
Abstract
This study investigated the rapid start-up of partial nitrification through different inhibitor addition, including hydroxylamine (NH2OH), potassium chlorate (KClO3), and formic acid (HCOOH). Then, the performances of the three inhibitors on the rapid recovery of partial nitrification once increasing the influent ammonium sharply were also compared. Finally, real-time aeration control strategy was adopted to maintain partial nitrification during a period. In the present work, batch experiments were used to confirm the optimum dosage of each inhibitor in a similar adding range first. The results showed that nitrite-oxidizing bacteria (NOB) were more vulnerable to NH2OH and KClO3 than HCOOH. Second, the rapid start-up of partial nitrification in a sequencing batch reactor was evaluated. It finished in 22 days without any inhibitor. However, only 12 days were needed when adding 0.15 mM NH2OH. It used 14 days when adding 0.5 mM KClO3 and 17 days under 0.5 mM HCOOH addition, respectively. The partial nitrification, which was destroyed by the high influent ammonium, was easily and rapidly recovered using the inhibitors. The nitrite accumulation rate was still more than 80% using the real-time aeration control strategy when ceasing the inhibitor addition. Furthermore, high-throughput sequencing analysis revealed that the main genera of NOB, Nitrospira, was suppressed by the three inhibitors; however, a difference in bacterial community structure was found due to the different influence mechanisms. Overall, inhibitor addition and real-time aeration control is an excellent strategy for establishing, recovering, and maintaining effective partial nitrification.
Introduction
The conventional biological nitrogen removal technology is based on the nitrification and denitrification processes, which require high energy for aeration and external biodegradable chemical oxygen demand for denitrification (Gabarró et al., 2012; Ma et al., 2016; Wang et al., 2019). Considering the processing cost, many novel processes such as the partial nitrification-anammox (PNA) process have recently been developed (Qian et al., 2013; Miao et al., 2018). The PNA process can reduce at least 60% of energy for aeration and save up to 100% of the organic matter requirement. However, the anammox process requires both ammonium and nitrite in a molar ratio of 1:1.32 as reaction substrates (Qian et al., 2013). Unfortunately in most wastewater, ammonium is the main nitrogen compound, and therefore, it needs a pretreatment for the achievement of the suitable molar ratio. Thus, partial nitrification, which converts 50–57% of influent ammonium to nitrite and prevents the production of nitrate, plays an important role in the PNA process (Wang et al., 2018; Yang et al., 2018).
However, the application of partial nitrification is rarely observed in conventional wastewater treatment processes. This is because nitrite-oxidizing bacteria (NOB) activity is usually similar or higher than ammonia-oxidizing bacteria (AOB) activity. As a result, ammonium can be easily oxidized to nitrate completely not to nitrite (Peng and Zhu, 2006; Miao et al., 2018). Thus, the key to start-up and maintain partial nitrification is to accumulate AOB and inhibit or washout NOB (Lackner et al., 2014; Li et al., 2019a). There is a growing effort toward addressing the problem, for instance, low dissolved oxygen (DO) (Chuang et al., 2007; Blackburne et al., 2008; Guo et al., 2009), high free ammonia (FA) and free nitrous acid (FNA) (Guo et al., 2009a; Park and Bae, 2009; Qiao et al., 2010, 2013), low sludge retention time (SRT) (Hellinga et al., 1998; Guo et al., 2009b, 2009c), and real-time control (Regmi et al., 2014) have been reported to achieve partial nitrification successfully. However, there are some drawbacks to these strategies. For example, Hulle et al. (2010) found that low DO effectively inhibited the NOB activity, but it also limited the reaction rate of partial nitrification. Moreover, low DO caused the excessive growth of filamentous organisms, leading to the unqualified sedimentation performance of sludge (Bao et al., 2009). Wang et al. (2014, 2017) illustrated a strategy for achieving partial nitrification, which involves using high FA and FNA as inhibitors. However, they also found that the strategy increased the input of additional nitrogen load, and the cumulative nitrite concentration in the common wastewater treatment would seldom reach the NOB inhibition threshold. Moreover, Bae et al. (2002) and Duan et al. (2019) reported that partial nitrification could deteriorate during long-term operation in the case of high FA. Ma et al. (2017) observed that NOB showed capable of tolerating increasing levels of FNA, causing the failure of the FNA established partial nitrification system. Yuan and Oleszkiewicz (2011) indicated that low SRT led to the low net biomass production of functional bacteria because of continuous biomass washout. In a word, achieving stable partial nitrification is a challenge through the normal control parameters. Effective and economical strategies for the start-up and maintenance of partial nitrification are of significance for the application of novel biological nitrogen removal processes.
