
Abstract
The phytoremediation technology is widely used due to its beautiful appearance and no secondary pollution. The mechanism of how the plants cope with the abiotic stresses imposed by polyaromatic hydrocarbon and other organic pollutants is not fully explored. Our previous study showed that the expression of MjHSP18.2 (small heat shock protein 18.2) and MjHSP17.7 (small heat shock protein 17.7) was remarkably upregulated in Mirabilis jalapa L. under petroleum stress through the predifferential proteome analysis. To improve our understanding of small heat shock proteins (sHSPs), we have characterized MjHSP18.2 and MjHSP17.7 from M. jalapa. Sequence alignments and phylogenetic analysis reveal these to be a cytosolic class I sHSPs. MjHSP18.2 and MjHSP17.7 expression was induced by petroleum stress in M. jalapa. Recombinants MjHSP18.2 and MjHSP17.7 were overexpressed in yeast Saccharomyces cerevisiae to study its possible function under stress conditions. The recombinant yeast cells that accumulated MjHSP18.2 and MjHSP17.7 conferred tolerance to tunicamycin compared with control cultures. Under tunicamycin-induced endoplasmic reticulum (ER) stress conditions, MjHSP18.2 and MjHSP17.7 transgenic yeast cells were found to depress the expression of ER molecular chaperones, increase cell viability, decrease cell apoptosis, and increase superoxide dismutase activities compared with control cultures. These results suggest that MjHSP18.2 and MjHSP17.7 could alleviate the ER stress caused by tunicamycin and promote cell survival in yeast cells.
Introduction
Small heat shock proteins (sHSPs) belong to the HSP superfamily with a molecular mass range of 12–42 kDa (Wang et al., 2004; Sun et al., 2012) and are characterized by the presence of a conserved α-crystallin domain (ACD) (Franck et al., 2004; Basha et al., 2012). According to their homology and intracellular location, the plant sHSP family can be classified into six groups: cytosolic class I, cytosolic class II, endoplasmic reticulum (ER), mitochondria, chloroplast, and peroxisome sHSPs, respectively (Löw et al., 2000; Ma et al., 2006; Mamedov and Shono, 2008; Siddique et al., 2008). The sHSPs are molecular chaperones that can either aid in the folding and maintenance of newly translated proteins, or they can bind to misfolded and destabilized proteins to prevent irreversible molecular aggregation or correct protein misfolding (Basha et al., 2012; Hilton et al., 2013; Haslbeck and Vierling, 2015).
sHSPs are ubiquitous in both eukaryotes and prokaryotes (Waters and Rioflorido, 2007; Aevermann and Waters, 2008; Waters, 2013). Most of the sHSPs are not expressed at normal conditions, but can be rapidly induced by various abiotic stress conditions, including salt, cold, drought, and plant hormone (Sun et al., 2002; Sun and Macrae, 2005; Park et al., 2013; Yang et al., 2014). Homologues of some sHSPs, such as in maize (Cao et al., 2010), tobacco (Hamilton and Coleman, 2001), Arabidopsis (Dafnyyelin et al., 2008), cotton (Maqbool et al., 2010), tomato (Frank et al., 2009), sorghum bicolor (Nagaraju et al., 2019), and Rosa (Jiang et al., 2009), have proven that they play a role in the protection of cells in the face of adverse effects, from harm caused by unfavorable factors. These results suggest that sHSPs may play an important role in plant response to general abiotic stress.
The ER is the site for protein synthesis, folding, modification, and transport. A variety of molecular chaperones and foldases are involved in this system (Halperin et al., 2014), such as the binding immunoglobulin protein (BiP) and protein disulfide isomerase (PDI) (Gething, 1999; Noiva, 1999). Also, the accumulation of unfolded proteins in the ER lumen will consequently lead to ER stress. To ensure the faithfulness of protein folding and prevent the occurrence of aggregation between proteins, cells will make a corresponding response, which is called unfolded protein response (UPR), to relieve the ER protein overload (Schröder and Kaufman, 2005; Wan and Jiang, 2015). In plants, abiotic stress can disturb the protein-folding process and result in ER stress under natural conditions. The ER stress is related to the signal pathway of environmental stress responses, such as signaling of reactive oxygen species (ROS) (Liu et al., 2007; Liu and Howell, 2010). In recent years, studies have shown that sHSPs can regulate intracellular redox state and control cell ROS content, thereby enhancing cell tolerance to oxidative stress and improving cell viability under oxidative stress (Prabhu et al., 2011). Overexpressed LeHSP21 protein of tomato in tobacco improved tolerance to high-temperature stress by enhancing antioxidation capacity (Neta-Sharir and Weiss, 2005; Zhang et al., 2016). As UPR not only induces survival adaptation but also induces apoptosis signal, cell autophagy was thought to be the main mechanism to counteract UPR in plant (Pu and Bassham, 2013). sHSP was able to directly regulate cell apoptosis by participating in the signal transduction pathway of apoptosis death (Deegan et al., 2013; Sano and Reed, 2013).
Due to the rapid development of the economy and increased pace of petroleum extraction, the resulting problem of pollution is increasingly significant. Petroleum contamination is one of the most serious kinds of soil pollution (Jha et al., 2008), which results in harmful effects, such as germinant inhibition of seeds, reduced photosynthesis, slowed nutrient assimilation, and shortened root growth (Smith et al., 2006; Rahbar et al., 2012; Redondo-Gómez et al., 2014). In recent years, phytoremediation was regarded as an emerging cleanup technology for contaminated soils (Gopalakrishnan et al., 2007; Chen et al., 2017). Thus, plants that have high tolerance to petroleum are desirable for bioremediation. Mirabilis jalapa, which is one of the most important ornamental plants, usually is selected as the suitable remediation plant in petroleum-contaminated soils because of its large biomass and strong adaptability (Peng et al., 2009). However, few reports demonstrating the role of sHSP genes in M. jalapa have been published to date. Thus, research to reveal the molecular mechanism of petroleum stress tolerance is of interest.
In our previous work, we found that the expression of two sHSPs (MjHSP18.2 and MjHSP17.7) was remarkably upregulated in M. jalapa under oil stress through the predifferential proteome analysis. In this study, the relationship between sHSP and adaptive UPR was observed by detecting the expression of transcription factors BiP and PDI in transgenic yeast with overexpressing sHSP and the control group. Simultaneously, we measured the cell apoptosis rate, cell survival rate, superoxide dismutase (SOD) activities of transgenic yeast, and the control group. The results of this study is to provide some new insights into the molecular mechanisms of sHSPs under abiotic stresses induced by polyaromatic hydrocarbon and other organic pollutants in M. jalapa.
