
Abstract
The wide and increasing use of nano-titanium dioxide (nTiO2) has led to its release in the environment that will likely impact aquatic eco-relevant biota. In this study, we investigated the impact of sublethal concentrations of nTiO2 on the nitrogen (N) metabolism of the primary producer, nitrogen-fixing cyanobacteria Anabaena PCC 7120, using transcriptional-level information of biomarker genes involved in global N regulation, N fixation-, N assimilation-, and N storage-specific pathways. The results showed that both the circadian patterns of cyanobacterial metabolism and nTiO2 intrinsic properties distinctively governed the toxicity responses during light and dark exposure to nTiO2. During illuminated conditions, the majority of genes linked to cellular nitrogen status exhibited a clear upregulation in the dose range of 6–60 mg/L·h, whereas overall gene downregulation was mainly observed at the end of dark cycle, characterized by low cell metabolism and energy (ATP) levels. The nTiO2 dose-dependent production of intracellular metabolites (amino acids) involved in both the GS-GOGAT pathway of N assimilation and the intracellular N storage pathway suggests that pathways involving amino acid biosynthesis or degradation might be activated or repressed. This, possibly, contributes to the increase of newly synthesized proteins needed for detoxification in response to the cellular stresses induced by nTiO2 treatment. These findings suggest that under environmental perturbations generated by nTiO2, cyanobacteria are likely to modify their intracellular carbon and nitrogen balance, thus impact, at a larger scale, ecological trophic interactions and food web dynamics within complex ecological systems.
Introduction
Emerging water pollutants are responsible for a significant share of risk with respect to ecological integrity and human health (Daughton, 2010; Petrie et al., 2015; Barbosa et al., 2016). The advancements in nanotechnology and application of engineered nanomaterials (NMs) have raised public concerns for their potential environmental and health impacts. Nanosize titanium dioxide (nTiO2) is among the most widely used NMs due to its excellent electrical properties and optical performance (Abdullah and Kamarudin, 2017; Mezni et al., 2018), and its projected production quantities are up to 7,800–38,000 tons/year (Mueller and Nowack, 2008; Hendren et al., 2011). The increased manufacture will inherently result in increased environmental exposure (Chapman, 2006; Leite et al., 2020). Effluents from wastewater treatment plant tertiary treatment have been previously reported to show detectable levels of nTiO2 ranging from <5 to 15 μgTi/L (Kiser et al., 2009; Zeumer et al., 2020), whereas nTiO2 concentrations in surface water were estimated to be in the range of 3 ng/L to 110 μg/L based on models of typical exposure scenarios (Mueller and Nowack, 2008; Gottschalk et al., 2013; Liu et al., 2019).
In the past years, increasing efforts have been made toward the understanding of the potential toxicity of NMs. Most of these studies have focused on occupational exposure and associated human health impact, mainly revealing cyto- and genotoxic effects of NMs on those routes with higher likelihood of exposure (i.e., inhalation, ingestion, and contact) (Klaine et al., 2008; Bierkandt et al., 2018; Pietroiusti et al., 2018). Today, knowledge on the ecotoxicological impact of NMs (i.e., metal nanoparticles, nTiO2) is rapidly growing, especially on freshwater organisms; interactions of nTiO2 with photosynthetic and nitrogen-fixing species (plant and phytoplankton) may lead to modifications of the organisms' membrane integrity (Hartmann et al., 2010; Cherchi et al., 2011), mechanical properties (Cherchi et al., 2011), physiology (Hartmann et al., 2010), and metabolic functions (Yang et al., 2006; Asli and Neumann, 2009; Nair et al., 2010; Cox et al., 2017; Hou et al., 2019). However, several gaps have been identified on the toxicological studies of NMs on aquatic species in the environmental health assessment context, such as limited range of test species and endpoints (Hall et al., 2009; Hartmann et al., 2010; Kulacki and Cardinale, 2012; Du et al., 2019), and lack of systematic understanding of the associated mechanisms (Rocha et al., 2017; Lekamge et al., 2020). Moreover, studies on cellular physiological changes at environmentally relevant subcytotoxic concentrations of nTiO2 are scarce. Particularly, one of our previous studies has observed the dose-dependent accumulation of cyanophycin (a nitrogen-rich storage polymer) in Anabaena sp. after treatment with nTiO2, raising a question on whether cell nitrogen (N) metabolism was more globally impacted (Cherchi and Gu, 2010).
In filamentous diazotrophic cyanobacteria species, such as Anabaena sp., the fixation of dinitrogen is catalyzed by the enzyme nitrogenase within terminally differentiated heterocysts, providing fixed nitrogen to vegetative cells and in turn receiving fixed carbon produced from their oxygenic photosynthesis (Burnat et al., 2014; Herrero et al., 2016; Harish, 2020). Hence, the coordination of carbon and nitrogen metabolism and intracellular nutrient balance is fundamental to cyanobacteria for maintaining productivity and acclimating to perceived stress in changing environments (Schwarz and Forchhammer, 2005; Jiang et al., 2018; Zhang et al., 2018). Cyanobacteria sense their intracellular nitrogen balance through the global PII and NtcA regulators, which relay signals to the gene expression machinery of main pathways associated with nitrogen and carbon metabolism, photosynthesis, and stress responses (Frías et al., 1994; Herrero et al., 2001; Forchhammer, 2004; García-Fernández and Diez, 2004; Zhang et al., 2018). The activity of NtcA, a transcriptional regulator synthetized by ntcA gene, is triggered by the accumulation of 2-oxoglutarate (2-OG), a known intermediate of the Krebs cycle, which provides the carbon skeleton for nitrogen assimilation and acts, along with glutamine, as important signaling molecules within the glutamine synthetase-glutamate synthase (GS-GOGAT) pathway (Herrero et al., 2001).
In this study, we hypothesized that exposure to subcytotoxic level of nTiO2 can lead to potential alterations in the molecular activities controlling the nitrogen and carbon metabolisms in cyanobacteria. The consequent changes in the level of intracellular metabolites perceived by the sensing systems as well as the initiation of a more economic usage of intracellular resources, might impact cells' perception of extracellular nutrients' availability, cellular nutrients' uptake, and the flux of energy between the organism and the surrounding environments (Muller et al., 2009). We examined the regulatory networking linked to the N metabolism in the cyanobacterium Anabaena PCC 7120 (hereafter Anabaena sp.) under exposure to nTiO2 at environmentally relevant concentrations. Temporal gene expression patterns during light and dark cycle of key marker genes associated with global regulatory pathways (ntcA, glnB), N fixation (nifH, nifK), N assimilation (glnA, all2934), and N storage (cphA, cphB) were investigated to elucidate the effect of nTiO2 on the global response, on the interplay between nitrogen and carbon metabolism, and on the modulation of N reserve inclusions in cyanobacteria. Intracellular nitrogen metabolites (i.e., amino acids) and the total proteins content have also been monitored along with gene expression trends, to reveal additional toxicity responses influencing nitrogen metabolism and the dynamics of nitrogen accumulation. Because of the important role of nitrogen in cyanobacterial ecology, molecular mechanisms regulating nitrogen metabolism in response to environmental stressors (i.e., exposure to pollutants) will progress our understanding on the adaptability of cyanobacteria to emerging environmental stimuli, such as NMs.