Recently, the addition of some inhibitors for realizing partial nitrification has received much attention. As an intermediate of nitrification, hydroxylamine (NH2OH) showed an irreversible inhibitory effect on the growth of NOB but not AOB (Van et al., 2008; Xu et al., 2012). Li et al. (2019b) investigated the partial nitrification for domestic wastewater through the short-term addition of 5 mg/L NH2OH. The results showed that the process was quickly established in 5 days. Hynes and Knowles (1983) found that potassium chlorate (KClO3) selectively inhibited NOB but not AOB. Xu et al. (2011) reported that the partial nitrification was successfully achieved with the addition of 5 mM KClO3 in the aerobic granule system. Formic acid (HCOOH) is an environmental-friendly organic acid and can hinder full nitrification (Eilersen et al., 1994). Wang et al. (2020a) evaluated the feasibility of achieving stable partial nitrification by adding 30 mM HCOOH. During 27 days of operation, the nitrite accumulation rate (NAR) was rapidly raised and maintained at ∼90%. In brief, the above inhibitors have been successfully proposed, while there is no consensus on the most effective inhibitor today, making it necessary to develop an effective and environmental-friendly inhibitor, which was easy to be decomposed by microorganisms and would not bring any toxicity to the environment or the bacteria in the subsequent wastewater treatment processes. It is interesting to compare the performances of different inhibitors in the start-up of partial nitrification when adding a similar dosage. Indeed, the inhibitor addition occurred in intermittent and continuous modes (Xu et al., 2011; Li et al., 2019a; Wang et al., 2020b). Until now, limited research has been done on the low dose concentration, as well as intermittent modes, of the three compounds on the recovery of partial nitrification once destroyed due to the changes of environmental conditions. Thus, data are essential in evaluating the feasibility and application of the inhibitor for achieving stable partial nitrification.
Previous studies found that the deterioration of partial nitrification appeared without continuous inhibitor addition. For example, Wang et al. (2015) illustrated that the nitrate buildup occurred within just 11 days after NH2OH dosing was stopped since the microorganism gradually acclimated to NH2OH inhibition. It is necessary to develop an efficient and cost-effective method to maintain partial nitrification once ceasing the addition of inhibitor. Real-time aeration control has been proven to be a feasible strategy to maintain stable partial nitrification of low strength domestic wastewater (Blackburne et al., 2008). They also pointed out the drawback of real-time aeration control: it usually needed a long period to initiate partial nitrification. Yang et al. (2007) reported that 76 days were required to achieve partial nitrification when using real-time aeration control strategy. Interestingly, the addition of appropriate inhibitors can enhance the start-up of partial nitrification rapidly, and the real-time aeration control can maintain partial nitrification steadily. The combination of these strategies seems to be a convenient and economical method for the application of partial nitrification.
The aims of this study were to (1) compare the performances for achieving partial nitrification by adding different inhibitors in a similar dosage range, including NH2OH, KClO3, and HCOOH; (2) study the feasibility of using the three inhibitors for recovering partial nitrification once destroyed due to the increase of influent ammonium; and (3) explore the potential of maintaining partial nitrification by real-time aeration control when ceasing the inhibitor addition. Batch experiments were established to explore the optimal conditions for inhibitors. Furthermore, the nitrogen removal performances of sequencing batch reactor (SBR) under the optimal inhibitor addition were evaluated during a long-term experiment. Then, variation in the nitrifying microbial community was assessed through the high-throughput sequencing analysis.
Materials and Methods
Seed sludge and synthetic wastewater
The activated sludge was taken from the Tuandao wastewater treatment plant in Qingdao, China, which used the Anaerobic–Anoxic–Oxic (A/A/O) biological process to treat the domestic wastewater. The concentrations of mixed liquor suspended solids (MLSS) and mixed liquor volatile suspended solid (MLVSS) in reactors after inoculation were ∼3,000 and 2,000 mg/L, respectively. NH2OH, KClO3, HCOOH, and other chemicals were reagent grade (AR) and supplied from Sinopharm Chemical Reagent Co., Ltd.
The synthetic wastewater used in the experiment was modified from a previous study (Wang et al., 2017), which comprised ∼229.25 mg/L of NH4Cl (60 mg/L
Batch experiment
Different dosage of NH2OH, KClO3, and HCOOH was used in the experiments, which were carried out in 500 mL glass bottles. According to the previous research (Li et al., 2019a, 2019b), the dosage of NH2OH was chosen from 0 to 0.3 mM. Previous research (Lees and Simpson, 1957) indicated that 1 to 10 mM KClO3 blocked the oxidation of nitrite completely. Xu et al. (2011) used 1 mM KClO3 for the selective inhibition of nitrite oxidation in the SBR. To compare the effect of the three inhibitors in the case of a similar dosage, the adding range of KClO3 and HCOOH was both from 0 to 1 mM in the present work. During operation, the initial ammonium was 60 mg/L, and the initial pH was adjusted to 8.0. The DO concentration was 2 mg/L. The temperature was maintained at 25°C using a thermostat water bath. After 15 cycles, the samples and sludge were collected at predetermined intervals and analyzed through the methods mentioned in Analytical methods.
Long-term experiment
The long-term experiments were conducted in four SBRs with a working volume of 2 L. It was divided into three phases. In phase I (day 0–22), the inhibitor was added every day and the influent ammonium was about 60 mg/L. The operational cycle of the SBRs consisted of 10 h, and the aeration time was 8 h. In phase II (days 23–35), the inhibitor was added every three days, and the influent ammonium increased to 150 mg/L. The operational cycle was also 10 h, while the aeration time was reduced to 6 h. In phase III (days 36–40), the addition of inhibitor was ceased. The aeration time was determined by a real-time aeration control strategy. The dosage of each inhibitor used in the experiments was the optimum concentration determined in Batch experiment. The pH of the SBRs was between 7.9 and 8.0, and DO was controlled at about 2 mg/L during the aeration. All reactors were operated at a temperature of 25°C, and no sludge was discharged during the whole process.