Materials and Methods
Plant materials, culture conditions, and stress treatments
In this study, M. jalapa seedlings were used as plant material. M. jalapa seedlings were grown on clean soils for 30 days in the greenhouse at Shenyang Agricultural University, Shenyang, China. Then seedlings were transferred into petroleum-contaminated soil (treated with 0, 5, 10, 20, 30, 40 g/kg petroleum for contaminated treatment) for 12, 24, and 36 h, respectively. The main components of petroleum were 72.17% of saturated hydrocarbon fraction, 17.82% of aromatic hydrocarbon fraction, and 10.01% of asphaltene and polar fraction. To ensure the soil homogeneity, the contaminated soils (1 kg clean soils with 40 g petroleum) were sieved through a 4 mm sieve and fully mixed. Through the addition with different clean soils, contaminated soils were diluted according to the experimental design. In other words, after the dilution, the concentration (Woil/Wsoil) of petroleum in the tested soils was 5, 10, 20, 30, and 40 g/kg, respectively (Peng et al., 2009; Agnello et al., 2016). The root was sampled and washed with distilled water. After drying by filter paper, the root was immediately frozen in liquid N2 and stored at −80°C for further analysis.
Total RNA extraction and single-strand complementary DNA synthesis
Total RNA was extracted from tissues with the TRIzol reagent (Invitrogen), according to the manufacturer's instructions. Then the quantity of the total RNA was confirmed by the 260/280 nm ratio using NanoDrop 2000 (Thermo Scientific). Also, to verify the quality of total RNA, we loaded the total RNA in a 1% agarose gel with ethidium bromide (EB) and checked the 18/28S ribosomal RNA integrity. The first single-strand complementary DNA (cDNA) was synthesized from total RNA using the PrimeScript™ RT Reagent Kit (TaKaRa). The synthesized cDNAs were finally stored at −80°C.
Cloning of MjHSP18.2 and MjHSP17.7 genes from M. jalapa
Our previous comparative proteomic data show that the expression of MjHSP18.2 and MjHSP17.7 was remarkably upregulated compared with the control group under the stress of petroleum. The full-length sequence was determined using 5′ and 3′ Rapid Amplification of cDNA Ends (RACE) with commercial kits (Invitrogen), according to the manufacturer's instructions. According to the partial peptide sequence of MjHSP18.2 and MjHSP17.7 from M. jalapa, we designed gene-specific primers using the Primer Express 3.0 software (PE Applied Biosystems) based on the conserved sequences of MjHSP18.2 and MjHSP17.7 from closely related species. The gene-specific primer sequences used for MjHSP18.2 and MjHSP17.7 amplification are given in Table 1. For 3′ end RACE PCR, the cDNA template was obtained using Adapter Primer (Invitrogen), and PCR was conducted with primers HSP18.2 3F and HSP17.7 3F and an adapter primer abridged universal amplification primer (Invitrogen). The PCR conditions were as follows: 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 60°C for 45 s, 72°C for 90 s, and finally, 72°C for 10 min, hold at 4°C. PCR products were cloned into a pMD-19T Cloning Kit (TaKaRa) and plasmids were purified using the MinElute gel extraction kit (Qiagen) and sequenced. For the 5′ end RACE PCR, the RNA was transcribed by a 5′ RACE kit (Invitrogen) with gene-specific primers HSP18.2 5R and HSP17.7 5R, and the cDNA was subsequently amplified with an adapter primer AAP (RACE kit; Invitrogen), and gene-specific primers HSP18.2 6R and HSP17.7 6R. The PCR conditions were one cycle of 94°C for 5 min, followed by 30 cycles at 94°C for 30 s, 53°C for 45 s, 72°C for 90 s, and finally, 72°C for 10 min. PCR products were cloned into a pMD-19T Cloning Kit (TaKaRa) and plasmids were purified using the MinElute gel extraction kit (Qiagen) and sequenced. All of the sequencing is done by the Shanghai Sangon Company.
The Gene-Specific Primer Sequences Used for MjHSP18.2 and MjHSP17.7 Amplification from Mirabilis jalapa Are Listed
F, forward; HSP, heat shock protein; R, reverse.
Construction and transformation of three yeast recombinant vectors
The MjHSP18.2 (with Flag tags) and MjHSP17.7 (with HA tags) open reading frame (ORF) was amplified from the abovementioned PCR products using two pairs of gene-specific primers. Meanwhile, we also obtained MjHSP18.2 and MjHSP17.7 fusion proteins through the means of fusion PCR with 2A self-cleaving linker peptide from Thosea asigna virus (T) sequence (Szymczak-Workman et al., 2012), which could simultaneously regularly express these two kind of proteins MjHSP18.2 and MjHSP17.7 in a single expression vector. The gene-specific primer sequences used for obtaining MjHSP18.2-Flag, MjHSP17.7-HA, and fusion gene of MjHSP18.2 and MjHSP17.7 are given in Table 2. The PCR-amplified products were digested by BamHI–XbaI and directionally cloned into the vector pYES2, designed for inducible expression of recombinant proteins in Saccharomyces cerevisiae, and the resulting plasmids were named pYES2-HSP18.2, pYES2-HSP17.7, and pYES2-HSP-fusion, respectively. Sequence analysis confirmed that the pYES2-HSP18.2, pYES2-HSP17.7, and pYES2-HSP-fusion genes were successfully inserted into the pYES2 vector. Then, the plasmid constructs were transformed into the S. cerevisiae strain INVSC1 with the Frozen-EZ Yeast Transformation II Kit™, according to the manufacturer's instructions. Yeast cells containing pYES2-HSP18.2, pYES2-HSP17.7, pYES2-HSP-fusion, and the control pYES2 vector were cultured in SC Dropout Medium without uracil Yeast Synthetic Drop-out Media (SC-URA) liquid medium containing 2% glucose at 30°C overnight. The resulting cultures were transferred to the SC-URA medium containing 2% (w/v) galactose and incubated at 30°C for 1 day to promote expression of MjHSP18.2, MjHSP17.7, and fusion gene of MjHSP18.2 and MjHSP17.7.