Materials and Methods
NMs preparation and characterization
Anatase TiO2 nanoparticles were chosen as the test substance in this study due to their broader applications (Leite et al., 2020), higher chemical reactivity (Zhang et al., 2015), and stronger cytotoxic effect (Wang et al., 2019) compared with the other two crystalline forms of TiO2 (rutile and brookite). Nano-TiO2 anatase (nTiO2, < 25 nm particle size, specific surface area 45–55 m2/g, purity 99.7% on trace metal basis; Sigma-Aldrich) was prepared in a nitrogen-free BG-11 medium and then dispersed through sonication in a high-energy cup sonicator (Brandson, Danbury, CT), at ∼90 W power for 15 min with 30 s pulse (Pal et al., 2012). Tween 20 (1‰) was added to enhance the dispersion for uniform dosing (El-Temsah and Joner, 2012). Particle size after dispersion in BG-11 media was measured through Dynamic Light-Scattering Analysis using a Malvern Nanosizer ZS90, and counted an average 689.8 ± 102.5 nm. The polydispersity index after dispersion in culture media was detected to be 0.549 and zeta potential reached −23.5 ± 3.95 mV (Supplementary Table S1).
Culture conditions and ecotoxicological tests
Anabaena PCC 7120 (UTEX No. 2576) was cultured at 20°C in a nitrogen-free BG-11 medium, as described in our previous work (Cherchi et al., 2011). For exposure tests, 1,800 mL volumes of an initial chlorophyll a concentration of 1.5 mg/L were incubated under a 12-h light/12-h dark regime to mimic natural environmental conditions. The light source consists of 1:1 ratio of 34 W cool white and 40 W Gro-Lux fluorescent bulbs (Sylvania, Danvers, MA), which yields a low photosynthetically active radiation value of 35 μE/[m2∙s]. The lamp wavelength output >400 nm is beyond the ultraviolet region (<400 nm) known to promote the photocatalytic activity of the anatase crystalline form of nTiO2 (Linsebigler et al., 1995). Batches were continuously mixed (300 rpm) and periodically aerated using compressed 0.2 μm filtered air. Cultures were exposed to different environmentally relevant nTiO2 concentrations, at 0 mgTiO2/L (control sample with same media matrix), 0.01, 0.1, 1, and 10 mgTiO2/L for an entire 24-h light/24-h dark cycle (L-D). Samples for analysis were collected at 0.25, 1.5, 3, and 6 h during light cycle, and at 12, 18, and 24 h during dark cycle. Exposure tests were performed in duplicate batches and analysis of each sample was performed in triplicates.
Chemical analysis of biomass and cellular components
Chlorophyll a was measured after ethanol extraction by a Synergy HT fluorometer (BioTek, Winooski, VT) with excitation and emission at 440 and 670 nm, respectively. Biomass dry weight was also measured according to previous studies (Zhu and Lee, 1997). Total proteins were assayed colorimetrically on sonicated cells (75 W, 60 s), with the Bicinchoninic Acid Protein Assay Kit (Pierce Chemical Co., Rockford, IL) and standards prepared with bovine serum albumin. All measurements were performed in triplicates.
Amino acid quantification
Quantification of amino acid content in algal cells was performed according to the method by Bartolomeo and Maisano (2006). Briefly, 15 mL of cell cultures were centrifuged, lyophilized, and then extracted with 0.5 mL boiling 6 M HCl at 110°C for 24 h. Extracted samples were centrifuged and hydrolyzed again to obtain dried residues, which were resuspended in 100 μL 0.1 M HCl. After a semiautomated derivatization with o-phthalaldehyde (OPA), samples were injected (5 μL) and analyzed on the Agilent 1260 Infinity Quaternary LC (Agilent, Inc., Santa Clara, CA) using an Eclipse AAA (4.6 × 150 mm, 5 μm) column. Mobile phase A was 40 mM NaH2PO4 with pH adjusted to 7.8, and mobile phase B was acetonitrile/methanol/water (45:45:10 in v/v/v). Amino acid separation was obtained at a flow rate of 2 mL/min with a programmed linear gradient program from a 1.9-min step using mobile phase A to a 16.3-min step for B. The following amino acids (retention times, min) were monitored: arginine (8.4), aspartic acid (1.2), glutamine (6.9), and glutamic acid (3.2). L-norvaline (retention time = 13.5 min) at 0.5 mM was used as internal standard for chromatographic spectra normalization. Calibration curves of the amino acids of interest were obtained by diluting the amino acid standard mixture (Pierce Biotechnology, Rockford, IL) in the concentration range of 10–500 μmol/mL. Measurements were performed in triplicates.
Monitor gene expression changes using quantitative reverse transcription polymerase chain reaction procedure
According to Simms et al. (1993), total RNA was isolated from 100 mL of mid-exponential phase cultures of Anabaena PCC 7120, precipitated with isopropanol, and washed with ethanol before final resuspension in RNAse-free water. The concentration and purity of the extracted RNA were determined based on the absorbance measurements at 260 and 280 nm, using Take3 Micro-Volume Plates (Synergy HT Microplate Reader; Biotek). Synthesis of complementary DNA (cDNA) was conducted using a Verso™ Reverse Transcriptase (Thermo Scientific, Barrington, IL) with random hexamers, according to the manufacturer's instructions.
RT-qPCR reactions were carried out in a Bio-Rad iQ5 Thermocycler (Bio-Rad, Hercules, CA). The amplification reactions were cycled as follows: 95°C for 3 min, 45 cycles at 94°C for 30 s, an annealing temperature primer specific (Supplementary Table S2) for 30 s, at 72°C for 30 s, and final elongation at 72°C for 3 min. The cphA primer was design using the Primer-BLAST design tool from NCBI and confirmed by endpoint melting curve (Supplementary Fig. S1). All other primers used in this polymerase chain reaction (PCR) was obtained from literature and listed in Supplementary Table S2. The absence of unspecific products in PCR was confirmed by the endpoint melting curve analysis. PCRs were run in triplicates.
Data analyses and statistics
Data analyses for a relative quantification of gene expression were performed by the comparative CT (threshold cycle) method. Fold changes of gene expression were normalized using the housekeeping rnpB gene, the RNA concentration, and the genomic 16S ribosomal DNA (rDNA). Clustering analysis and visualization were performed using the MeV (MultiExperiment Viewer) software version 4.8.1 (www.tm4.org/mev). Hierarchical clustering of gene trees and sample trees construction was performed employing Euclidean distance using an average linkage clustering. Statistical significance of gene expression and amino acid quantification data were investigated using a pairwise, two-tailed t-test at p < 0.05.
Results and Discussion
Distinct N metabolism transcriptional profiling under light and dark cycle
The linked pathways involved in nitrogen fixation, metabolism, and storage in vegetative and heterocyst cells of Anabaena sp. were illustrated in Fig. 1. Nitrogen fixation is carried out in heterocyst, where the enzyme nitrogenase catalyzes the conversion of one mole of dinitrogen into two moles of ammonia (Burnat et al., 2014; Herrero et al., 2016; Zhang et al., 2018). The energy (ATP) and reducing power (NADPH) required for the reaction are, respectively, provided by the photosystem I (PSI)-mediated cyclic photophosphorylation and the oxidative pentose phosphate cycle (PPC) through the electron carrier heterocyst ferredoxin (FdxH) (Bothe et al., 2010; Issa et al., 2014). The fixed ammonia is assimilated in the GS-GOGAT cycle (Wolk et al., 1976), through the reaction with glutamate to form glutamine, which is then delivered to vegetative cells and subsequently transformed into glutamate along with 2-OG, and is finally recycled back into the heterocyst (Böhme, 1998; Burnat et al., 2014; Issa et al., 2014; Zhang et al., 2018).