Analytical measurements
The parameters, including MLSS, MLVSS,
High-throughput sequencing analysis
The sludge samples collected in the reactors were freeze-dried by a lyophilizer (Labconco Co., Free Zone, USA). DNA was extracted from 0.10 to 0.20 g dry sludge samples using FastDNA™ SPIN Kits for Soil (M5635-02; Omega, USA), and the DNA concentration was determined using a Qubit2.0 DNA Detection Kit (Q10212; Life, USA). The universal bacterial 16SrRNA genes for total bacteria were amplified. The standard curves were generated in duplicate using serial decimal dilutions of plasmid DNA and adopted when their amplification efficiency was between 90% and 110% and correlation coefficient above 0.99.
Analytical methods
The ammonium removal efficiency (ARE) and NAR were estimated with Equations (1) and (2) (Miao et al., 2018; Li et al., 2019b; Wang et al., 2020a).

Where
Previous studies used the biomass-specific ammonium and nitrite oxidation rates to evaluate the activities of AOB and NOB, respectively (Wang et al., 2017; Cui et al., 2020). In the present work, the biomass-specific ammonium and nitrite oxidation rates were calculated using the following formula [Eqs. (3) and (4)]. All experiments were performed in duplicate. One-way analysis of variance (ANOVA) was used to assess the significance of results with p < 0.05.

Results and Discussion
Effect of different inhibitors in batch experiments
The identical reactors were used to select the optimum inhibitor dosage for establishing efficient partial nitrification. In all reactors, steady and complete nitrification was obtained initially with a NAR of less than 1%. Following this, different volumes of inhibitors (as shown in the Batch experiments) were injected into each of the reactors per cycle.