The Gene-Specific Primer Sequences Used for Obtaining HSP18.2-Flag, HSP17.7-HA, and Fusion Gene of MjHSP18.2 and MjHSP17.7 from Mirabilis jalapa Are Listed
Real-time PCR quantification
To elucidate the role that MjHSP18.2 and MjHSP17.7 played in ER stress, transgene yeast was treated with 10 μg/mL tunicamycin for 0, 4, 8, 12, or 10 h to simulate ER stress. The expression of ER-stress-related genes was measured by real-time reverse transition-polymerase chain reaction (RT-PCR) using CFX96™ (Bio-Rad, USA). The RealMaster SYBR Green PCR Mixture contained 8.5 μL RNase-free ddH2O, 12.5 μL 2 × Taq PCR Master Mix, 1 μL 5′ primer (10 μM), 1 μL 3′ primer (10 μM), and 2 μL cDNA template (equivalent to 100 ng of total RNA). The reaction conditions were as follows: 94°C for 5 min, followed by 40 cycles at 94°C for 30 s, 60°C for 45 s, 72°C for 1 min, and 72°C for 10 min. The fluorescence signal was collected during the elongation at 72°C of every cycle. Each sample was run in duplicate wells, and each experiment was carried out in three independent replicates. The fold change in the relative gene expression was calculated according to the 2−ΔΔCT formula (Livak and Schmittgen, 2001). All gene-specific and internal control primer sequences used for real-time RT-PCR are shown in Table 3.
All Gene-Specific and Internal Control Primers Sequences Used for Real-Time RT-PCR are Listed
Western blot analysis
Proteins were extracted by homogenizing induced yeast cells in extraction buffer [15 mL 1 mol Tris-HCl (pH 8.0), 25 mL glycerol, 2 g insoluble polyvinyl pyrrolidon (PVP), water up to 100 mL], according to the CelLytic™ Y Plus Kit (Sigma). Protein samples were separated on 15% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to the polyvinylidene fluoride membranes (Bio-Rad) for antibody probing. Primary antibodies targeting the MjHSP18.2, MjHSP17.7, and fusion proteins of MjHSP18.2 and MjHSP17.7 were purchased from the Abmart Company. Goat anti-mouse IgG (dilution at 1:2000) was used as a secondary antibody. SDS-PAGE and Western blotting were performed as described previously (Sambrook et al., 2001).
Sequence alignment and phylogenetic analysis
The amino acid sequence of MjHSP18.2 and MjHSP17.7, along with other representative plant cytosolic class I sHSP genes, was constructed with ClustalX (www.clustal.org/clustal2). The sequence alignment was performed to generate a phylogenetic tree with bootstrapping (1000 replicates), following the neighbor-joining method provided by the software MEGA5 (Tamura et al., 2011).
Cell growth, apoptosis, and survival assays
For the stress treatment (Gao et al., 2011), after the initial OD600 reached 0.8–1.0, 10 μg/mL tunicamycin was added into cultures. The samples were centrifuged and washed once with 1 × phosphate-buffered saline. Cultures grown in yeast extract peptone dextrose (YEPD) medium were diluted 10-fold in five series, and samples from each diluent (5 mL) were spotted onto SC-URA medium containing 2% (w/v) galactose and incubated, and then incubated at 28°C for 2–3 days.
For survival assessment, the 3-(4,5)-dimethylthiahiazo (-z-y1)-3,5-di-phenytetrazoliumromide (MTT) was conducted according to the manufacturer's instructions (Sigma) in a 96-well microculture plate format. Cells were seeded at 106 cells and plates were incubated in a humidified incubator in 5% carbon dioxide (CO2) for 24 h at 37°C. To each well was added 10 μL MTT (10 mg/mL) and incubated in a humidified incubator in 5% CO2 for 24 h. After centrifugation for 10 min, the supernatant of the plate was sucked out, and then adding 200 μL dimethyl sulfoxide to dissolve precipitate. Plates were read at an absorbance of 490 nm with Microplate Reader. Cell apoptosis was evaluated by the Dead Cell Apoptosis kit with Annexin V Alexa Fluor™ and propidium iodide according to instruction (Vermes et al., 1995).
Measurement of SOD activities
Yeast strain INVSC1 and the transgenic yeast cells were treated with 10 μg/mL tunicamycin for 0, 4, 8, 12, and 16 h to measure SOD activity. Each sample (1 g) was homogenized in 5 mL of 0.05 molL−1 potassium phosphate buffer, pH 7.0, containing 1 mM ethylene diamine tetraacetic acid and 1% PVP. The homogenate was centrifuged at 30,000 × g for 30 min at 4°C, and the upper aqueous layer containing the enzyme was immediately used for detection of enzyme activities. SOD activities were assayed by the nitro-blue tetrazolium method as described by Beyer Jr. and Fridovich (1987). The total SOD activities were then measured at an absorbance of 560 nm with a spectrophotometer at 25°C. Each experiment was carried out three times at least.
Statistical analysis
The data analyses were performed using the statistical software SPSS (version 19.0). Data are expressed as mean ± standard deviation. The significance of the correlations was performed using one-way analysis of variance followed by Duncan's multiple range test. A p < 0.05 was considered to indicate statistical significance.
Results
MjHSP18.2 and MjHSP17.7 as the members of plant cytosolic class I sHSP gene family
Two full-length cDNAs containing 483 and 474 bp ORF were isolated from M. jalapa, respectively, and showed significant homology to other cytosolic class I sHSP genes from various species by basic local alignment search tool (BLAST). One was predicted to encode a protein of 161 amino acid residues with a molecular mass of 18.2 kDa. The other was predicted to encode a protein of 158 amino acid residues with a molecular mass of 17.7 kDa. Based on the predicted molecular weights, the identified genes were designated as MjHSP18.2 and MjHSP17.7 (GenBank accession number), respectively. The deduced amino acid sequences of MjHSP18.2 and MjHSP17.7 were aligned with other representative cytosolic class I sHSPs and demonstrated significant homology to sHSPs of other plants (Fig. 1). The deduced amino acid sequences of MjHSP18.2 and MjHSP17.7 aligned with other seven representative cytosolic class I sHSPs revealed that both of them contain an ACD, which includes two conservative regions [Pro-X(14)-Gly-Val-Leu; Pro-X(14)-Val/Leu/Ile-Val/Leu/Ile] of sHSPs using BLAST (Fig. 1).