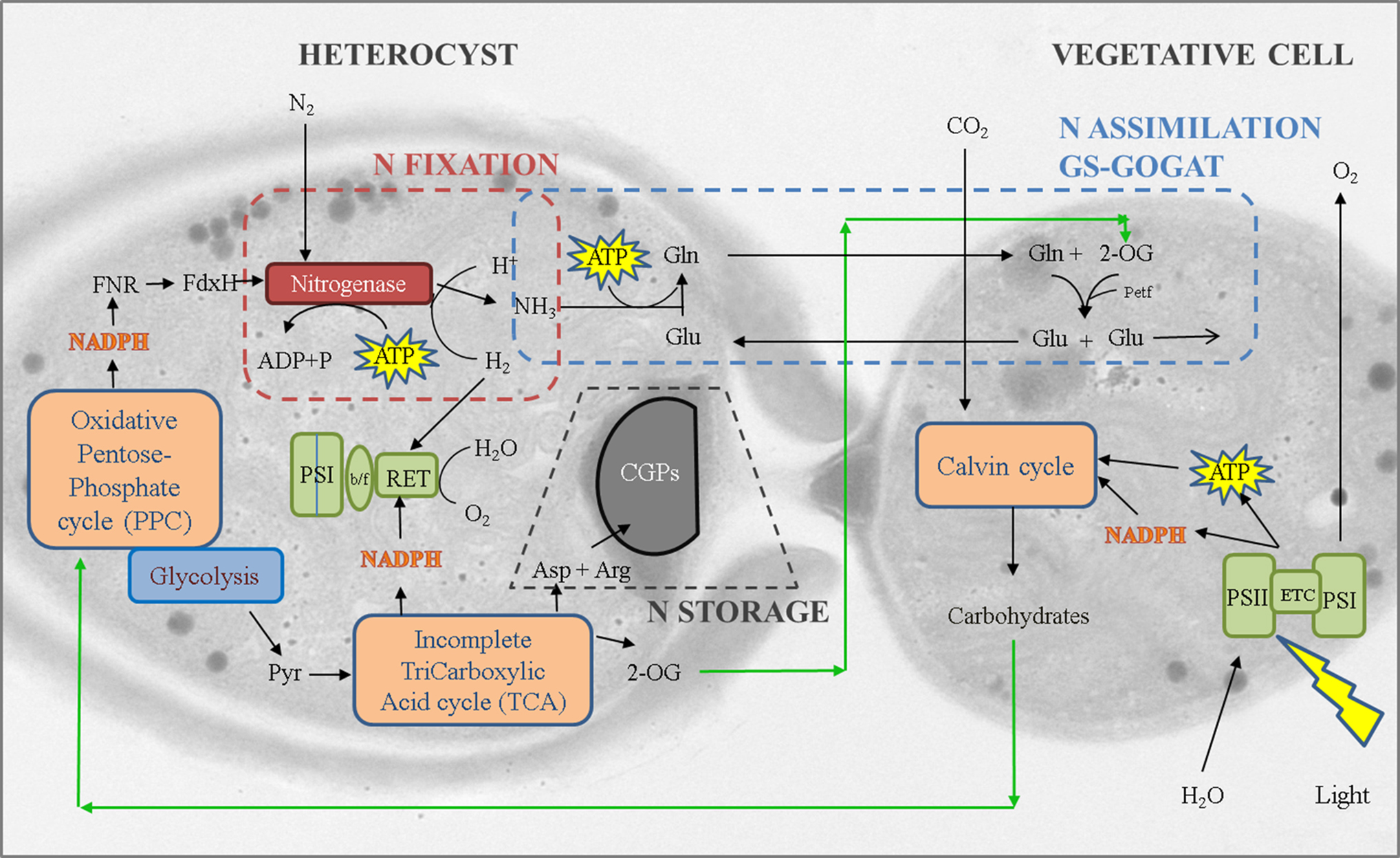
A transmission electron micrograph showing a heterocyst cell (left) and its adjacent vegetative cell (right) of Anabaena sp., overlaid with the schematic of pathways involved in the cell nitrogen metabolism. 2-OG, 2-oxoglutarate; Arg, arginine; Asp, aspartic acid; b/f, cytochrome b6f complex; CGPs, cyanophycin grana proteins; ETC, electron transport chain; FdxH, heterocyst ferredoxin; Gln, glutamine; Glu, glutamate; Petf, vegetative cell ferredoxin; PSI and PSII, photosystem I and II; Pyr, pyruvate; RET, respiratory electron transport.
Eight genetic biomarkers associated with the activity of above key N metabolism pathways were monitored under exposure to different concentrations of nTiO2 for 24 h (Supplementary Table S1). Concentration-specific and time-dependent transcriptional patterns and magnitude of expression levels (fold-changes) are shown in Fig. 2. Distinctive transcriptional effects were observed for dark and light cycles, likely due to the essential role of light in photosynthesis and metabolism (Golden, 1995), its direct interconnection with nitrogen metabolic network (Lopez-Gomollon et al., 2007), and the recognized photo-induced toxicity of nTiO2 (Wiesner and Bottero, 2007). During light exposure, hierarchical clustering grouped similar expression profiles of all genes into three major clusters (cutoff distance 2.64) based on treatment concentrations and exposure time lengths (Fig. 2A). Interestingly, the gene expression profiles reflecting immediate responses at 0.25-h exposure clustered together disregarding of different doses, indicating similar subcellular responses at the very early stage of nTiO2 exposure. Three treatments with relatively lower doses, defined as CT (concentration × time) <0.15 mg/L·h, clustered together with most of genes showing downregulation compared with untreated control. However, as doses increased to be higher than 6–60 mg/L·h, the same genes exhibited clear and significant upregulation. The dose-dependent distinctive clustering patterns indicate that the subcellular responses are sensitive to both nTiO2 concentration and exposure time and, nitrogen metabolic activities respond quite differently depending on the dose.

Temporal expression profiles (fold change) of genes describing Anabaena PCC 7120 nitrogen status in illuminated
In contrast, during dark conditions (Fig. 2B), exposure time, rather than nTiO2 concentration, governs the grouping patterns with treatments at the initial (12 h), intermediate (18 h), and final phases (24 h) of dark cycle clustered separately. While minimal gene expression levels were observed at the beginning of the dark stage, most genes showed upregulation after 6 h into the dark cycle (18 h) and finally transited into downregulation at the end of the dark cycle (24 h). The observed distinctive light/dark cycle-dependent nitrogen metabolism patterns demonstrated the complex subcellular responses of Anabaena sp. in exposure to nTiO2 that may be associated with a battery of external and internal impact factors such as toxicant doses, light, cellular circadian metabolism regulation, and energy (ATP) levels.
Dynamics of nitrogen storage under nTiO2 exposure
In aquatic environments, cyanobacteria are known to implement unique metabolic strategies when “sensing” adverse conditions to their optimal growth. Cells are known to initiate a “forward planning” (Oppermann-Sanio and Steinbüchel, 2002), adjusting the redistribution of internal resources, such as in the synthesis and accumulation of intracellular storage polymers. In our previous work, a dose-dependent accumulation of the nitrogen-rich cyanophycin grana proteins (CGPs) in Anabaena sp. was observed during the 96-h treatment of nTiO2 (Cherchi and Gu, 2010). CGP is a copolymer of arginine and aspartic acid, serving as a dynamic reservoir of newly fixed nitrogen before redistribution to vegetative cells, or to overcome periods of nutrient shortage (Sherman et al., 2000). Synthesis and degradation of CGPs are catalyzed by cyanophycin synthetase (encoded by cphA) and cyanophycinase (encoded by cphB), respectively.
In this study, we monitored the transcription levels of cphA and cphB genes involved in the CGP metabolism, as well as the cellular arginine and aspartic acid concentrations during the nTiO2 exposure (Figs. 3 and 4). An immediate upregulation of cphA gene involved in the CGP synthesis was observed at the beginning of exposure, whereas no significant changes were detected in CGP degradation, suggesting the increase of the nitrogen reserves as one of the initial autoprotective mechanisms of the cells to nTiO2 toxicity (Fig. 3). As exposure time increased to 6 h, only the highest nTiO2 concentration of 10 mg/L exerted the stimulating effects on both CGP synthesis and degradation. After transition to dark growth cycle, the cphA gene continued to be upregulated under most exposure scenarios, while the cphB gene concurrently shifted to be mostly downregulated regardless of nTiO2 concentration and exposure time. Figure 4 shows significant changes in intracellular amino acid pools of arginine and aspartic acid after the nTiO2 treatment at >0.1 mg/L compared with the untreated control (p < 0.001). Both arginine and aspartic acid exhibited dose-dependent increase over time and highest levels of amino acid accumulation occurred at L-D transition time points (12 and 24 h).