The effect of NH2OH is shown in Fig. 1a and b. After 15 cycles, ARE without NH2OH addition was 97.87% within 10 h, while it increased to 99.83% when adding 0.15 mM NH2OH. However, it decreased to 82.98% when 0.3 mM NH2OH was used (Fig. 1a). For NAR, it was 38% with no inhibitor, suggesting the accumulation of nitrite. Suthersan and Ganczarczyk (1986) also found that when the pH value of wastewater was controlled above 8.0, the accumulation of nitrite could be realized. When adding 0.15 mM NH2OH, the maximum value of NAR was 81.5%, while the value decreased to 57.8% as increasing the dosage to 1 mM. The results indicated that NH2OH could not only enhance the removal of
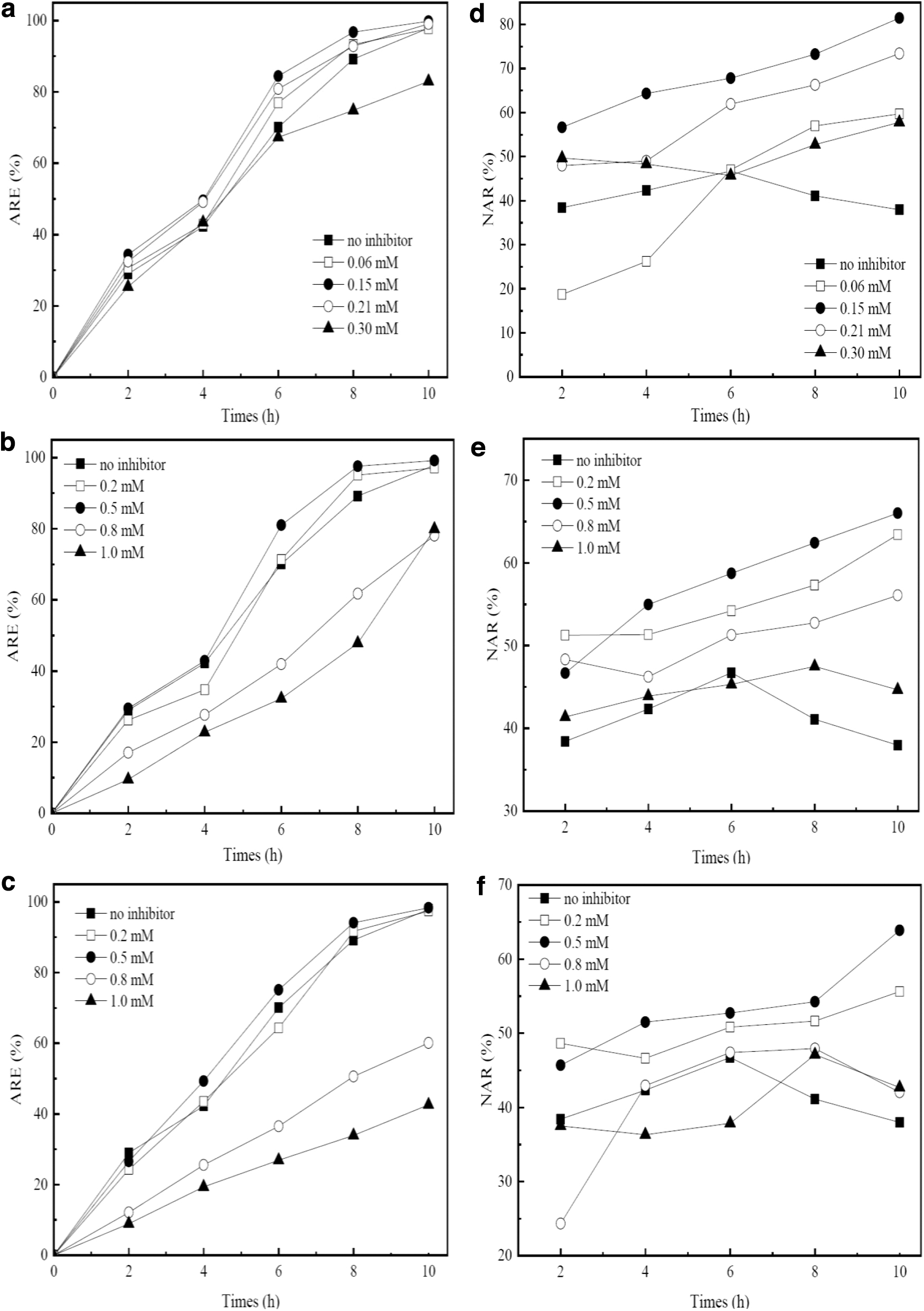
The results of ARE and NAR at various concentrations of inhibitors:
As shown in Fig. 1c, ARE was 97.1, 99.2, 78.1, and 78.9% with the addition of 0.2, 0.5, 0.8, and 1 mM KClO3, respectively. NAR increased from 38% to 66% by increasing the dosage to 0.5 mM, while it decreased to 44.65% when adding 1 mM KClO3 (Fig. 1e). The result was consistent with that obtained by Lees and Simpson (1957), who indicated that low dosage KClO3 had no effect on AOB, but could block the oxidation of nitrite. We also found that high KClO3 would suppress the performance of
The results of exploring the optimal dosage of HCOOH are shown in Fig. 1c–f. ARE increased to 98.36% when adding 0.5 mM HCOOH, while it decreased to 42.55% with 1 mM addition. NAR increased from 38% to 63.9% when adding HCOOH from 0 to 0.5 mM. However, as continuously increasing the dosage to 1 mM, it decreased sharply to 42.7%. We measured the pH of the system and found it changed to acidic by adding more HCOOH. Law et al. (2011) indicated that low pH inhibited the activity of nitrifying bacteria obviously, resulting in the reduction of both
The activity of AOB and NOB in the four systems after 15 cycles was compared in the present work. As shown in Supplementary Fig. S1, the activity of AOB without any inhibitor was 2.934