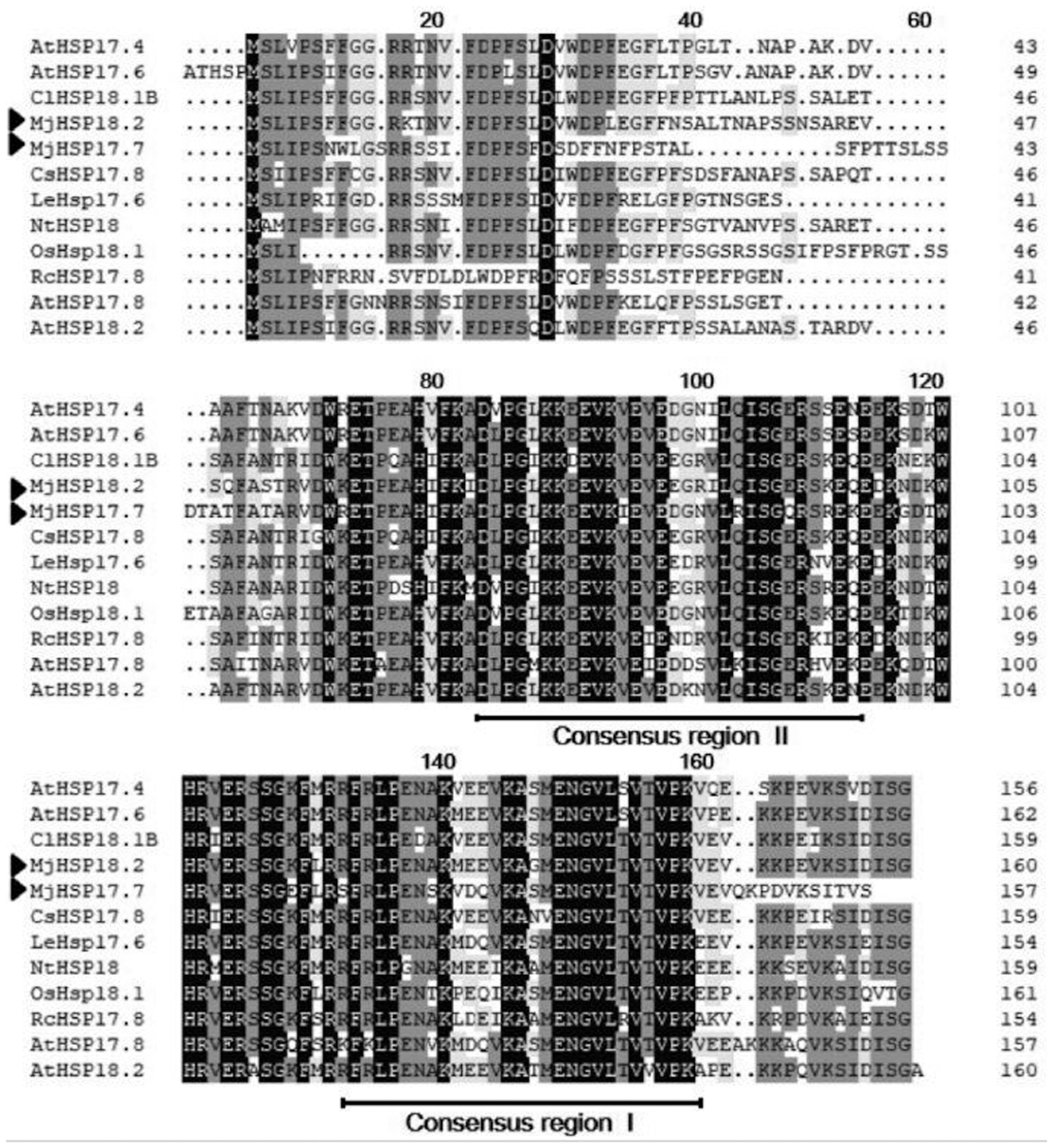
Alignment of the deduced amino acid sequences of MjHSP18.2 and MjHSP17.7 with other plant sHSPs. The two conserved regions in the sHSPs are boxed and identical amino acid residues are highlighted. Their GenBank accession numbers are as follows: Mirabilis jalapa (MjHSP17.7); Mirabilis jalapa (MjHSP18.2); Arabidopsis thaliana (AtHSP17.4, NM_114492); (AtHSP17.6, NM_121240); (AtHSP17.8, NM_100614); (AtHSP18.2, NM_125364); Rosa chinensis (RcHSP17.8, EF053229); Nicotiana tabacum (NtHSP18, X70688); Lycopersicon esculentum (LeHSP17.6, LEU72396); Cucumis sativus (CsHSP17.8, XP_004149942); Oryza sativa (OsHSP18, FJ383169); and Citrullus lanatus (ClHSP18.1B, ADU55794). MjHSP18.2 and MjHSP17.7 are labeled with the solid triangle at the left. HSP, heat shock protein; sHSPs, small heat shock proteins.
The evolutionary relationships of MjHSP18.2 and MjHSP17.7 with other sHSPs were also analyzed. The phylogenetic tree (Fig. 2) analysis indicated that two MjHsps were classified into cytosolic class I, which was consistent with the alignment. To further confirm that MjHSP18.2 and MjHSP17.7 belong to the cytosolic sHSP family, the amino acid sequence of MjHSP18.2 and MjHSP17.7 was analyzed in the ProtComp v.9.0 database. MjHSP18.2 and MjHSP17.7 were predicted to be localized to the cytosol. Taken together, the results suggested that MjHSP18.2 and MjHSP17.7 are a member of the plant cytosolic class I sHSP gene family.
