
Abstract
The control of infectious diseases is a critical challenge in developing and developed nations alike. Humans can be exposed to pathogenic microorganisms such as viruses and bacteria through numerous routes of transmission, including waterborne transmission. Accordingly, effective water disinfection processes are paramount for public health protection. In this study, the inactivation efficacy of P22 virus and Escherichia coli bacteria was tested using ultraviolet light-emitting diodes (UV LEDs), which provided effective disinfection of both waterborne microorganisms. Using a peak wavelength emission of 255, 265, or 285 nm, the batch-test, UV LED collimated beam system demonstrated high inactivation potential for both microorganisms at all wavelengths, suggesting that it could be an effective alternative to typical, low-pressure (LP) or medium-pressure (MP), mercury-based UV systems. Increased inactivation kinetics for both microorganisms was observed with decreasing wavelength (255 > 265 > 285 nm). However, the trend reversed when accounting for energy efficiency of the systems (255 < 265 < 285 nm) due to the currently lower wall plug efficiency for lower UV LED wavelength emissions. Substantial improvements in LED efficiency are needed to make them energetically competitive with current LP and MP UV technologies.

Introduction
Microbial pathogens, including viruses and bacteria, account for an annual global disease burden of more than 400 million years of life lost (exceeding that associated with either cancer or cardiovascular disease) (Naghavi et al., 2017; Fitzpatrick et al., 2019). Increasing world population and global interconnectivity magnify the risks of disease transmission through routes such as waterborne (e.g., adenovirus and Escherichia coli), foodborne (e.g., Campylobacter and Salmonella), and person-to-person (e.g., coronavirus and Ebola virus) transmission. Accordingly, effective means of microbial disinfection are essential for public health protection.
Ultraviolet (UV) irradiation offers an increasingly common approach to waterborne pathogen disinfection, with over 7,000 municipal UV installations around the world, primarily in Europe and North America (Muller, 2011; Song et al., 2016; Kheyrandish et al., 2018a). UV irradiation has high potential to inactivate most pathogens and is particularly effective against Giardia and Cryptosporidium, organisms that exhibit increased resistance to chlorine-based disinfection. However, inactivation of some microorganisms is far less efficient using UV irradiation. For example, spore-forming Bacillus subtilis is used as the European biodosimetry standard for low-pressure (LP) UV disinfection of drinking water due to its higher UV resistance relative to vegetative bacteria (Nicholson and Galeano, 2003).
Additionally, double-stranded DNA viruses demonstrate UV resistance. Of particular relevance is adenovirus, a nonenveloped icosahedral virus comprising an outer protein capsid and an inner core of double-stranded DNA. Human adenovirus is the most UV-resistant, waterborne enteric virus known and, as such, it drives UV fluence (It) requirements for target levels of microbial inactivation in water treatment (Gerba et al., 2002; Eischeid et al., 2009; USEPA, 2016). For example, the United States Environmental Protection Agency's (USEPA's) Ultraviolet Disinfection Guidance Manual specifies a UV fluence of 186 mJ/cm2 for 4-log inactivation of viruses despite the vast majority of viral contaminants being sufficiently mitigated using a fluence of 40 mJ/cm2 (Beck et al., 2013; USEPA, 2016).
The higher energy inputs needed to treat UV-resistant microorganisms such as adenovirus can make UV irradiation a cost-prohibitive drinking water treatment technology without the supplement of other treatment processes (Hijnen et al., 2006; USEPA, 2016). If required fluences can be reduced through the use of novel UV configurations, UV irradiation could be used to handle a range of treatment concerns at drinking water treatment plants.
Typical UV systems used in large-scale applications feature LP UV lamps. These lamps emit essentially monochromatic output near 254 nm, which aligns closely with the wavelength of peak DNA absorbance (≈260 nm) (Chen et al., 2009). Although most microbial inactivation occurs due to genomic damage during UV treatment, a fraction is also linked to protein damage (Wigginton et al., 2010, 2012; Mayer et al., 2015).