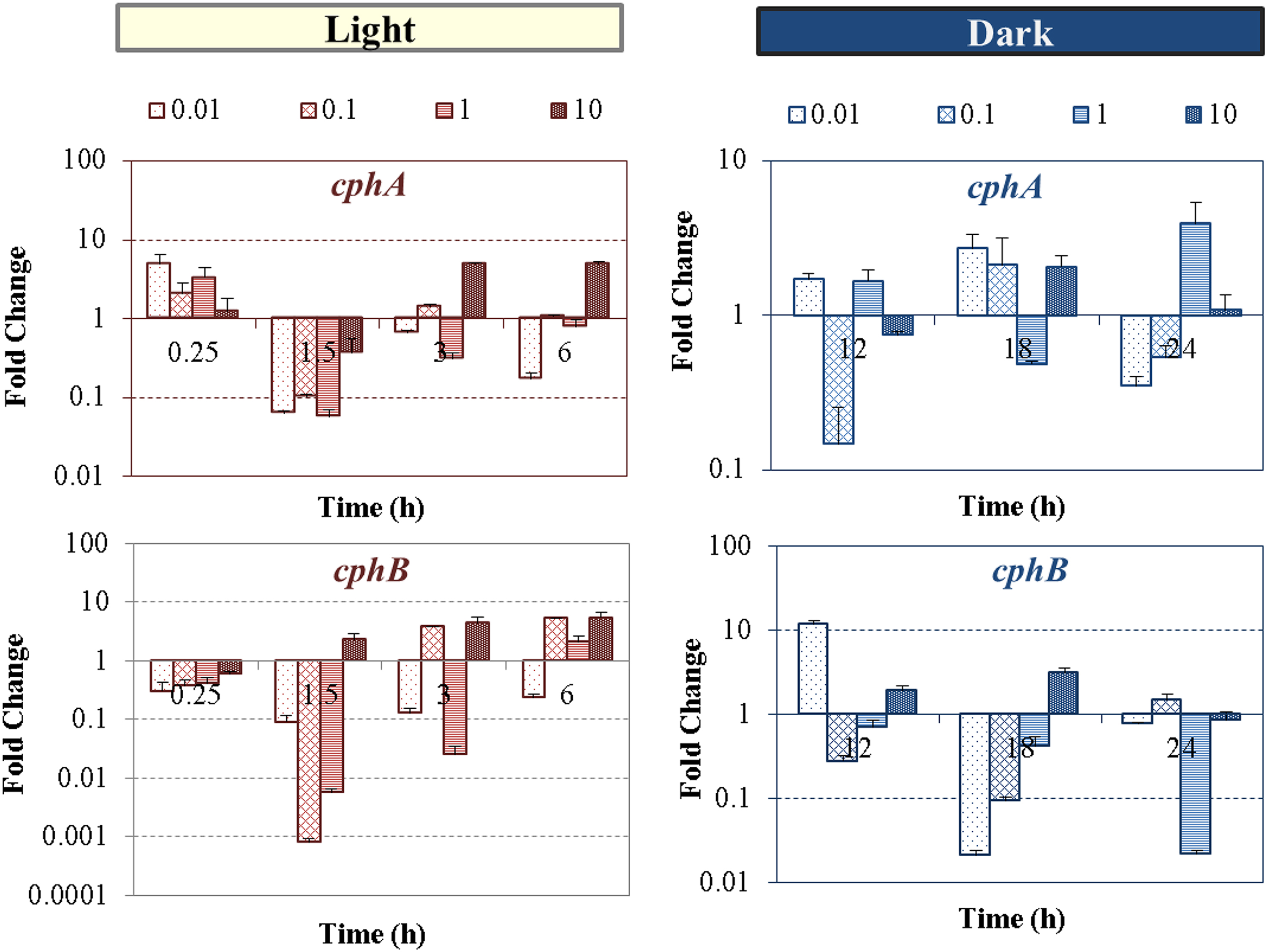
Time-dependent levels of transcriptional expression fold changes of genes involved in nitrogen storage (cphA, top and cphB, bottom) of Anabaena PCC 7120 exposed to nTiO2 concentrations ranged from 0.01 to 10 mg/L at different times of light/dark regimens. Standard deviations are of PCR reactions. PCR, polymerase chain reaction.
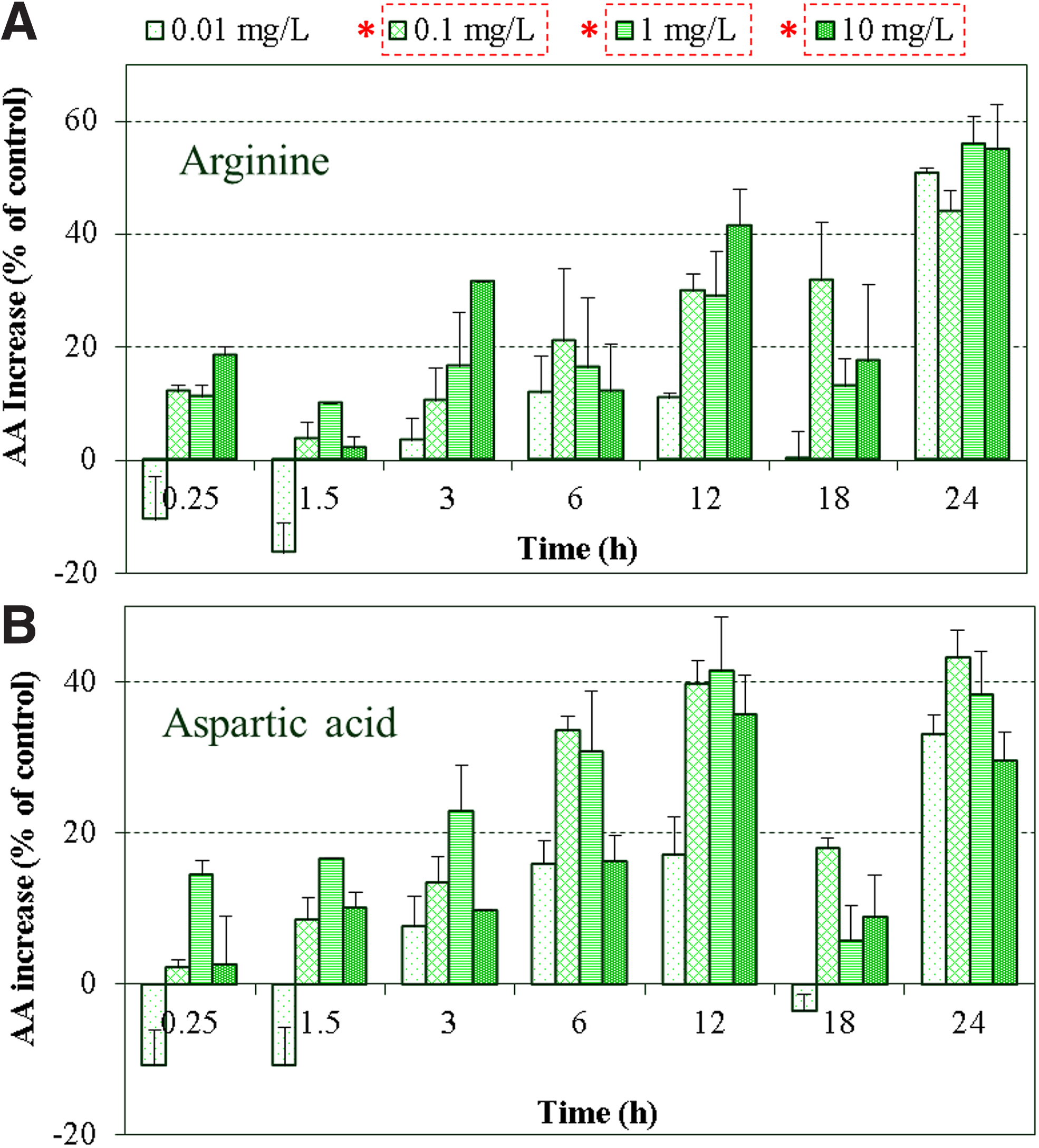
Time-dependent increase of arginine
The mechanisms regulating cyanophycin accumulation have not been fully elucidated (Maheswaran et al., 2006), therefore, definite correlations between the rate of CGP accumulation and cyanophycin synthetase activity are yet difficult to confirm. Nevertheless, the progressive increase of the intracellular metabolites, which are essential for CGP formation induced by nTiO2 treatment, still clearly indicates its impact on cellular N storage processes. Arginine is the amino acid with the highest nitrogen content (Burillo et al., 2004) that can be catabolized by multiple pathways (Maldener and Muro-Pastor, 2001). It serves as a source for protein synthesis and a nitrogen buffer within cyanophycin (Maheswaran et al., 2006). Aspartic acid can be utilized for the synthesis of methionine, lysine, and threonine (Wegener et al., 2010) if needed in response to stress, or can combine with 2-OG to produce glutamine, which can then be combined with ammonia to produce glutamate.