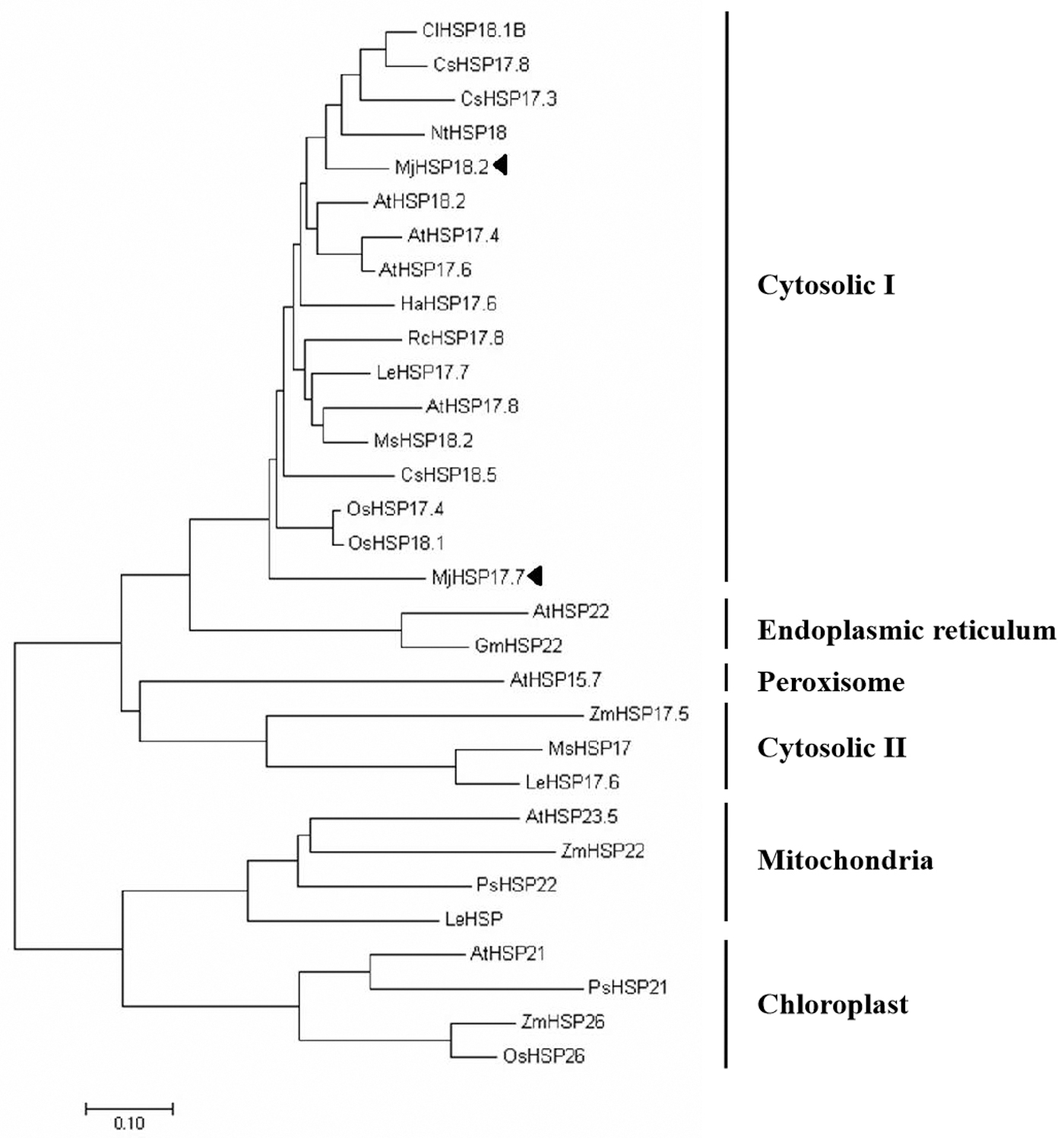
Phylogenetic tree of the deduced amino acid sequences of MjHSP18.2 and MjHSP17.7 and other sHSPs. The accession numbers for all sequences and the abbreviations of the species are as follows: Mirabilis jalapa (MjHSP17.7); Mirabilis jalapa (MjHSP18.2); Citrullus lanatus (ClHSP18.1B, ADU55794); CsHSP17.3, XP_004150234); (CsHSP18.5, XP_004153730); Arabidopsis thaliana (AtHSP17.4, NM_114492); (AtHSP17.6, NM_121240); (AtHSP17.8, NM_100614); (AtHSP18.2, NM_125364); (AtHSP21, NM_118906); (AtHSP23.5, NM_124523); (AtHSP15.7, DQ403190); (AtHSP18.2, NM_125364); Nicotiana tabacum (NtHSP18, X70688); Helianthus annuus (HaHSP17.6, X59701); Rosa chinensis (RcHSP17.8, EF053229); Lycopersicon esculentum (LeHSP17.6, LEU72396); (LeHSP17.7, AF123255); (LeHSP, AB017134); Medicago sativa (MsHSP18.2, X58711); (MsHSP17, X98617); Oryza sativa (OsHsp18.1, Q84Q72); (OsHsp17.4, P31673); Glycine max (GmHSP22, X63198); Zea mays (ZmHSP17.5, EU970990); (ZmHSP26, L28712); ZmHSP22, AY758275); (ZmHSP26, L28712); (ZmHSP22, AY758275); Pisum sativum (PsHSP22, X86222); (PsHSP21, X07187). MjHSP18.2 and MjHSP17.7 are labeled with the solid triangle.
Expression of MjHSP18.2 and MjHSP17.7 in M. jalapa induced by petroleum stress
In our previous work, two small heat shock protein (sHSP) genes have been identified, which were remarkably upregulated in M. jalapa under oil stress through the predifferential proteome analysis (Chen et al., 2017). To investigate whether the two sHSPs were associated with petroleum-contaminated soil restoration, we examined the expression of MjHSP18.2 and MjHSP17.7 under petroleum treatment. As shown in Fig. 3, when subjected to 5, 10, 20, and 30 g/kg of petroleum 12 h after treatment, MjHSP18.2 was significant increased, reaching a peak at 30 g/kg and then decreased. There was no change in the MjHSP18.2 transcription level under 40 g/kg treatment conditions. As shown in Fig. 3, when subjected to 5, 10, 20, and 30 g/kg of petroleum 12, 24, and 36 h, MjHSP17.7 expression presented upgrade first, and then, the descending later tendency, and reached a maximum at 24 h of treatment with 20 g/kg petroleum at various levels of petroleum stresses (5, 10, 20, 30 g/kg). Similarly, MjHSP18.2 expression had no significant changes under 40 g/kg treatment.