For viral contaminants that are not inactivated effectively using conventional LP UV irradiation, variable wavelengths may enhance damage to the protein capsid, which could potentially increase the effectiveness of UV irradiation for more UV-resistant contaminants, such as adenovirus (Tokuno and Gough, 1976; Mayer et al., 2015). For example, medium-pressure (MP) UV irradiation can be more effective than LP UV in treating human adenovirus (Linden et al., 2007). MP UV lamps emit a polychromatic distribution across the UV-C spectrum, which may facilitate improved degradation of compounds that effectively absorb light outside of the typical ideal germicidal range based on light absorption by DNA.
As a potential alternative to LP and MP mercury-based lamps, UV light-emitting diodes (UV LEDs) are an emergent option. UV LEDs offer many benefits, including high-intensity near-monochromatic outputs, longer life spans, and lower energy inputs (Würtele et al., 2011). They have also proven effective in inactivating bacterial, viral, and protozoan pathogens (Beck et al., 2017). UV LEDs offer variable wavelength emissions, which may facilitate disruption of a range of biological materials, including amino acids, in addition to nucleic acids. For example, proteins exhibit an absorbance peak near 280 nm, which could contribute to virus inactivation (Beck et al., 2017). This may enable tuning the UV LED treatment process to more effectively target a range of contaminants with different absorption spectra (Kheyrandish et al., 2018b).
However, as UV LED technology in the germicidal UV-C, or deep UV, range is a developing technology, with most of the work being done in the last decade at the bench scale, further research is needed to determine the viability of these systems as a feasible alternative to conventional LP UV systems (Argyraki et al., 2016; Song et al., 2016; Beck et al., 2017; Chen et al., 2017; Kheyrandish et al., 2017; Li et al., 2017).
To address this research need, the objective of this study was to determine the treatment efficacy of P22 virus and E. coli bacteria using UV LED disinfection at a peak wavelength emission of 255, 265, or 285 nm. Use of these nonpathogenic surrogate microorganisms as models of pathogen behavior during water treatment is common as they are safer to work with and easier to cultivate (Grabow, 2001; Sinclair et al., 2012; Heffron and Mayer, 2016). The well-characterized bacteriophage P22 was used as a surrogate for human adenovirus as both are similarly sized, icosahedral, double-stranded DNA viruses, making P22 a satisfactory model for treatability studies (Davies et al., 2009; Heffron et al., 2019). E. coli 15597 inactivation was assessed as E. coli and/or fecal coliform bacteria are typically used as indicators of disinfection efficacy for bacterial pathogens (Edberg et al., 2012).
Materials and Methods
This study received IRB approval through protocol BR-171.
Water matrix and microbes
All experiments and dilution series were prepared in pH 7 buffered demand-free water (BDF, comprising 0.54 g of G-Biosciences® Na2HPO4 and 0.88 g of Sigma-Aldrich® ≥ 98% KH2PO4 in 1.0 L of Milli-Q water). pH was measured with a VWR (Radnor, PA) B40PCID pH meter and adjusted to near-neutral levels using 0.1 M HCl and 0.1 M NaOH. Each test was performed in 14 mL of BDF spiked with an initial concentration of either 106 PFU/mL P22 bacteriophage or 106 CFU/mL E. coli.
Bacteriophage P22 (American Type Culture Collection [ATCC] 19585-B1) was used as a laboratory surrogate for human adenovirus. As described by Mayer et al. (2008), the virus was propagated using the double agar layer method and further purified to minimize organic content and monodisperse viruses using two sequential polyethylene glycol (PEG) precipitations (9%, PEG plus 1 M NaCl), followed by a Vertrel XF® (DuPont™) extraction to remove lipids from the system and promote equal distribution of viral particles. The host bacteria for the double agar layer method were Salmonella typhimurium LT2 (ATCC 19585).
E. coli (ATCC 15597) was tested as a representative bacterium. On the day of each experiment, bacteria were cultured to the log phase in BD Difco™ tryptic soy broth. To minimize organic oxidant demand/UV absorbance during experiments, log-phase E. coli 15597 was further processed using three cycles of centrifugation at 2000 rpm for 10 minutes, followed by decanting the liquid, and resuspending the bacterial pellet in BDF. Of note, the use of laboratory strains grown under ideal culture conditions facilitates controlled experimentation, but the efficiency of UV disinfection at full scale can vary due to physiological differences (e.g., culture growth phase) between laboratory studies and actual drinking water organisms, highlighting the value of full-scale tests under the conditions of interest.
