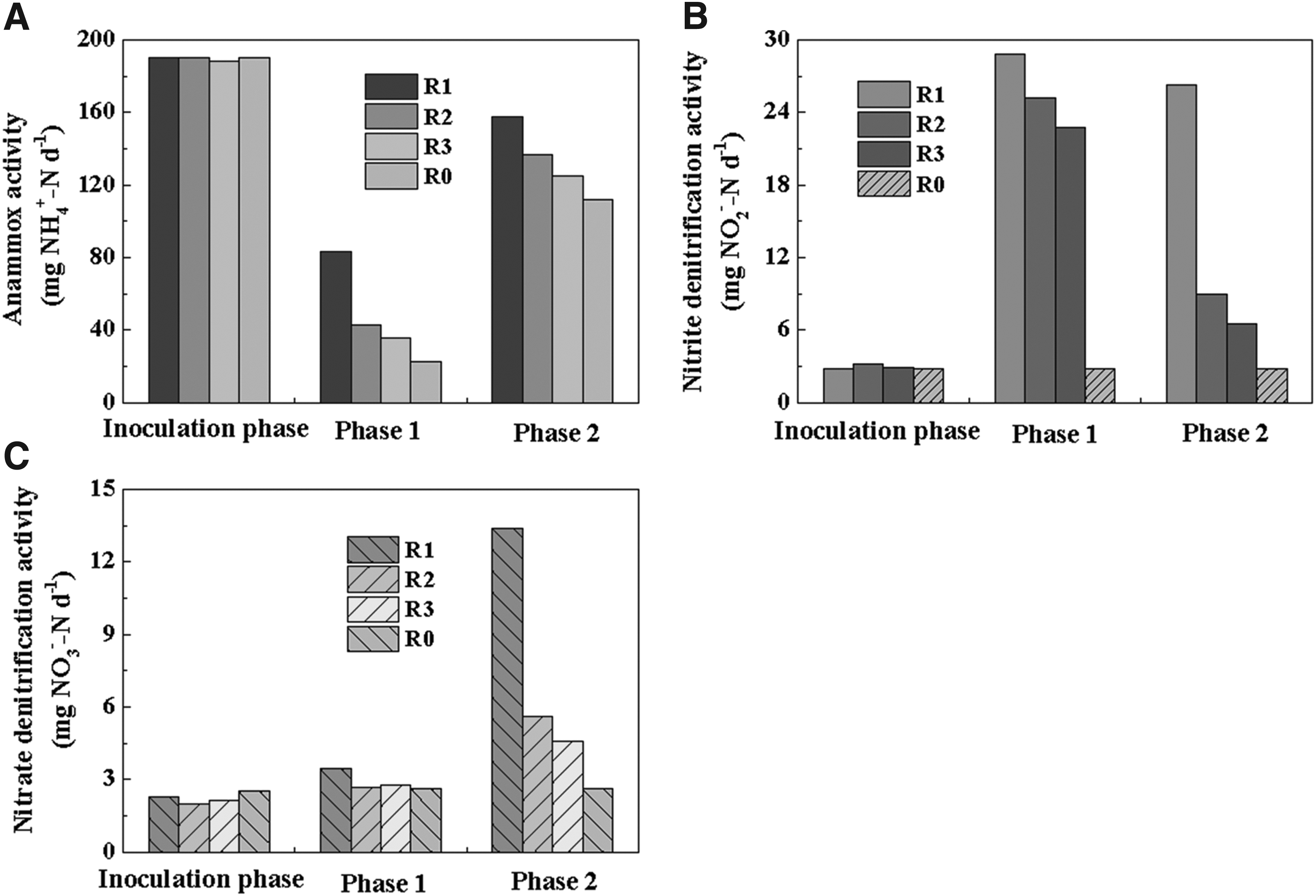
Variations of bacterial activity.
In addition, the nitrite-denitrifying activity decreased with the increase of anammox activity. This was probably due to the low COD concentration in lixivium of the corncob–anammox reactors (Fig. 3), which limited the nitrite-denitrifying activity. Thus, consumption of NO2−-N through anammox occupied a dominant position during phase 2. As demonstrated by Xu et al. (2009), the relative proportion of soluble components in the corncob decreased with time, which explained the phenomenon described above. Zheng et al. (2017) also reported that anammox bacteria could perform with higher competitive ability of nitrite compared with denitrifiers in the low COD/NO2−-N ratio condition. As shown in Fig. 5C, the reactors containing corncobs had higher nitrate-denitrifying activity during phase 2 than phase 1. The probable reason was that more nitrate was produced during phase 2 in the anammox process when anammox activity was gradually recovered, which provided more electron acceptors for denitrification. However, the anammox pathway still occupied the dominant position in corncob-dosed reactors.
By analyzing the SAA and denitrifying activity of the corncob biofilm and bulk sludge in R1, higher nitrite- and nitrate-denitrifying activities were observed in the corncob biofilm (Supplementary Table S1). The nitrite- and nitrate-denitrifying activities of the corncob biofilm were 20.20 ± 0.08 mg NO2−-N d−1 and 10.28 ± 0.02 mg NO3−-N d−1, respectively, which were 32% and 26% higher than that of bulk sludge. In the meantime, the SAA of bulk sludge and corncob biofilm was 115.10 ± 0.64 and 42.46 ± 0.58 mg NH4+-N d−1, respectively (Table 2). Therefore, the corncob was a suitable biofilm carrier that could make anammox bacteria and denitrifiers work cooperatively in the system.