Long-term operating performance in the presence of different inhibitors
A long-term experiment of SBR in the presence of different inhibitors was divided into three phases. In phase I (0–22 days), the ammonium concentration of effluent was 13.5 mg/L after 22 days without any inhibitor. A measure of 25.4 mg/L nitrite and 25.5 mg/L nitrate was also detected in the effluent (Fig. 2a). In addition, Fig. 3a showed that ARE reached above 81.6% throughout the acclimation period. Simultaneously, NAR increased gradually and reached above 42.3%, suggesting the start-up of partial nitrification (Fig. 3b). When adding 0.15 mM NH2OH, 11.8 mg/L of ammonium, 29.9 mg/L of nitrite, and 15.5 mg/L of nitrate were detected in the effluent on day 12 (Fig. 2b). Meanwhile, ARE was 80.4% and NAR was 61.9%, respectively. When 0.5 mM KClO3 was added for 14 days, ARE was 80.7% and NAR was 52.85%. The start-up needed 17 days for 0.5 mM HCOOH addition. It was 80% of ARE and 53.27% of NAR, respectively. These results illustrated that the addition of an inhibitor could reduce the start-up time of partial nitrification significantly.
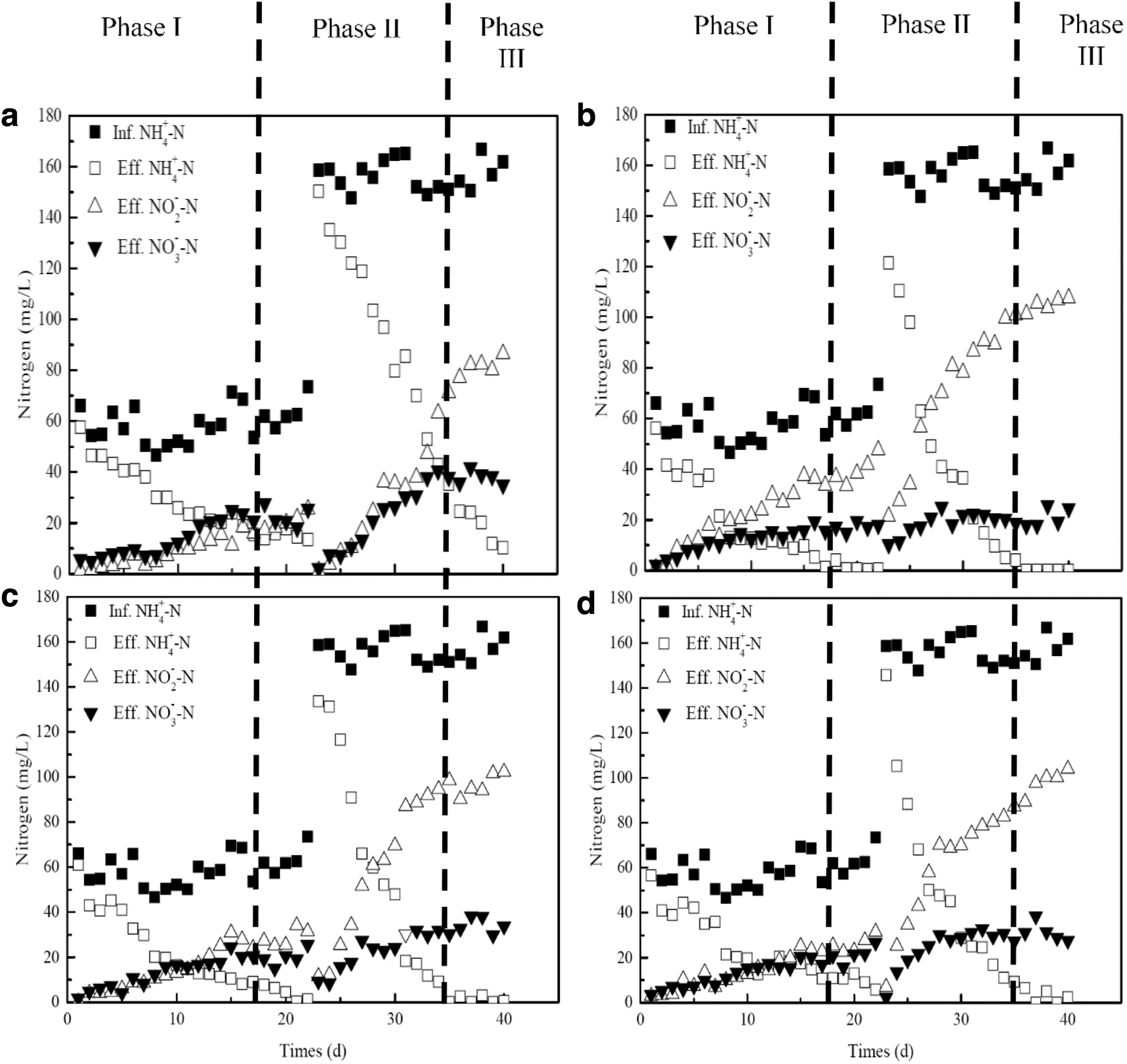
Profiles of ammonium, nitrite, and nitrate concentrations during the experimental. The different phases were indicated by lines:

The results of
In phase II (23–35 days), the influent ammonium increased to around 150 mg/L, and the aeration time was reduced to 6 h. The partial nitrification was destroyed once increasing the influent ammonium initially. ARE reduced to 5.23% (without any inhibitor), 23.4% (NH2OH), 15.8% (KClO3), and 8.13% (HCOOH), respectively. For NAR, it was 13.25%, 57.41%, 46%, and 54.26%, respectively (Fig. 3). However, the results showed that a long-term stimulation by inhibitor improved the steady operation of partial nitrification. Furthermore, the ammonium was removed thoroughly, and NAR returned to 68.7% with the NH2OH addition on day 35. It is clear that the usage of inhibitor was to the advantage of the recovery of partial nitrification in a short time.