UV LED tests
An AquiSense Pearlbeam™ UV LED collimated beam equipped with variable peak wavelength settings of 255, 265, and 285 nm was used for all UV tests. The 255 nm wavelength served as a comparison with conventional, monochromatic LP UV irradiation. The higher wavelengths (265 and 285 nm) may more effectively target some amino acids, thereby contributing to microbial inactivation.
Irradiance was measured at the water surface using an IL-1700 radiometer and SED 270 detector (purchased from, and calibrated by, International Light Technologies, Peabody, MA). Corrections were applied to calculate the average irradiance in the sample (Bolton and Linden, 2003), including the reflection factor = 0.975, Petri factor ≈0.894, water factor ≈0.981, and divergence factor = 0.975. A sensor factor accounting for polychromatic LED emissions was also applied using the relative lamp emission assessed using an Ocean HDX-UV-VIS spectrometer (Ocean Optics, Dunedin, FL) and wavelength-specific sensitivity of the radiometer detector (as reported by International Light Technologies) (Bolton and Linden, 2003). Finally a germicidal factor was applied based on UV absorption by DNA (Bolton and Linden, 2003; Beck et al., 2017). Accordingly, the adjusted average germicidal irradiances were calculated as 18, 125, and 157 μW/cm2 for the 255, 265, and 285 nm peak wavelength emissions, respectively.
Each water sample was spiked with 106/mL microbes in a sterile, 60 × 15 mm, oxidant demand-free quartz Petri dish. The sample was centered under the collimated beam with the water surface 2 mm from the end of the column (Ryu et al., 2008) and mixed at 20 rpm using a minimagnetic stir bar for sufficient time to achieve the targeted UV fluence (mJ/cm2). A range of fluences from ∼4 to 77 mJ/cm2 was tested under different treatment conditions (i.e., UV LED wavelengths and microorganism combinations) with fluence adjustments on a test-by-test basis to yield reportable data (plates within the countable range of 30–300 PFU or CFU per plate). All experiments were conducted in triplicate.
Microbial assays
After UV treatment, samples were diluted in BDF using a 10-fold dilution series. Infectious P22 bacteriophage was quantified using the double agar layer technique (Adams, 1959). Base plates were prepared using 1.5% tryptic soy agar (TSA, BD Difco, Franklin Lakes, NJ), while the top agar layer consisted of 0.7% TSA. Infectious E. coli was quantified using the membrane filtration method using freshly prepared BD Difco m Endo broth MF™ [in accordance with Method 9222 by APHA, AWWA, WEF (2012)]. Plates were enumerated after 24 h of incubation at 37°C.
Data analysis and statistics
The kinetics of microbial inactivation were modeled using the base-10, pseudo-first-order Chick–Watson approach applied to UV disinfection [Eq. (1)]:
where N = initial microbe concentration (PFU/mL or CFU/mL), No = microbe concentration following treatment (PFU/mL or CFU/mL), Λcw = Chick–Watson coefficient of specific lethality (mJ/cm2), I = UV irradiance (mW/cm2), and t = time (seconds).
The Chick–Watson coefficient of specific lethality was determined as the slope of the linear regression line for plots of logs of inactivation versus It (fluence [mJ/cm2]). The Chick–Watson relationship [Eq. (1)] is the most widely applied kinetic model for disinfection studies; however, nonzero intercepts are sometimes used since distorted model fits may arise when forcing the regression fit through the origin (Crittenden et al., 2012). Accordingly, results for both zero and nonzero intercept models (most often yielding very similar goodness of fit) are shown in Supplementary Table S1 of the Supplementary Data.