Comparison of Anammox and Denitrification Activity of the Bulk Sludge and Corncob Biofilm
Microbial community analysis
High-throughput sequencing analysis was used to investigate the microbial community structure of R1. The Shannon index of the inoculum was 3.87, lower than that of the bulk sludge (3.94) and corncob biofilm (4.33) in the anammox–corncob system (Table 3). This indicated that the corncob biofilm had comparatively higher bacterial diversity than the bulk sludge as well as inoculum.
Richness and Diversity Estimators of Microbial Communities in the R1 Reactor
The microbial community structure was analyzed at the phylum level, as shown in Fig. 6a. In the inoculum, 20 phyla were detected, which was lesser than that observed in the corncob-dosed system (26 phyla). In the corncob-dosed system, the dominant phyla in the bulk sludge were Proteobacteria (25.63%), Firmicutes (15.45%), Armatimonadetes (12.53%), Ignavibacteriae (9.99%), Chloroflexi (7.45%), Planctomycetes (6.71%), and Bacteroidetes (5.98%), and the dominant phyla in the corncob biofilm were Proteobacteria (42.82%), Firmicutes (19.81%), Planctomycetes (7.62%), Chloroflexi (7.46%), and Bacteroidetes (3.12%). The phyla, Proteobacteria and Chloroflexi, were responsible for autotrophic and heterotrophic denitrification (Wen et al., 2016; Ding et al., 2018). The relative abundance of these two phyla in the corncob biofilm (50.28%) was higher than that in bulk sludge (33.08%), suggesting that the corncob biofilm might contain a higher percentage of denitrifiers than bulk sludge, which corresponded to the denitrification activity tests (Table 2). Another phylum, Bacteroidetes, in connection with filamentous bacteria, has the property of degrading organic carbon (Guo and Zhang, 2012; Xu et al., 2017). There was a high abundance of phylum Bacteroidetes found in the corncob-dosed system, which suggested that Bacteroidetes might play a key role in degrading organic matter released from the corncob. Last, the phylum Planctomycetes, which is related to anammox bacteria, was detected simultaneously in the corncob biofilm and bulk sludge. The relative abundance of the phylum Planctomycetes in the corncob biofilm (7.62%) was higher than that in the bulk sludge (6.71%), which illustrated that anammox bacteria could be enriched in the corncob biofilm.

The relative abundance of bacterial communities in R1 at different taxonomic classifications of
The dominant bacterial species at the genus level are shown in Fig. 6b. The genera Candidatus Brocadia and Candidatus Kuenenia belonging to Planctomycetes were detected as anammox bacteria (van der Star et al., 2007; Cao et al., 2016a), both of which existed in the corncob biofilm and bulk sludge. Relative abundance of these two anammox bacteria in the biofilm (6.68%) was higher than that in bulk sludge (5.06%), indicating that the corncob was a suitable biofilm carrier for enrichment of anammox bacteria. Thiobacillus, Pseudomonas, and Thauera were the functional bacteria detected in the corncob-dosed system, which related to denitrification (Kelly and Wood, 2000; Peix et al., 2009; Du et al., 2017). There was low abundance of Pseudomonas in the inoculum (0.1%) without Thiobacillus and Thauera. Nevertheless, the corncob biofilm contained a higher percentage of denitrifiers (4.03%), higher than that in the bulk sludge (0.3%) as well, which was consistent with denitrification activity tests (Table 2). Notably, the corncob biofilm contained a higher percentage of anammox bacteria and denitrifiers than bulk sludge in the corncob-dosed system. Meanwhile, the abundance of anammox bacteria in the corncob biofilm was higher than that in denitrifiers. The result was similar to that Wen et al. (2017) found the abundance of Thauera (2.3%) and Pseudomonas (0.6%), which were dominant denitrifying bacteria in the simultaneous partial nitrification, anammox and denitrification (SNAD) process, was much lower than anammox bacteria (52.%), where the COD/TN ratio varied at 0.2–1.0. Furthermore, the sludge concentration in reactors remained stable during phases 1 and 2. The MLVSS measurements were 2.60 ± 0.06, 2.58 ± 0.12, and 2.56 ± 0.04 g VSS/L of the inoculum and bulk sludge in the reactors at the end of phases 1 and 2, respectively. Therefore, the anammox biofilm formed in a short time considering the transfer of anammox bacteria from bulk sludge. Taken together, corncobs were applicable biofilm carriers for anammox microorganisms and denitrifiers to coexist, which successfully achieved the synergistic effect for nitrogen removal in the coupling system.
Conclusions
Corncobs were successfully used as biofilm carriers in anammox reactors to tackle inhibition of nitrite and enhance nitrogen removal. Corncobs provided organics for denitrification in the bulk sludge and carrier biofilm, which enabled anammox bacteria and denitrifiers to work cooperatively in the reactor. High nitrite-denitrifying activity of the corncob biofilm significantly accelerated the performance recovery rate of the anammox process after nitrite inhibition. The abundance of anammox bacteria in the corncob biofilm was higher than that of denitrifiers. Furthermore, the synergistic effect of anammox and denitrification was established, which enhanced performance robustness and nitrogen removal in the corncob-dosed system.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was funded by the National Natural Science Foundation of China (51878232), the Major Science and Technology Research Project in Anhui Province (2017YF-05), and the Foundation of Anhui Shun Yu Water Co., Ltd. (W2018JSKF0137).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.