However, Li et al. (2019a) found that the deterioration of partial nitrification appeared during a long-term operation without continuous NH2OH addition. Real-time aeration control strategy is developed to effectively avoid excessive aeration, which is favorable to the maintenance of stable partial nitrification (Qian et al., 2017). In phase III (36–40 days), we ceased the inhibitor addition and the aeration time was controlled in real time, which was determined based on the performances of ARE and NAR in a cycle. As shown in Fig. 4, ARE in the absence of inhibitor was 100% after 300 min, while it reached 100% within 180 min under the NH2OH addition. It needed 240 min to remove ammonium thoroughly for both KClO3 and HCOOH addition. Meanwhile, NAR was only 37.25% without any inhibitor, while it was 51.9% for NH2OH addition, 50.93% for KClO3 addition, and 50.9% for HCOOH addition, indicating the occurrence of partial nitrification. As a result, the aeration time was 300 min for no inhibitor addition, while it was 180, 240, and 240 min for the three inhibitors, respectively. Nitrogen variations under real-time control are also shown in Fig. 2. Ammonium level was reduced, and nitrite was continuously accumulated. ARE was more than 90% (Fig. 3a), and a relatively stable partial nitrification performance was obtained to more than 60% (Fig. 3b) under the three inhibitor addition.
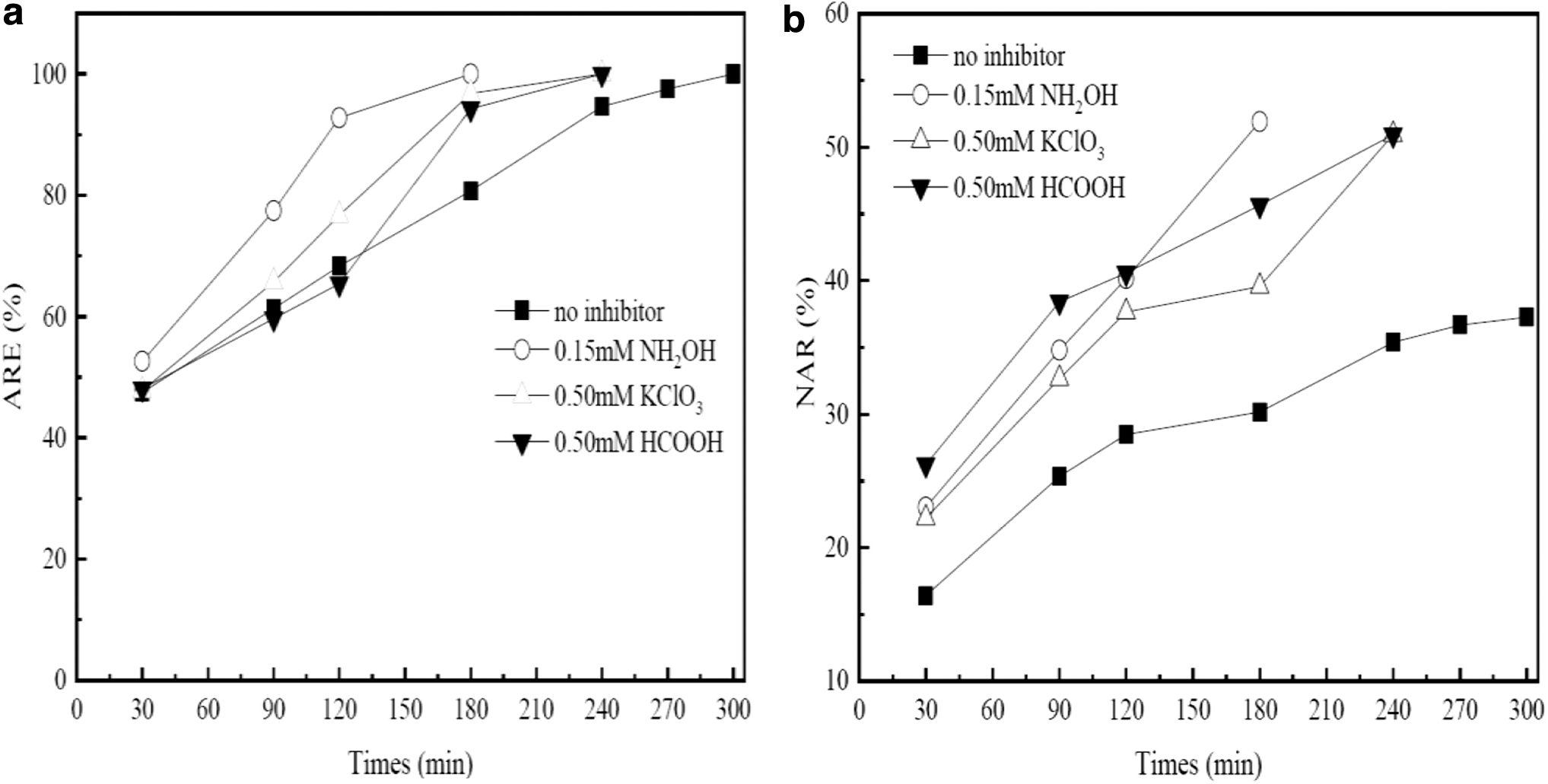
The results of
Figure 5 showed the variations of nitrifying bacteria activities during the experimental period. As shown in Fig. 5a, the initial activity of AOB activity was 0.38
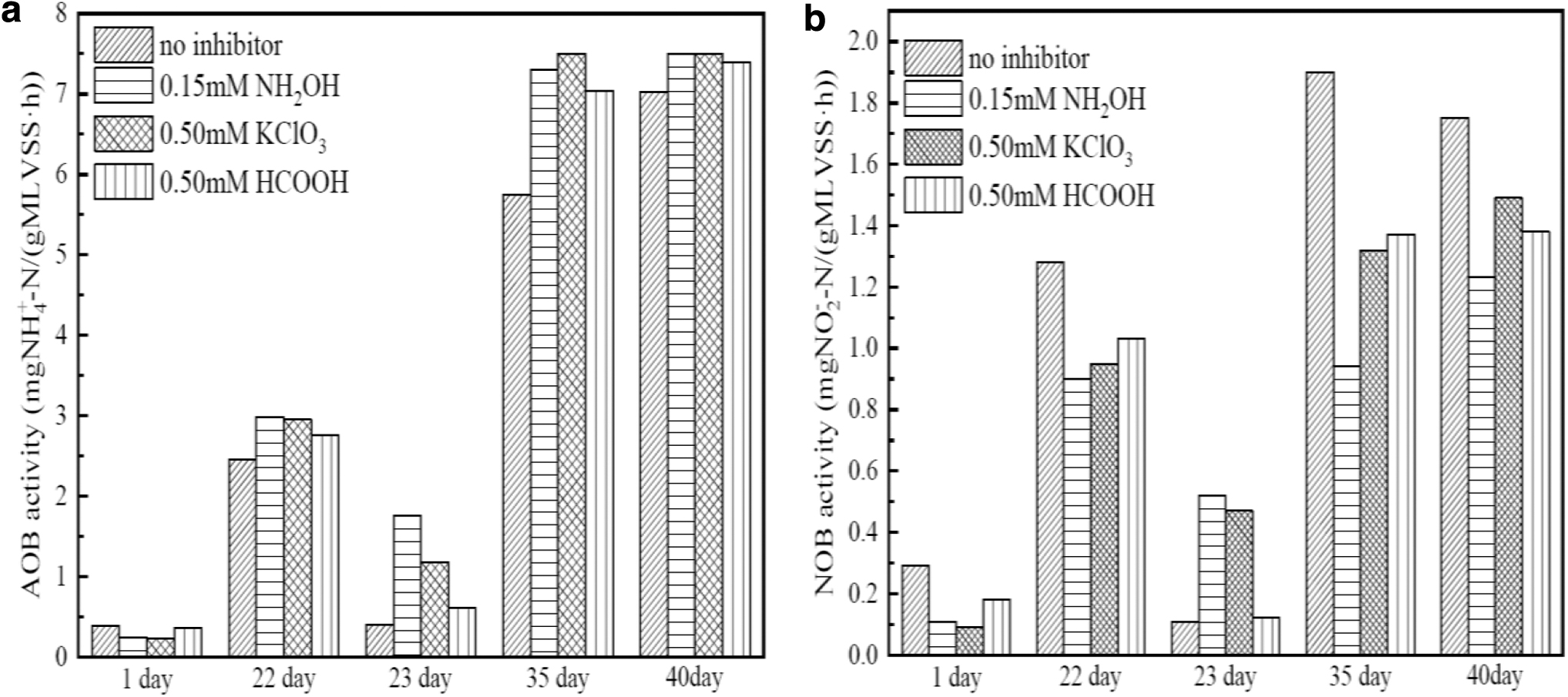
Variations of nitrifying bacteria activities