The energy efficiency of microbial inactivation was assessed as the electrical energy per order magnitude reduction (EEO, [Eq. (2)]), as described previously (Bolton et al., 2001; Bolton and Stefan, 2002; Sharpless and Linden, 2005; Beck et al., 2017):
where EEO = electrical energy per order or the amount of energy needed to decrease the concentration of a contaminant by one order of magnitude (kWh/m3) (Bolton et al., 1996); A = irradiated surface area (cm2); V = sample volume (L); WPE = wall plug efficiency reported by the manufacturer (0.3% for 255 nm, 0.7% for 265 nm, and 1.6% for 285 nm); and WF = water factor accounting for UV absorbance and depth of water. The factor of 3.6 × 106 accounts for conversions between hours and seconds, mW and kW, and m3 and L.
All statistics were computed at a significance level of α = 0.05 using GraphPad Prism 7 or Microsoft Excel. D'Agostino and Pearson normality tests were used to assess the normality of individual data sets. Statistical variance of data sets was tested using one- or two-way analysis of variance. Tukey's multiple comparison post hoc test was used to assess statistically significant differences among data points. Differences in inactivation efficacy (Λcw) were assessed using sum of squares F tests.
Results and Discussion
Inactivation of P22 bacteriophage and E. coli bacteria using UV LED was evaluated at three different peak wavelength emissions: 255, 265, and 285 nm.
P22 virus inactivation
Treatment using variable wavelength UV LED irradiation provided effective P22 bacteriophage inactivation, where increasing fluence significantly increased virus inactivation (Fig. 1, full statistics have been provided in Supplementary Table S2 in the Supplementary Data). Table 1 summarizes the Chick–Watson coefficient of specific lethality (Λcw) using the three different peak wavelength UV LEDs. System efficacy in terms of fluence-based kinetics significantly improved using lower wavelengths: UV255>UV265>UV285 (p < 0.0001). Similarly, Aoyagi et al. (2011) reported that UV255 provided greater disinfection efficiency compared with UV280 for bacteriophages MS2 and Qβ. However, they concluded that in spite of its lower efficacy, UV280 was more suitable for application in water treatment as it offers much higher external quantum efficiency (Aoyagi et al., 2011).

Kinetics of P22 inactivation using UV LED operated at a peak wavelength emission of 255, 265, or 285 nm. Each point represents the average of triplicate experiments and at least triplicate microbial assays. The error bars show ±1 standard deviation. Some error bars are too small to see. Solid lines indicate base-10 Chick–Watson pseudo-first-order kinetic model fit, while dotted lines show the 95% confidence intervals. UV LED, ultraviolet light-emitting diode.
Base-10 Chick–Watson Fluence-Based Kinetic Parameters (Coefficient of Specific Lethality, Λcw) and the Fluence Required to Achieve 4-Log Inactivation of P22 Bacteriophage Using Variable Wavelength Ultraviolet Light-Emitting Diodes
The kinetic rate constants (Λcw) were used to determine the fluence required to reach 4 logs of P22 inactivation (Table 1). As per USEPA Surface Water Treatment Rules, 4 logs of virus inactivation are required to meet drinking water regulations for surface water or groundwater under the influence of surface water (USEPA, 2016). The dramatic differences observed in the fluence requirements clearly illustrated that UV255 was much more effective for P22 inactivation compared with the other wavelengths tested (p < 0.0001). While not at peak DNA absorbance values, all wavelengths were still in the germicidal region (200–300 nm) (Beck et al., 2015), and therefore genetic damage ostensibly occurred in tandem with damage to the protein capsid of the bacteriophage.
The P22 inactivation efficacies observed here for UV255 and UV265 treatments were ∼30% and 16% better, respectively, than reports using the LP UV (UV254) treatment to inactivate other double-stranded DNA bacteriophages (PRD1) (Fig. 2). With increasing wavelength, the fluences to achieve 4-log inactivation increased, but were generally in line with typical LP UV and MP UV fluence requirements. Exceptions to this observation included φ-X174 (ssDNA), which was rapidly inactivated using all systems, and MS2 (ssRNA), which is reportedly more resistant to UV-C disinfection in comparison with other bacteriophages (Aoyagi et al., 2011).