Despite the observed downregulation of the four genes involved in nitrogen fixation (nifH, nifK) and assimilation (glnA, all2934) at the end of dark phase (Figs. 5 and 6), the production of arginine and aspartic acid was significantly increased compared with the untreated control after nTiO2 treatment (p < 0.001), suggesting the activation of alternative pathways by cells to provide nitrogen for storage (Rabouille et al., 2006) or for protein synthesis (Daday et al., 1988). In this regard, the increase in total protein production was also detected in this study (Supplementary Fig. S2) and found to be, for the most, both time and concentration dependent, reaching over 50% increase (of the control) in the sample exposed to 10 mgTiO2/L during the dark phase. The overproduction of amino acids might partially contribute to the increasing synthesis of new detoxifying proteins needed to cope with other cellular stresses besides nitrogen metabolism that were likely induced by nTiO2 treatment, such as previously reported inflammatory effect, DNA damage, or oxidative stress (Xue et al., 2010; Wang et al., 2019). Indeed, previous studies have identified a series of proteins and peptides that actively participate in the detoxification of cellular stresses induced by various external stimuli in algae (Balzano et al., 2020); among them, metallothionein has been suggested to act as free radical scavengers to protect cells from oxidative stress (Thornalley and Vašák, 1985), while glutathione and phytochelatin have been recognized as the main antioxidants in eukaryotes and are mostly involved in the cellular defense against oxidative and metal stress (Singh et al., 2018; Balzano et al., 2020).
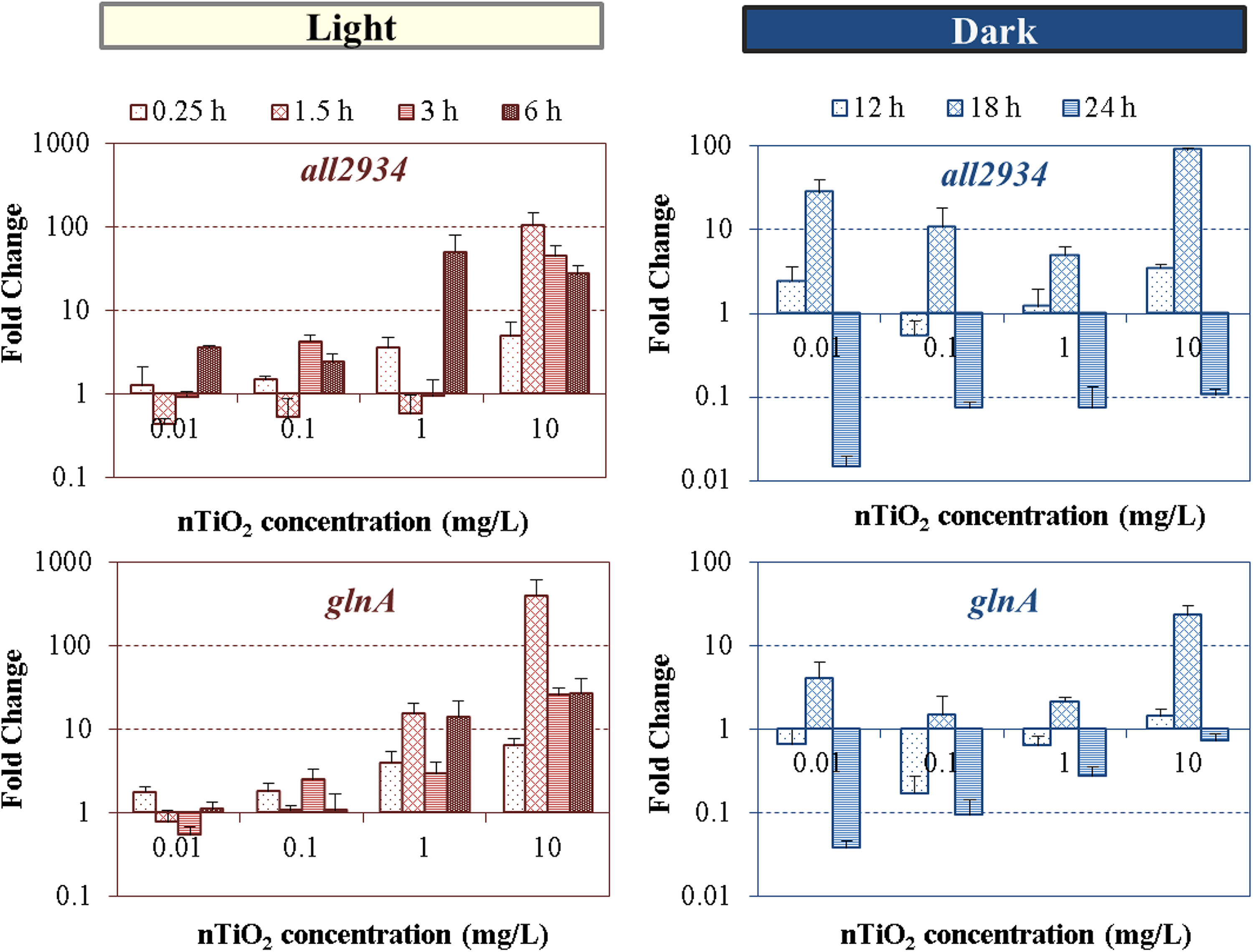
Concentration-dependent levels of expression of all2934 and glnA genes of Anabaena PCC 7120 exposed to nTiO2 concentrations ranged from 0.01 to 10 mg/L at different times of light/dark cycle. Standard deviations are of PCR reactions.