Effect of inhibitor on microbial community
To understand the changes in active bacterial diversity and structure of functional bacteria, high-throughput sequencing was used to test the activated sludge samples of day 35. The Good's coverage values were above 98% in all samples, suggesting that the microbial sequencing matched the requirements of analysis. The difference analysis of operational taxonomic units (OTUs) was presented through the Venn diagram (Fig. 6). A total of 3,496 OTUs were detected, whereas only 26.2% (916 OTUs) species were shared by the four systems, indicating that the active microbial community structure was different in each system. Figure 7a showed the main genus with relative abundances of more than 1%. The results indicated that the active microbial community without any inhibitor was similar to that with HCOOH addition, while it was quite different from that under NH2OH or KClO3 addition. Comamonas, a typical bacillus, was found much more in the NH2OH or KClO3 system than in the HCOOH system. However, plenty of Rhodobacteraceae, which are common heterotrophic nitrifying bacteria, was found in the HCOOH system, suggesting that the effect of HCOOH acted as a carbon source. Figure 7b showed the AOB and NOB activity levels. Nitrosomonas was the main genera of AOB, and the dominant NOB genus Nitrospira was detected. The relative abundance of AOB in the absence of an inhibitor was 1.58%, while it was 3.46% (0.15 mM NH2OH), 2.8% (0.5 mM KClO3), and 2.43% (0.5 mM HCOOH), respectively. However, NOB represented 1.03%, 0.57%, 0.4%, and 0.78% of the microbial communities, respectively. The results indicated that not only AOB activity, the AOB population also increased when using the inhibitors, while the NOB population was also inhibited by the inhibitors.
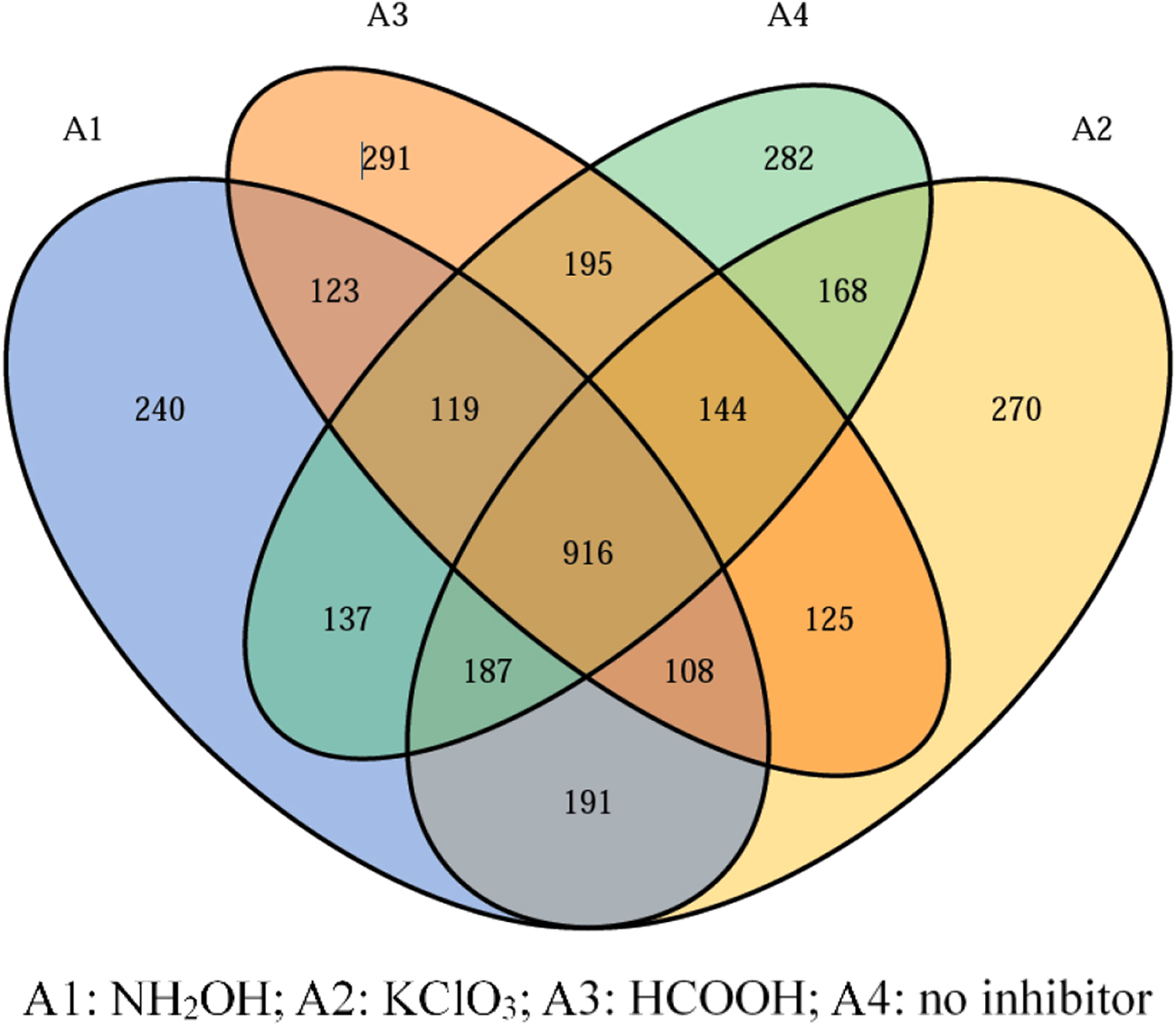
Shifts in community structure as illustrated by Venn.