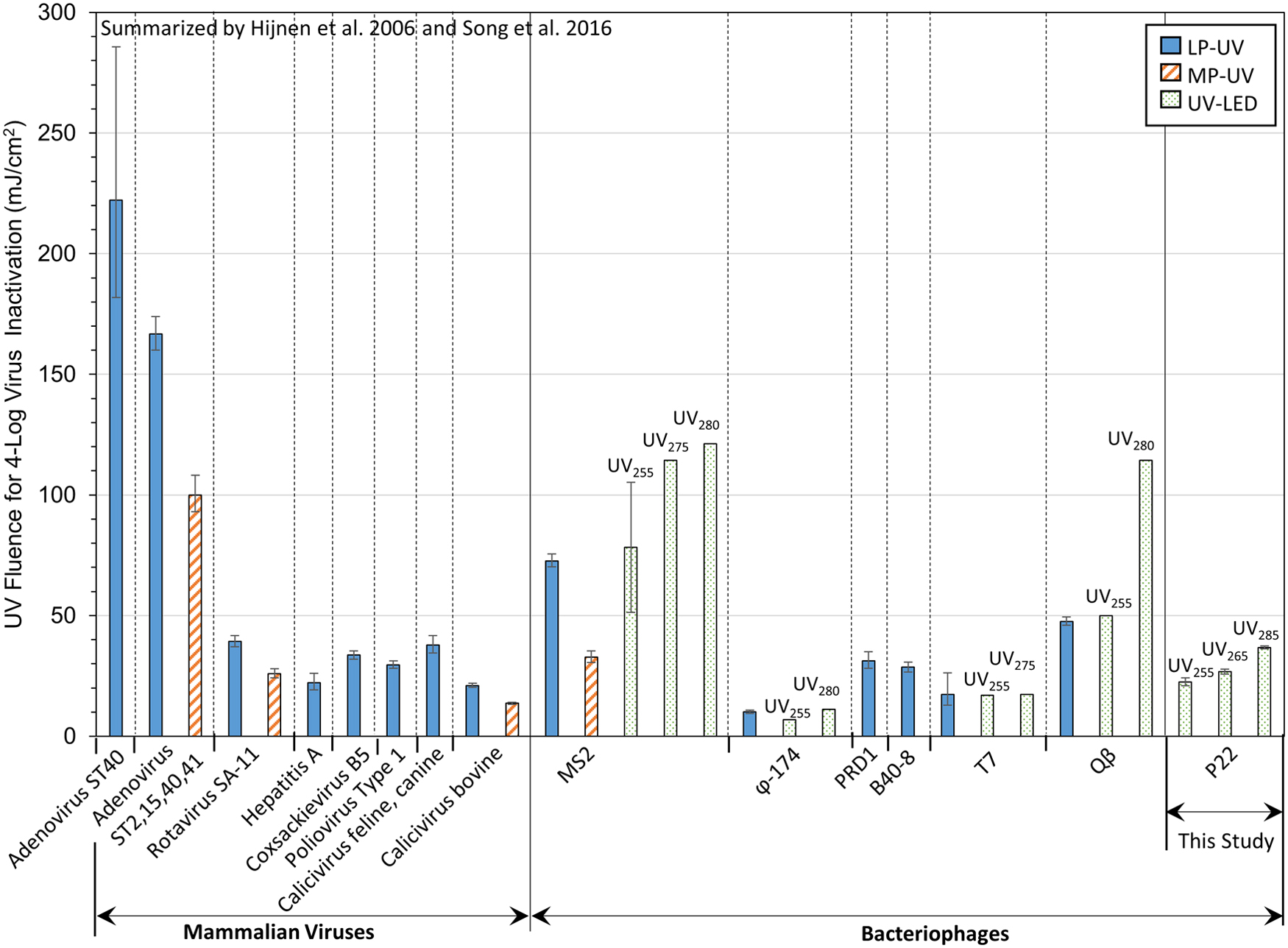
Comparison of virus inactivation efficacy using UV irradiation. Comparative LP UV and MP UV virus data were calculated from the first-order rate constants (k) summarized by Hijnen et al. (2006), where the error bars illustrate fluences determined using k ± 95% confidence intervals. Comparative UV LED virus data were calculated from the first-order rate constants summarized by Song et al. (2016). Data on the far right side of the figure are from this study using P22 bacteriophage, showing fluences calculated using Λcw ± 95% confidence intervals for at least triplicate experiments and triplicate microbial assays for UV255, UV265, or UV285 LED treatment. LP UV, low-pressure UV; MP UV, medium-pressure UV.