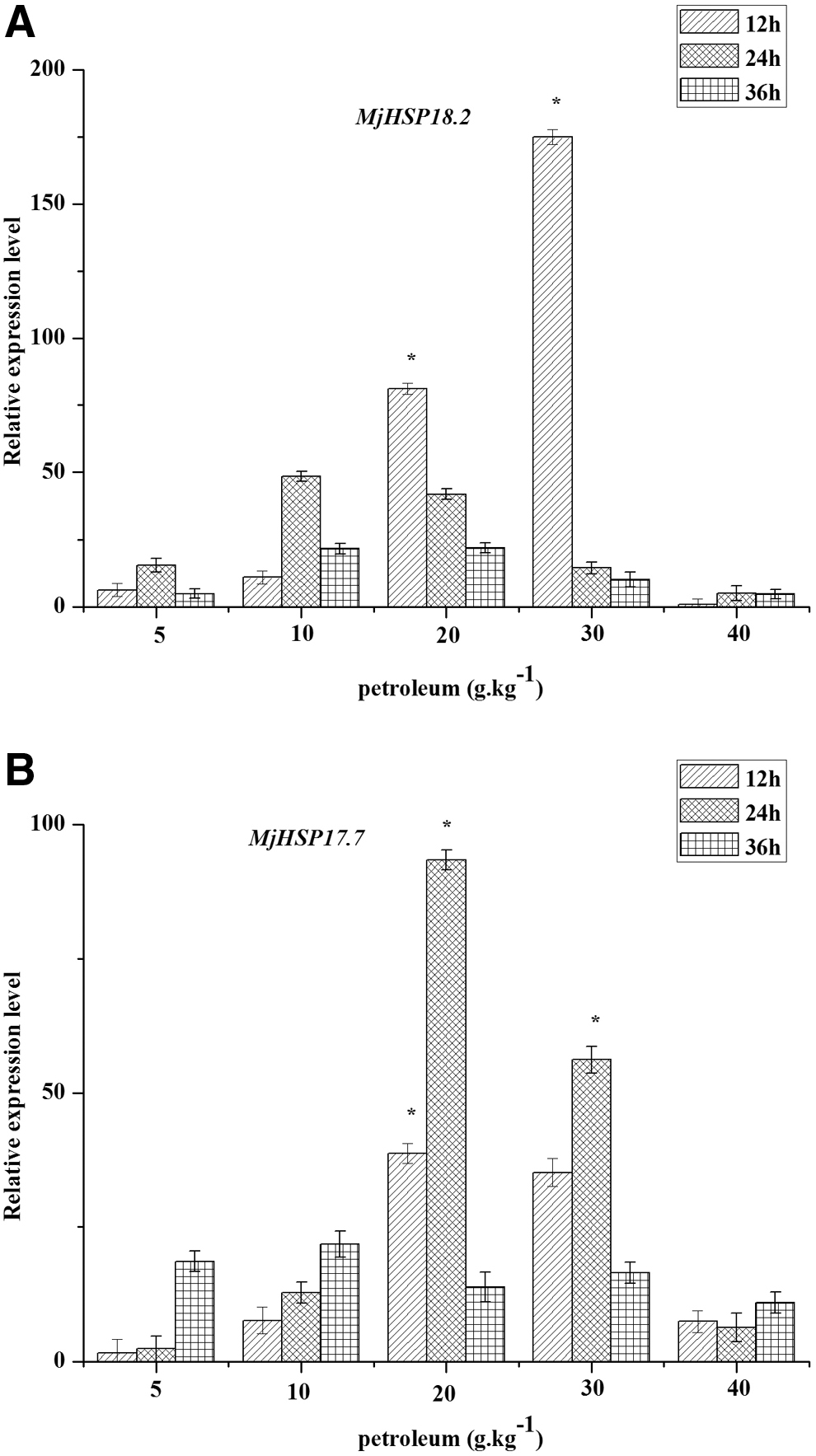
Expression analysis of MjHSP18.2
Tunicamycin-resistant ability of transgenic S. cerevisiae
To examine the sensibility of the introduced MjHSP18.2 and MjHSP17.7 on tunicamycin, four types of transformed yeast, which were pYES2-MjHSP18.2, pYES2-MjHSP18.2, pYES2-MjHSP-fusion, and an empty pYES2 plasmid as a control, were generated. Western blot analysis confirmed that all of them were expressed in yeast (data not shown). To test this, cultures of empty vector and vectors carrying pYES2-MjHSP18.2, pYES2-MjHSP17.7, and pYES2-MjHSP-fusion strains were streaked on YEPD medium plates with 15 ug/ml tunicamycin treatment, and then observed the growth of these cultures. As shown in Fig. 4, the growth of Saccharomyces cells expressing MjHSP18.2, MjHSP17.7, and MjHSP-fusion was better than cells compared with controls under the same tunicamycin stress. The results showed that MjHSP18.2 and MjHSP17.7 could enhance the tolerance to tunicamycin stress of Saccharomyces cells.
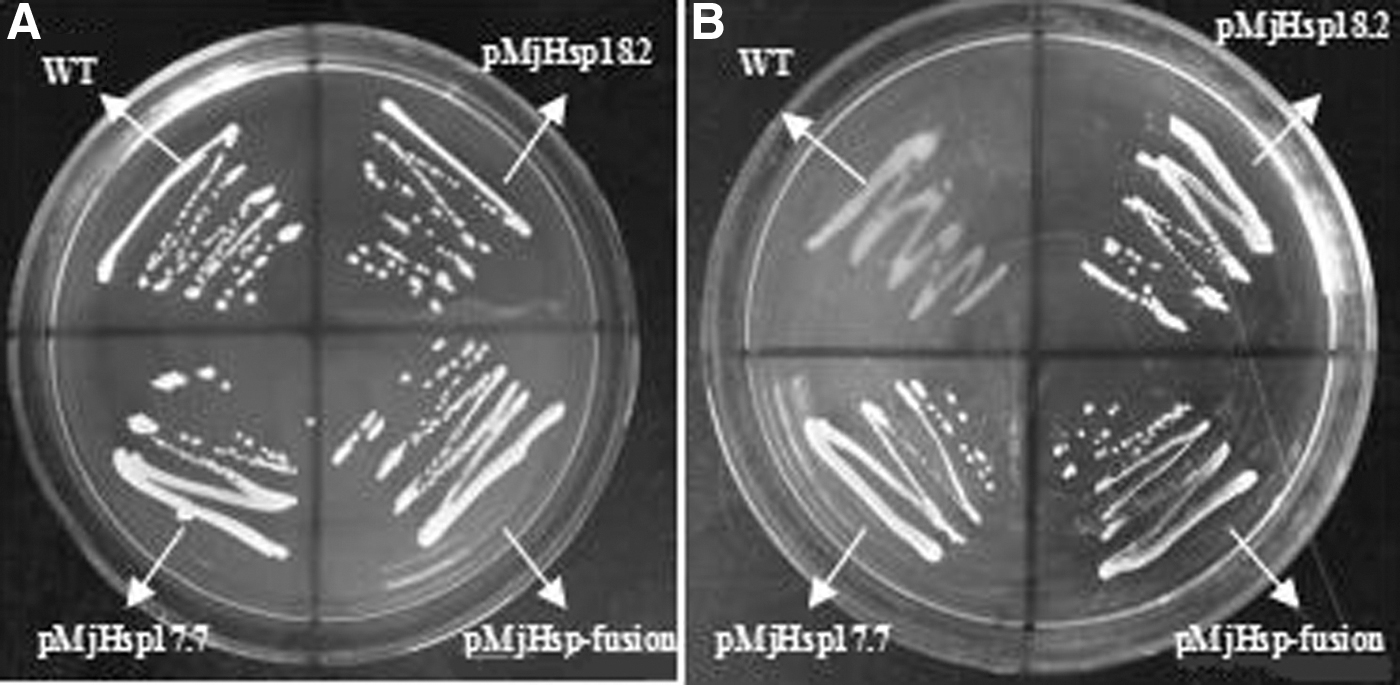
Tunicamycin resistance ability analysis of transgenic yeast cells and the control yeast strain INVSC1.