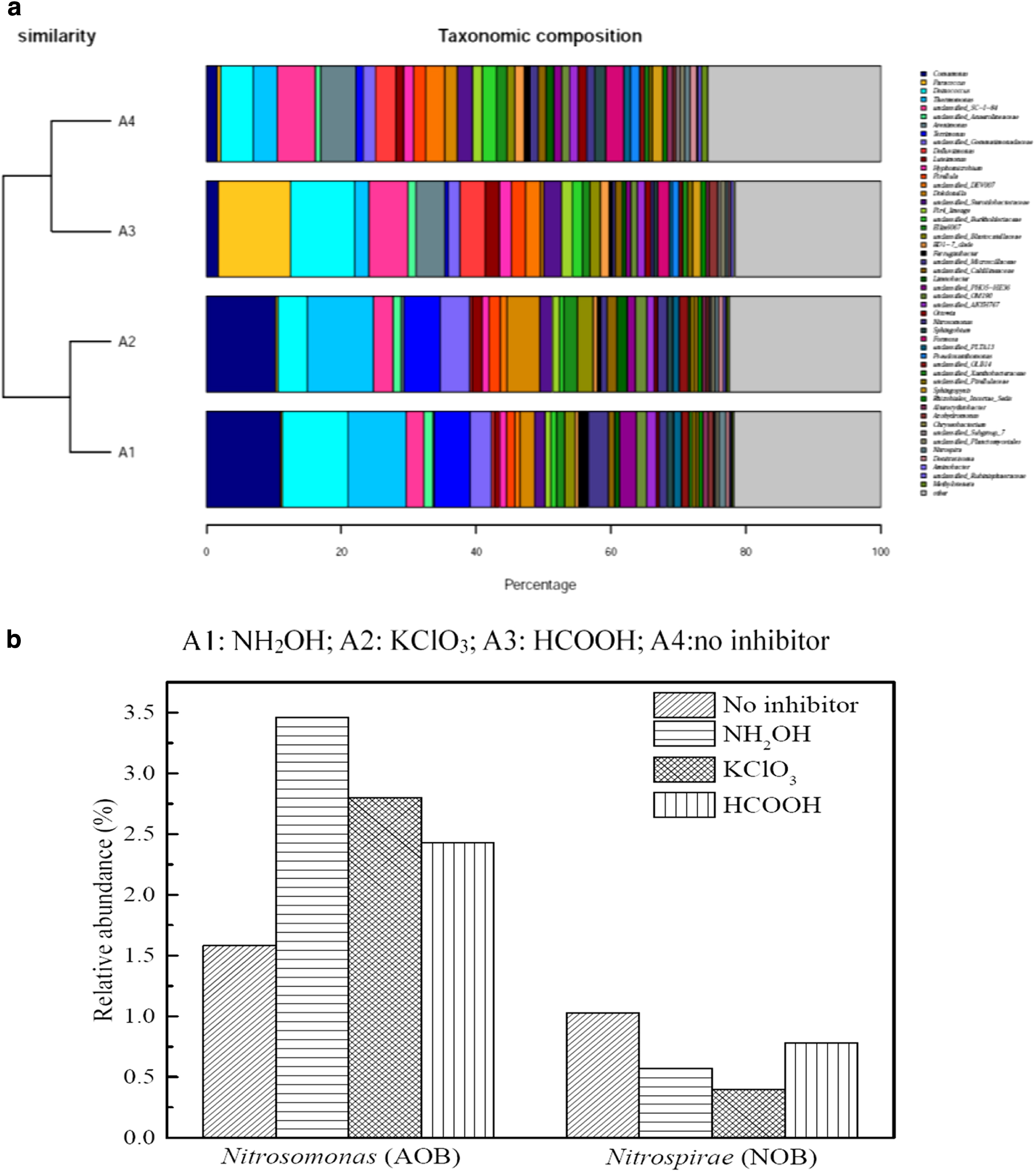
The composition of
Conclusion
This study investigated the feasibility of inhibitor addition on the start-up and real-time aeration control strategy on the maintenance of partial nitrification. First, the performances of different dosage inhibitors on the activity of AOB and NOB were compared. The results showed that NOB was more sensitive to NH2OH and KClO3 than HCOOH. Second, based on the results of long term experiments, the addition of each inhibitor was a favor for the rapid start-up and recovery of partial nitrification. NH2OH performed better than the other two inhibitors in a similar range. When using real-time aeration control strategy after ceasing the inhibitor addition, a stable level with a NAR of 80% was still achieved. The results of high-throughput sequencing revealed that the inhibitor selectively suppressed the transcription and growth of NOB. In addition, different inhibitors caused a difference in bacterial community structure. The method herein may have important scientific and practical significance for the practical application in partial nitrification.
Footnotes
Author Disclosure Statement
No potential conflict of interest was reported by authors.
Funding Information
This study was financially supported by the National Natural Science Foundation of China (Grant No. 51408295), Key Research and Development Project of Shandong Province (Grant Nos. 2017GSF217013 and 2018GSF117007), and Natural Science Foundation of Shandong Province (Grant No. ZR2019BEE047).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.