Moreover, Fig. 2 clearly shows that adenovirus demonstrates substantially greater UV resistance compared with other dsDNA viruses (P22 and PRD1) as well as all other viruses shown here; accordingly, future testing of adenovirus as a function of UV LED peak wavelength is critical. Notably, adenovirus appears to be less resistant to MP UV compared with LP UV treatment, a trend that is consistent for all viruses tested using both LP UV and MP UV systems (Fig. 2).
E. coli bacterial inactivation
E. coli was selected as the bacterial contaminant in this study as it is commonly used to assess the microbial quality of drinking water, both at full-scale and in laboratory studies of treatment processes. As minimal work has been done to evaluate UV LED treatment systems, demonstrating successful treatment of E. coli provides a reference point for comparison against more conventional UV configurations as well as future technology improvements. The USEPA's Guide Standard and Protocol for Testing Microbiological Water Purifiers (1987) specifies target reductions of 6 logs for bacteria, which provides a reasonable goal for bacterial inactivation.
Inactivation of E. coli using variable wavelength UV LEDs is shown in Fig. 3. As expected, increasing fluence significantly increased the corresponding bacterial inactivation for all wavelengths tested (a full summary of these statistics is provided in Supplementary Table S3 in the Supplementary Data). Table 2 summarizes the kinetic rates of inactivation (Λcw) as well as fluences needed to achieve the targeted level of 6-log inactivation. Following the same trend as the virus data, fluence-based inactivation efficiency decreased as wavelength increased, UV255>UV265> UV285 (p < 0.0001), with a notable drop in efficiency for UV285.

Kinetics of Escherichia coli inactivation for UV LED operated at a peak wavelength emission of 255, 265, or 285 nm. Each point represents the average of triplicate experiments and at least triplicate microbial assays. The error bars show ±1 standard deviation. Some error bars are too small to see. Solid lines indicate base-10 Chick–Watson pseudo-first-order kinetic model fits, while dotted lines show the 95% confidence intervals.
Base-10 Chick–Watson Fluence-Based Kinetic Parameters (Coefficient of Specific Lethality, Λcw) and the Fluence Required to Achieve 6-Log Inactivation of Escherichia coli Bacteria Using Variable Wavelength Ultraviolet Light-Emitting Diodes
As shown in Fig. 4, UV LED results from this study were comparable to other reports of E. coli inactivation using UV LEDs. However, compared to the inactivation of a range of bacteria in response to LP UV and MP UV treatment, inactivation of E. coli 15597 in this study generally required higher fluences to meet treatment targets using all three UV LED wavelengths tested. The notable exception to this finding was spore-forming Bacillus subtilis. Due to its demonstrated UV resistance, B. subtilis is commonly used in UV biodosimetry studies, including its use as the European biodosimetry standard for LP UV disinfection of drinking water (Nicholson and Galeano, 2003).