Tunicamycin induced the expression of ER-stress-related genes
Tunicamycin is an inducer of ER stress. To show the occurrence of the ER stress caused by tunicamycin and the activation of UPR at the transcript level, the expression of ER-stress-related genes was measured. So, we investigated the effect of the introduced MjHSP18.2 and MjHSP17.7 on the UPR under tunicamycin stress. The tunicamycin-induced expression of BiP and PDI1 rapidly increased in both transgenic and nontransgenic yeast cells, but this increase was less evident in the transgenic yeast cells compared with the nontransgenic yeast cells (Fig. 5). These results indicated that overexpressing MjHSP18.2 and MjHSP17.7 can alleviate the ER stress caused by the tunicamycin stress in yeast cells, and may participate in the UPR signal transduction pathway.

Expression of UPR genes under tunicamycin stress. Relative expression of
Tunicamycin-induced apoptosis and Yac1 expression
Excessive or sustained ER can cause cell death or apoptosis. To clarify the effect of ER stress on the apoptosis, we measured the apoptosis and cell survival rate of Saccharomyces cells. Our data showed that Annexin V-positive characteristics were more prone to occur in Saccharomyces cells at 8 h after tunicamycin treatment (data not shown). Only 10.04% of the yeast cells that were not treated with tunicamycin showed Annexin V positive. However, 70.87% of the yeast cells, which were treated with tunicamycin but not genetically modified, showed Annexin V positive. The apoptosis rate of MjHSP18.2 and MjHSP17.7 transgenic group was lower than that in the nontransgenic group under the same tunicamycin treatment (Fig. 6A). Our data showed that tunicamycin treatment resulted in a significant increase of Yac1 messenger RNA (mRNA) expression (Fig. 6B). Otherwise, the cell survival rate of the unmodified yeast cells was significantly lower than that of the yeast cells that were transferred to MjHSP18.2, MjHSP17.7, and MjHSP-fusion (Fig. 6C). These results suggest that MjHSP18.2 and MjHSP17.7 play a role in reducing apoptosis and increasing cell viability in the condition of ER stress.

ER stress induced the SOD activities
Under the condition of ER stress, the formation of protein disulfide bond in ER also produces oxidative stress. SOD has been suggested to be an important enzyme in protecting cells against oxidative stress. To test whether MjHSP18.2 and MjHSP17.7 regulate the activities of antioxidant enzymes, SOD was measured in response to ER stress. Yeast strain INVSC1 and the transgenic yeast cells were treated with tunicamycin for 0, 4, 8, 12, and 16 h to measure SOD activity. As shown in Fig. 7, SOD activity showed a trend from rising to decline and reached the highest point after 8 h, which indicated that it was the critical value of the peroxidation of the yeast membrane. Compared with nontransgenic yeast cells, SOD activities were significantly higher in the transgenic yeast cells in response to tunicamycin stress. The results showed that transgenic yeast cells had strong antioxidant capacities, and MjHSP18.2 and MjHSP17.7 enhanced the activity of S. cerevisiae in the stress of ER.
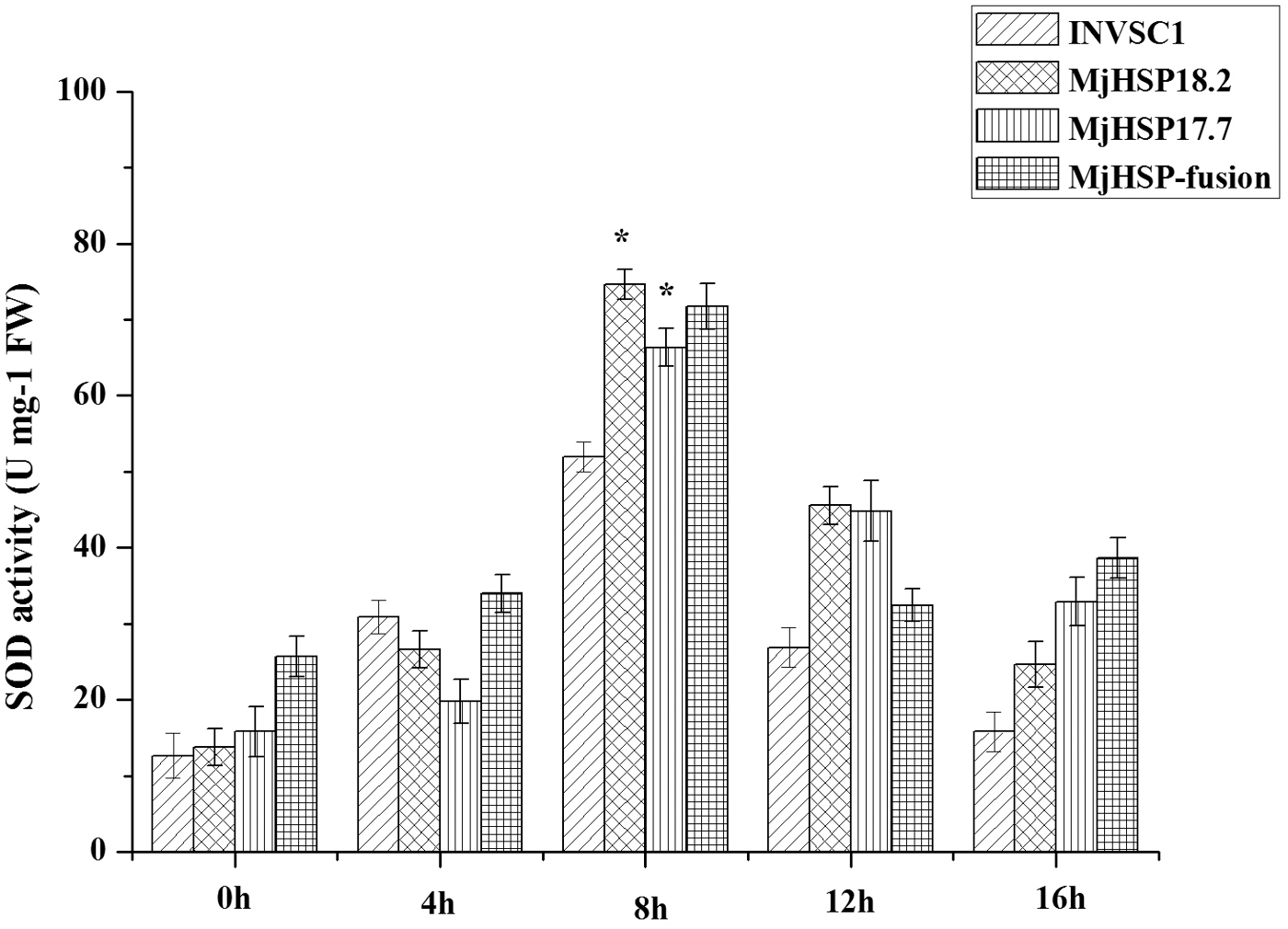
SOD activity assays of transgenic yeast cells and the yeast strain INVSC1 under tunicamycin stress. Yeast cells were treated with tunicamycin for 0, 4, 8, 12, and 16 h for SOD activity assays. Results were from three independent experiments. Bars show mean ± SD. *Indicates significant differences between transgenic yeast cells and the yeast strain INVSC1 at p < 0.05, according to Duncan's multiple range test. INVSC1, yeast strain under normal conditions for control; SOD, superoxide dismutase.