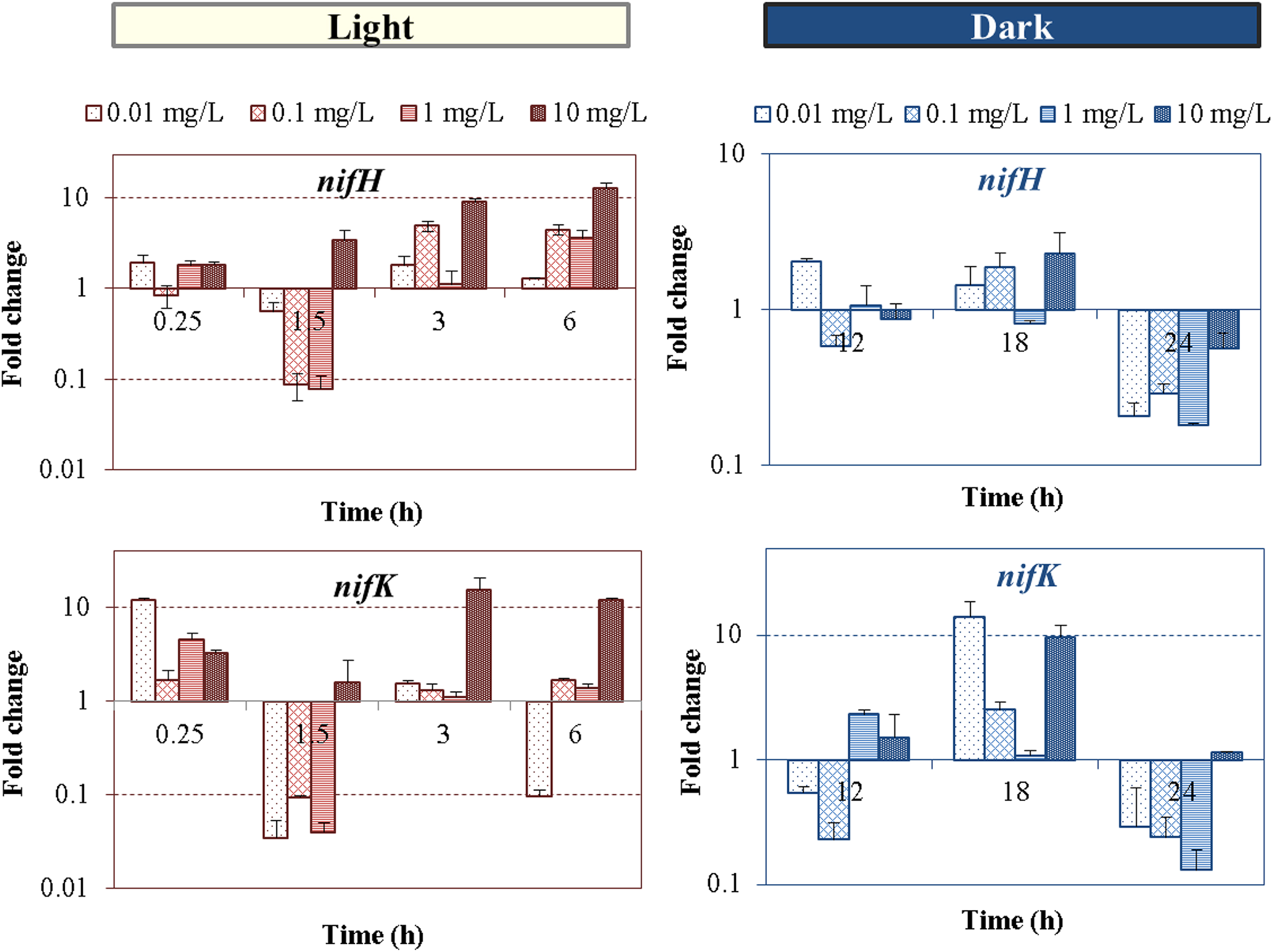
Time-dependent levels of transcriptional expression fold changes of genes involved in nitrogen fixation (nifH and nifK) of Anabaena PCC 7120 exposed to nTiO2 concentrations ranged from 0.01 to 10 mg/L at different times of light/dark cycle. Standard deviations are of PCR reactions.
Impact of nTiO2 on the GS-GOGAT nitrogen assimilation
Heterocysts and vegetative cells of nitrogen-fixing cyanobacteria share the GS-GOGAT metabolic pathway (Fig. 1) to achieve ammonia transfer and assimilation into cellular carbon skeleton (Muro-Pastor and Florencio, 2003). Glutamine synthetase (glnA) and glutaminase (all2934) play essential role in this pathway. Figure 5 shows dose-dependent, yet distinctive patterns in gene expression changes for the light and dark regime, respectively. Pairwise t-test on distinct nTiO2 treatments during the entire 24 hours L-D period showed high similarities between the expression levels of both transcripts (p > 0.27), which is likely related to their sequential interrelationship and mutual dependence within the GS-GOGAT pathway.
In the light cycle, high nTiO2 concentrations induced elevated upregulation levels of glnA and all2934, reaching up to 26 and 27-fold changes at 6 h at 10 mg/L, respectively (Fig. 5). Upregulation of the glnA transcript is modulated at both transcriptional and posttranscriptional level (Galmozzi et al., 2010), dependent on both carbon and nitrogen supply and their ratio. Particularly, conditions of nitrogen shortage warrant higher production of the nitrogen-starvation-signaling molecule 2-OG, in a positive feedback mechanisms (Muro-Pastor et al., 2001), thus promoting the expression of NtcA and PII-regulated genes. Therefore, the activation of differential gene expression involved in nitrogen fixation (i.e., nifH) (Fig. 6) and nitrogen assimilation (i.e., glnA), may attribute to the need for cells to compensate for the shortage of intracellular NH3, and may associate with the decrease in nitrogen fixation activity observed in our previous study (Cherchi and Gu, 2010). This is essential in Anabaena sp. to maintain carbon to nitrogen balance within the filament and therefore, its homeostatic state.
The upregulation of glnA and all2934 genes induced by nTiO2 treatment led to the overall increase of key metabolites (i.e., glutamine and glutamate) in the GS-GOGAT reactions pathway (Fig. 7). With a few exceptions at 0.01 mg/L exposure, nTiO2 led to increased levels of glutamate compared with the untreated control in a dose-dependent manner during the light cycle. The intracellular content of glutamine, however, was almost similar to those in the control (p > 0.05) during the first 3 h of exposure, while significant increases (p < 0.02) of glutamine occurred from 6 to 24 h. The overproduction of intracellular metabolites of glutamine at 24-h exposure (Fig. 7B) might have potentially caused the downregulation of glnA and all2934 genes through mechanisms of cumulative feedback inhibition (Stadtman, 2001), to maintain cells' homeostatic equilibrium in conditions of external perturbations (Muro-Pastor and Florencio, 2003).