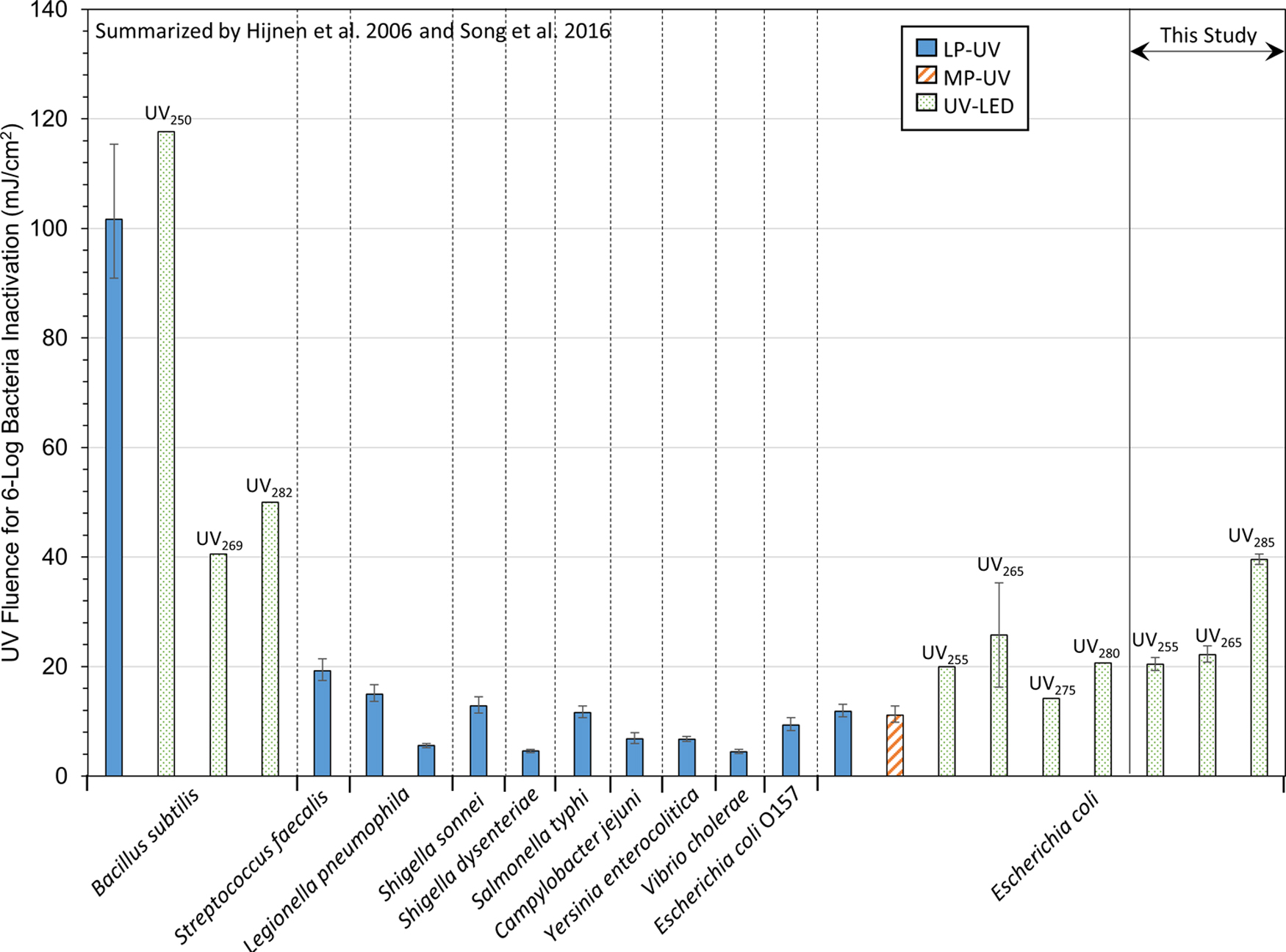
Comparison of bacterial inactivation efficacy using UV disinfection. LP UV and MP UV bacterial data were calculated from the first-order rate constants (k) summarized by Hijnen et al. (2006), without correction for environmental species. The error bars illustrate fluences determined using k ± 95% confidence intervals. Comparative UV LED data were calculated from the first-order rate constants summarized by Song et al. (2016). Data on the right side of the figure are from this study for E. coli 15597, showing fluences calculated using Λcw ± 95% confidence intervals for triplicate experiments and at least triplicate microbial assays for UV255, UV265, or UV285 LED treatment.
Energy efficiency
The predominant metric of UV efficacy has typically been fluence- or time-based kinetic rate constants, for example, coefficient of specific lethality, which translates into the fluence required to achieve a target level of inactivation. While this value remains important moving forward, the use of alternative light sources, such as UV LEDs, highlights the importance of additional metrics, including energy efficiency of the irradiation system.
Tunable UV-C LEDs offer tremendous promise in terms of targeting contaminants, smaller footprint/greater flexibility in design, robustness, being mercury free, instantaneous on/off, and greater lifetime (Bowker et al., 2011; Beck et al., 2017; Kheyrandish et al., 2018a). However, as the LED wavelength decreases, output efficiency currently decreases. Thus, unlike mature visible and UV-A LED technologies, the potential of UV-C LEDs has yet to actualize in municipal water treatment applications as they are currently characterized by low-power output efficiency, that is, higher cost per treatment (Lui et al., 2016).