Discussion
Numerous studies have reported the role of sHSP in response to various environmental stresses in plants (Mahmood et al., 2010). However, few reports demonstrating the role of sHSP genes in pollution stress mechanism have been published to date. In this study, we isolated and characterized two full-length cDNA clones that encode MjHSP18.2 and MjHSP17.7 from M. jalapa. Sequence alignment and phylogenetic analysis showed that MjHSP18.2 and MjHSP17.7 belong to the cytosolic class I sHSP (Figs. 1 and 2). We established that MjHSP18.2 and MjHSP17.7 are involved in response to petroleum stress. Overexpression of MjHSP18.2 and MjHSP17.7 in yeast S. cerevisiae conferred tolerance to tunicamycin, and depressed expression of ER molecular chaperones, increased cell viability, decreased cell apoptosis, and increased SOD activities under tunicamycin stress condition.
sHSP production has now been found against many abiotic stresses (Sun et al., 2002; Sun and Macrae, 2005; Park et al., 2013; Yang et al., 2014). Petroleum stress is also one of the abiotic stresses. As shown in Fig. 3, the expression of MjHSP18.2 peaked at 30 g/kg at 12 h, while the peak of MjHSP17.7 was 20 g/kg at 24 h, which was in accordance with our previous predifferential proteome analysis. These results suggest that MjHSP18.2 and MjHSP17.7 may play an important role in response to petroleum stress. However, the molecular mechanism of MjHSP18.2 and MjHSP17.7 on petroleum stress tolerance needs validation. The yeast is an excellent eukaryotic model system to quickly identify genes involved in abiotic stress tolerance. The stress tolerance of many genes has been determined using the yeast expression system (Mahalakshmi et al., 2006; Jiang et al., 2009; Gao et al., 2011, 2012). So, we successfully generated overexpressing MjHSP18.2 and MjHSP17.7 yeast S. cerevisiae to further understand the function of MjHSP18.2 and MjHSP17.7 under abiotic stress (Supplementary Fig. S1). The ER is the site for protein synthesis, folding, modification, and transport, which is quite sensitive to the environmental stress. Almost all plants are constantly exposed to a multitude of abiotic stresses under natural conditions, which can disturb the protein-folding process and result in ER stress. ER stress causes the occurrence of UPR, and the molecular mechanism of UPR in yeast has been studied in great detail for many years (Gething, 1999; Noiva, 1999; Halperin et al., 2014). Similarly, tunicamycin can inhibit protein glycosylation modification, and the accumulation of unfolded proteins in the ER lumen will consequently lead to ER stress and induce expression of chaperones and foldases (Mori et al., 1992; Travers et al., 2000). In the present study, we found that the growth of Saccharomyces cells expressing MjHSP18.2, MjHSP17.7, and MjHSP-fusion was better than cells compared with controls under the same tunicamycin stress (Fig. 4). Consistently, the tunicamycin-induced expression of BiP and PDI1 rapidly increased in both transgenic and nontransgenic yeast cells, but this increase was less evident in the transgenic yeast cells compared with the nontransgenic yeast cells (Fig. 5). These results indicated that overexpressing MjHSP18.2 and MjHSP17.7 could enhance the tolerance to tunicamycin stress of Saccharomyces cells significantly, alleviate the ER stress caused by the tunicamycin stress in yeast cells, and may participate in the UPR signal transduction pathway.
A moderate ER stress plays a protective role by the mechanisms of cellular protection, and excessive or sustained ER can cause cell death or apoptosis (Rutkowski and Kaufman, 2004; Schröder and Kaufman, 2005). In this study, we found that the apoptosis rate of the MjHSP18.2 and MjHSP17.7 transgenic group was lower than that in the nontransgenic group under the same tunicamycin treatment, with a significant increase of Yac1 mRNA expression (Fig. 6A, B). Also, the cell survival rate of the unmodified yeast cells was significantly lower than that of the transgenic yeast cells (Fig. 6C). These results suggest that MjHSP18.2 and MjHSP17.7 play a role in reducing apoptosis and increasing cell viability in the condition of ER stress. Furthermore, overexpressed LeHSP21 protein of tomato in tobacco improved tolerance to high-temperature stress by enhancing antioxidation capacity (Neta-Sharir and Weiss, 2005; Zhang et al., 2016). Overexpression RcHSP17.8 enhanced SOD activity in transgenic Arabidopsis under various stresses (Jiang et al., 2009). In this study, we found that SOD activities were significantly higher in the transgenic yeast cells in response to tunicamycin stress compared with nontransgenic yeast cells (Fig. 7). The results showed that transgenic yeast cells had strong antioxidant capacities, and MjHSP18.2 and MjHSP17.7 enhanced the activity of S. cerevisiae in the stress of ER. Based on the results, we propose that MjHSP18.2 and MjHSP17.7 as molecular chaperones may enhance plant stress tolerance through the protection of antioxidative enzymes. However, the chaperone activity of MjHSP18.2 and MjHSP17.7 protein deserve further assays to confirm. Taken together, overexpressing MjHSP18.2 and MjHSP17.7 confer higher tunicamycin tolerance and resistance to ER stress. It provides molecular basis for the rapid screening of petroleum-contaminated plants.
Conclusion
Phytoremediation was regarded as an emerging cleanup technology for contaminated soils. However, the mechanism of how the plants cope with the abiotic stresses imposed by polyaromatic hydrocarbon and other organic pollutants is unknown. In this study, MjHSP18.2 and MjHSP17.7, two petroleum-induced sHSPs of M. jalapa, could alleviate the ER stress caused by tunicamycin and promote cell survival in yeast cells. In the future, it would be interesting to further explore the role of MjHSP18.2 and MjHSP17.7 in regulating environmental adaptation via modulating the intracellular redox state. This study provided molecular basis for the rapid screening of petroleum-contaminated plants.
Footnotes
Authors' Contributions
Q.S., H.L. and M.Z. conceived and designed the research. H.L., J.X., Q.S., and X.W. conducted the experiments. H.M. contributed new reagents. J.L. and S.C. analyzed the data. H.L. and J.X. wrote the article. All authors read and approved the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 31070448).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.