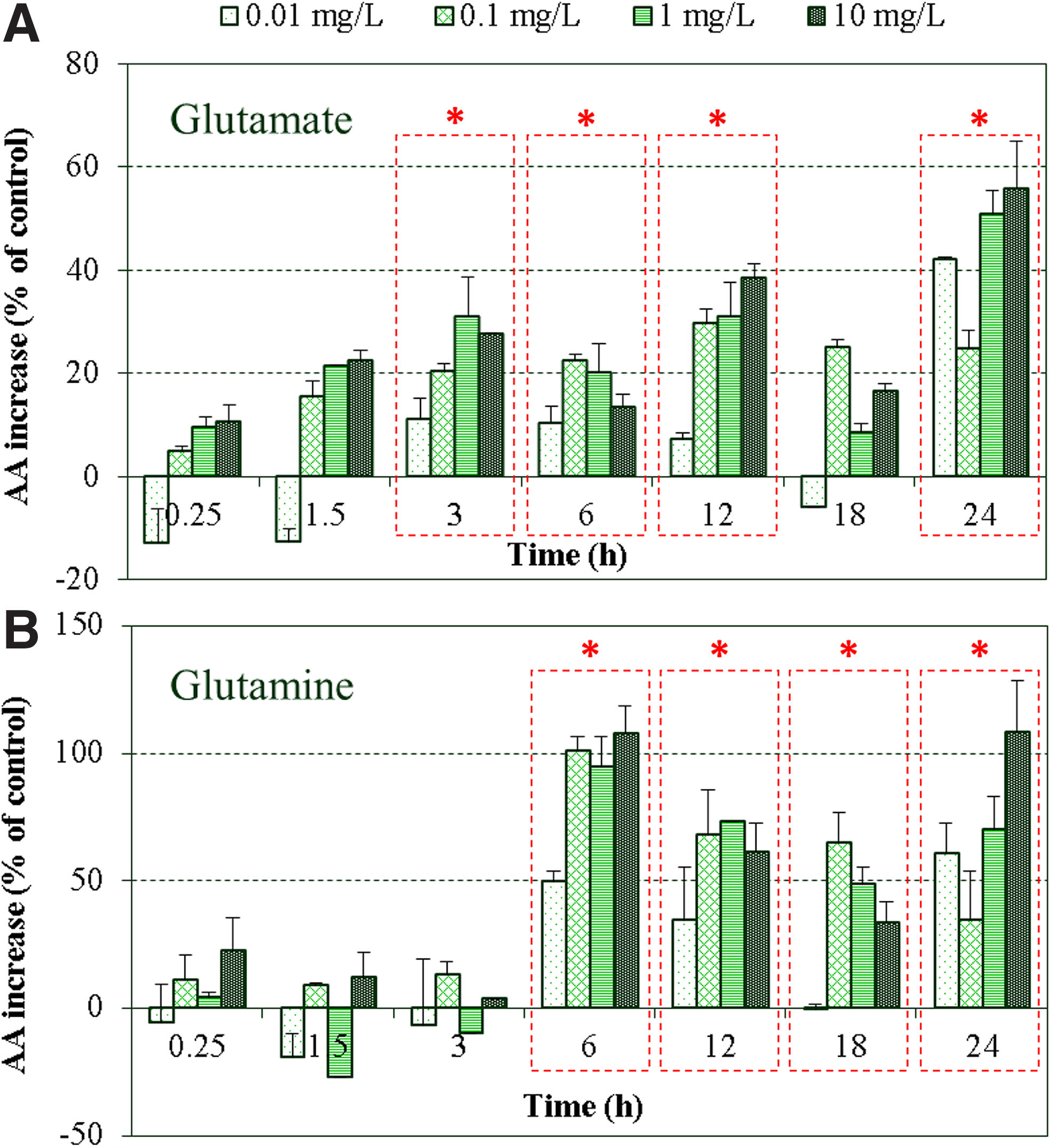
Time-dependent increase of glutamate
Glutamine is uniquely produced by the GS-catalyzed addition of ammonia to glutamate, while different pathways exist for glutamate formation, including the reaction GOGAT catalyzed from glutamine, the catalysis of glutamate dehydrogenase from ammonia and 2-OG, the loss of carbon and nitrogen atoms from the degradation of other amino acids, and the transfer of an amino group to 2-OG through transamination (Berges and Mulholland, 2008). Additionally, of the two molecules of glutamate produced during GS-GOGAT (Fig. 1), only one is reutilized for glutamine production, while the other is expended in vegetative cells for net amino acid synthesis. It was previously reported that under abiotic stress (i.e., metal exposure), plants and algae are known to oversynthetize a number of diverse metabolites, such as proline that primarily derive from glutamate (Sharma and Dietz, 2006). Therefore, the involvement of glutamate in other pathways might justify the different trends and levels of glutamine and glutamate observed, although both are key players of equal importance in the nitrogen assimilation pathway. As key players in the assimilation of ammonia into carbon skeleton, the increased levels of amino acids involved in the GS-GOGAT pathway clearly demonstrated the alteration in C/N equilibriums induced by nTiO2 exposure, indicating the possible impairment of intracellular nutrient balance and homeostatic stability.
Responses of global nitrogen metabolism regulators to nTiO2
The regulation of intracellular carbon and nitrogen balance is crucial for cyanobacteria to meet their metabolic requirements under regular growth conditions and to seek survival during environmental perturbations (Wegener et al., 2010). The global regulator, NtcA protein, integrates signals from carbon and nitrogen metabolisms, and thus mediates the cellular nitrogen status under the control of a large set of marker genes (Muro-Pastor et al., 2001). The binding of NtcA protein to its target promoters is triggered by the intracellular levels of 2-OG, the carbon skeleton involved in the assimilation of nitrogen. The signal transducer PII protein (encoded by the glnB gene) can sense the carbon/nitrogen status of the cell through the synergistic binding of effector molecules (ATP and 2-OG) (Fokina et al., 2010), and regulate the cellular C/N balance through phosphorylation (Watzer et al., 2019). The protein NtcA and the PII regulator are functionally interdependent, since they are mechanistically linked by the transcriptional coactivator PipX protein (Espinosa et al., 2006). The glnB gene is transcriptionally activated by NtcA and the PII protein is required for the activation of NtcA-regulated genes (Tolonen et al., 2006).
Dose-dependent differential gene expression patterns of the two global nitrogen regulators compared with control without nTiO2 dosage were illustrated in Fig. 8. At the highest concentration of 10 mg/L, significant transcriptional changes as high as 20-fold for ntcA and 7-fold for glnB were observed at 6-h exposure in light cycle, and 5.5-fold (ntcA) and 5-fold (glnB) changes at 18-h exposure during dark. These results clearly demonstrated the impact of nTiO2 exposure on subcellular nitrogen regulation. Recently, the knowledge about the role of NtcA has been broadened to the regulation of genes involved in carbon metabolism, photosynthesis, and stress responses (Chen et al., 2011). This indicates that other factors, rather than only nitrogen perturbations, can affect ntcA and glnB expression. A previous study has also demonstrated the importance of maintaining internal reducing conditions and optimal redox status to favor the DNA-binding capacity of NtcA to target promoters in Anabaena sp. (Jiang et al., 1997). Therefore, the expression patterns observed during dark and light cycles are possibly affected by the unbalanced oxidative/reducing conditions caused by intracellular reactive oxygen species formation upon nTiO2 treatment (Cherchi et al., 2011), as well as the potential changes in the levels of reducing power linked to the alteration of cell photosynthetic, nitrogen fixation potential (Cherchi and Gu, 2010), and electron transport (Alfonso et al., 2001). Note that the changes in gene regulators do not fully reflect the changes at translational and post-translational level; nevertheless, our results evidenced the active regulation of nitrogen metabolism in cyanobacteria through the transcriptional-level response of two global nitrogen regulator genes (ntcA and glnB) upon the exposure to sublethal environmentally relevant concentrations of nTiO2.