To better reflect the differences in efficiencies among the three LEDs tested, EEO values were calculated to normalize the fluence-based kinetic data to electrical efficiency. Figure 5 illustrates that this reverses the efficiency trend, such that the order of energy efficiency was UV255<UV265<UV285 for treatment of both P22 and E. coli. This effect was similarly reported by Song et al. (2016), wherein wavelengths of 255 and 265 nm had greater germicidal effectiveness, but the higher wall plug efficiency of 280 nm compensated for lower germicidal effectiveness. This demonstrates that not only inactivation rate constants but also energy efficiency must be considered when selecting a UV LED emission wavelength (Rattanakul and Oguma, 2018).

Electrical energy per order magnitude reduction (EEO, where lower values are less energy intensive) for
Figure 5 also shows that the dsDNA P22 bacteriophage was inactivated more efficiently than the ssRNA MS2 bacteriophage using UV LEDs. Somewhat lower energy efficiency for E. coli inactivation was observed in this study compared with Beck et al.'s (2017) results (performed using the same UV LED testing apparatus), ranging from EEOs of 0.34 to 1.1 kWh/m3. Of critical import is that LP and MP UV systems are still much more efficient in converting input energy to germicidal output compared with UV LEDs (Fig. 5). Specifically, the wall plug efficiencies of Beck's LP UV and MP UV systems were ∼20–120 and 9–50 times greater than the UV LEDs used in this study, respectively.
While rapid changes in bulb properties are anticipated in the near term—Haitz's law states that every decade the amount of light generated by an LED increases 20 times, while the cost per lumen falls 10 times (Haitz's law, 2007; Autin et al., 2013)—current LED technology may be limited to point-of-use type of applications (Matafonova and Batoev, 2018). Accordingly, substantial advancements in UV-C energy efficiency are still needed before this emerging technology can be sufficiently cost competitive with conventional UV treatment to be practical for large-scale installations (i.e., wall plug efficiency must increase from current levels of <5% to ∼35%) (Autin et al., 2013; Beck et al., 2017).
Conclusions
The objective of this research was to determine the treatment efficacy of P22 virus and E. coli bacteria using variable wavelength UV LEDs. The results showed that UV LED at 255 nm offers potential as a feasible alternative to typical LP and MP UV disinfection systems based on effective inactivation of P22 and E. coli at relatively low fluences. As fluences increased, greater inactivation resulted. Higher wavelengths resulted in less efficient time-based kinetics of inactivation of both viruses and bacteria (UV255>UV265>UV285). However, in terms of energy efficiency, the trend reversed (UV255<UV265<UV285) as lower wavelength LED systems are currently characterized by much lower wall plug efficiencies. This highlights the need for substantial advances in LED technology before practical implementation at municipal water treatment scales. In the interim, current LED technology may be limited to point-of-use type of applications.
Beyond the two microorganisms tested here, future work must consider the treatability of other contaminants. For example, it is imperative to directly test more robust viral contaminants, such as human adenovirus [highly UV resistant, with a more robust capsid than its P22 counterpart (Tokuno and Gough, 1976; Susskind and Botstein, 1978; Walker, 1978; Prevelige et al., 1988; Foguel et al., 1995)] or rotavirus, which is another commonly analyzed enteric virus. Direct testing of other structurally dissimilar microorganisms (e.g., spore-forming bacteria and enveloped viruses) is also needed to deepen understanding of the overall microbial mitigation potential of UV LEDs.
Notably, although enveloped viruses (e.g., SARS-CoV-2, the virus responsible for the 2020 COVID-19 pandemic) are typically less persistent in water compared with nonenveloped viruses, UV susceptibility is expected to be similar for enveloped and nonenveloped viruses based on the mechanism of microbial genomic damage under UV irradiation (Ye et al., 2018; Wigginton and Boehm, 2020). By directly analyzing the disinfection responses of pathogenic viruses and bacteria under variable water quality conditions and UV LED reactor configurations, a clearer picture of the potential for these novel treatment methods can be established.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.