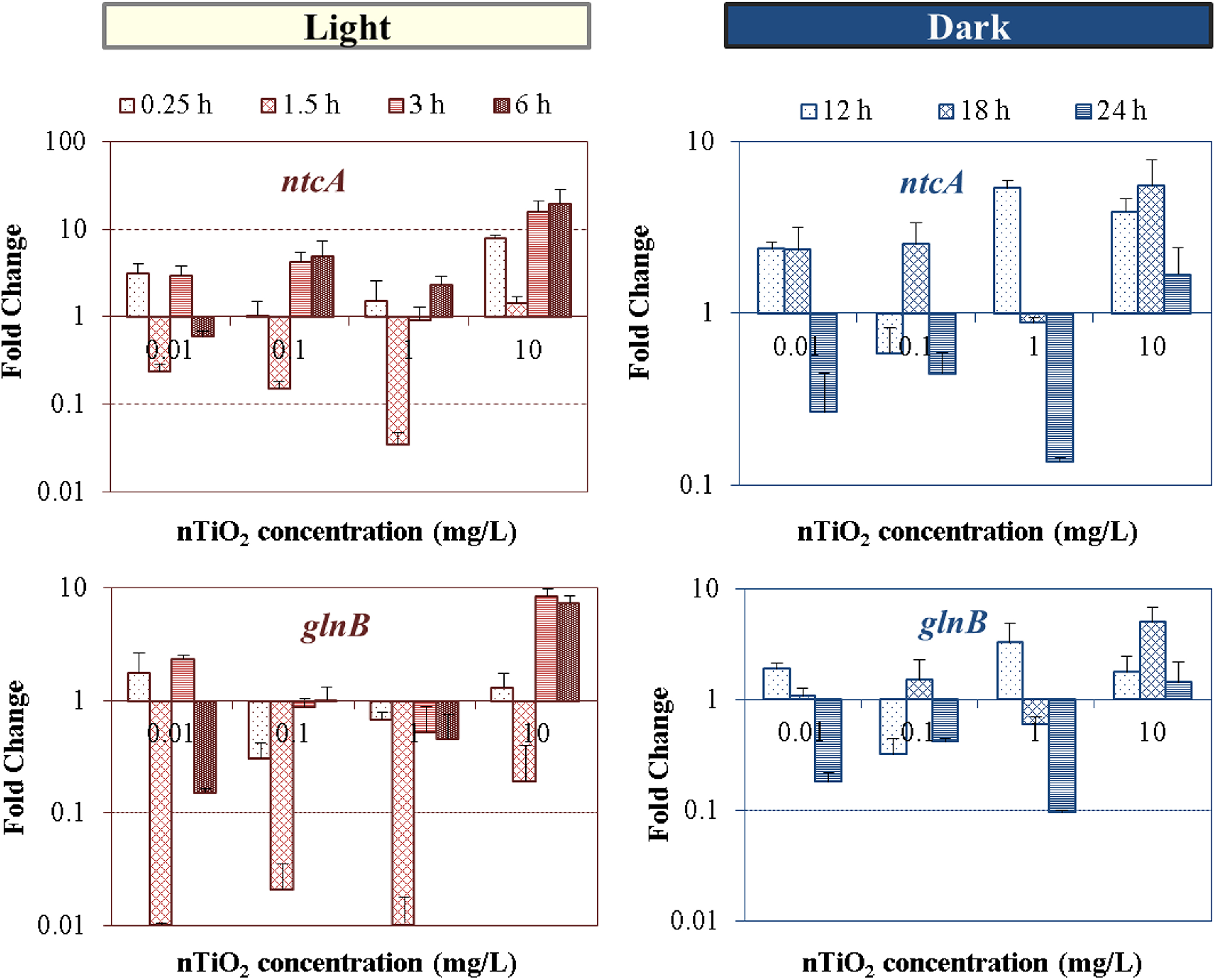
Concentration-dependent alteration in the fold change expression (relative to control) of ntcA and glnB genes of Anabaena PCC 7120, after exposure to nTiO2 concentrations ranged from 0.01 to 10 mg/L at different time points during the 24-h light/dark cycle.
Impact of nTiO2 on nitrogen fixation-related transcriptional activity
The two-component complex of nitrogenase catalyzes the fixation of dinitrogen through the expression of ∼20 different nif genes (Howard and Rees, 1996). Among all, the nifH gene encodes for dinitrogenase reductase, which is the obligate electron donor to dinitrogenase during the nitrogenase turnover, whereas nifK encodes for the β subunit of dinitrogenase.
Exposure to nTiO2 induced changes on the two nif gene expression during both light and dark cycles (Fig. 6). Alterations in the gene expression levels of both nifH and nifK genes were observed, which exhibited a dose-dependent trend, particularly in the light cycle. Nano-TiO2 at higher doses (10 mg/L) clearly led to significant response of the nif genes, indicating the perturbation of nitrogen fixation activity, which in turn may affect the intracellular C/N balance. During the dark cycle, expression levels of the two nif gene appeared not to be dose dependent, but dose specific, with upregulation mainly occurring in the middle of dark cycle (18 h). Moreover, downregulation of the majority of tested marker genes was mainly observed at 1.5, 12, and 24 h after nTiO2 dosing (Fig. 2), which correspond to the time points close to L-D and D-L switches. Differences in N fixation activities between light and dark cycles are expected since the expression and activity of nitrogenase require a high portion of the cellular energy pool (Maner and Oelze, 1999) and are tightly controlled by reducing power (NADPH) and ATP supply that are light dependent. Lower nitrogen fixation rates are known to occur during dark conditions when PSI-mediated photophosphorylation is lacking, and ATP is only provided by the oxidative phosphorylation. A previous study in our group showed a decrease in nitrogenase activity and nitrogen fixation inhibition in Anabaena variabilis at comparable and higher doses of nTiO2. The definite link between nif genes, nitrogenase, and nitrogen fixation is hard to draw because the control and regulation of nitrogen fixation in cyanobacteria occurs at transcriptional, posttranscriptional, and translational level, as previously reported (Severin and Stal, 2010). Additionally, the nitrogenase components are not immediately competent for nitrogen fixation after synthesis, and they acquire catalytic activation afterward by the actions of a number of nif and non-nif gene products (Rubio and Ludden, 2005).
The metabolic connection of the obligate electron donor role of the Fe protein dinitrogenase reductase (nifH) with the MoFe protein component of dinitrogenase (nifK) was not evident for nTiO2 concentrations in the range of 10–1,000 μg/L (Pearson coefficient <0.52), showing that low nTiO2 doses have the potential to hamper the relative nifH and nifK transcription, thus the structural stability of nitrogenase subunits. Additionally, if nTiO2 affects the efficiency of their translation, the decoupled accumulations of these two genes may represent the cell potential to rebalance the relative levels of their transcript and reoptimize their functional linkage. For example, different levels of nifH and nifK messenger RNA (mRNA) were accumulated in Azotobacter vinelandii reaching steady-state nitrogen fixing conditions, to maintain high Fe protein-to-MoFe protein ratios required for an optimal nitrogenase catalytic activity (Hamilton et al., 2011). On the other hand, high correlation (Pearson coefficient >0.75) between the two genes in their temporal L-D responses at higher doses (10 mg/L) was observed, suggesting their concomitant up- and downregulation when the toxicity effect of nTiO2 became dominant.
Conclusions
Thus far, the report of molecular and genetic response to NM exposure of ecologically relevant organisms and, particularly of photoautotrophic species, has been scarce. The present work evidenced the potential of nTiO2 to perturb multiple cellular pathways involved in the nitrogen status of nitrogen-fixing organisms and to alter their complex gene regulation systems. Both the circadian and fluctuating patterns of cyanobacterial metabolism and nTiO2 intrinsic properties distinctively govern the toxicity response during light and dark exposure to nTiO2.
From our results, cell response to nTiO2-induced stress involves increased levels of proteins, which was reflected in the observed increase of amino acid content. Additionally, in consequence of the potentially decreased nitrogen fixation activity, cells might have sensed lower intracellular levels of ammonia, and thus limited availability of nitrogen. Immediately, cells activate nitrogenase to provide more nitrogen for assimilation, with consequent activation of genes involved in the GS-GOGAT, which showed dose dependency. In parallel to the immediate requirement for proteins, cells tend to prepare for upcoming periods of nutrient shortage and stress by accumulating nitrogen in form of storage products (CGPs), particularly at night, when the lower metabolic requirements for proteins and nTiO2 toxicity facilitate nitrogen diversion into CGPs. These specific responses are clearly modulated by the expression of the two global nitrogen regulators, implying that the signaling networks linking not only to cell nitrogen status, but more broadly, to carbon and nitrogen balance, are affected by nTiO2 exposure.
Ecosystem functions are controlled by flows of matter and energy, whose equilibrium is prone to be altered by anthropogenic materials and pollutants released in the environment. The findings of this study warn the potential of NMs to affect ecological nutrient balances and metabolic capabilities in organisms, such as cyanobacteria, which significantly contribute to the biologically available nitrogen on earth. This will broadly lead to adverse impacts on ecosystems' stoichiometry, fitness and organisms' growth rates, and will indirectly affect biological productivity and the dynamics of communities in food webs.
Footnotes
Acknowledgment
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Science Foundation (NSF) Nanoscale Science and Engineering Center (NSEC) for High-Rate Nanomanufacturing (grant No. 0425826) and the NSF, CBET-1437257, and CBET-1829754.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.