
Abstract
Biochar has been widely used for soil remediation and carbon sequestration. However, the effects of biochar and earthworms on the mineralization of soil organic matter (SOM) remain controversial. In this study, SOM mineralization in top and deep soils with or without earthworm activity and 0.5% (w/w) maize straw biochar was investigated during a 43-day incubation experiment. The results showed that the organic carbon content was reduced after incubation, particularly during earthworm activity. The addition of biochar retained easily degradable fractions (such as tryptophan and tyrosine), whereas earthworms promoted the degradation of these fractions. Earthworms significantly increased the soil respiration rate and soil carbon emissions. The addition of biochar stimulated SOM mineralization under the effect of earthworms in the top soil within 7 days; however, biochar inhibited carbon emissions under earthworm activity for a longer time. Compared with deep soil, earthworms had a stronger respiration effect on top soil, which was mainly attributed to the more easily degradable SOM in the top soil. Overall, the positive effects of biochar on carbon sequestration were more pronounced under earthworm activity. This study emphasizes that soil mineralization was affected by biochar and soil macrofauna, particularly in areas with intense human activity.
Introduction
Mineralization is the process of decomposition of soil organic matter (SOM) into inorganic compounds and carbon dioxide (CO2) under the action of soil microorganisms (Shahbaz et al., 2017) and is a key link between soil carbon stabilization and carbon loss processes. When the ground surface is covered by exogenous organic matter (OM), such as litter, OM mineralization is enhanced because the original microorganisms boost their metabolism (Yu et al., 2020). The main mechanisms by which exogenous OM promotes or inhibits OM mineralization include the promotion of synergism between microorganisms and their secreted extracellular enzymes (Geisseler et al., 2011; Li et al., 2017), promotion of the use of limited resources by microorganisms (Paterson and Sim, 2013; Wang et al., 2015), and selective use of decomposable substrates by microorganisms (Miao et al., 2017). Thus, SOM mineralization is closely related to the quality of exogenous OM and microbial activity.
Biochar is prepared from solid biomass via pyrolysis under anoxic or low-oxygen conditions (Wang et al., 2017). Most studies have suggested that the addition of biochar reduces carbon emissions, increases SOM, and improves soil quality effectively (Ameloot et al., 2014; Lu et al., 2014). However, some studies have found that biochar can increase carbon emissions under certain circumstances. For example, the solubility of soluble OM, microbial activity, and SOM mineralization can be enhanced by biochar (Luo et al., 2011; Zimmerman et al., 2011). Moreover, biochar promotes the mineralization of carbon, which is attributed to the fast utilization of a small labile component of biochar (Chen et al., 2023).
Recently, a study reported that the addition of biochar significantly improved the accumulation of inorganic carbon in calcareous subsoil (Wang et al., 2023), which is important for understanding carbon sequestration in calcareous soils. Therefore, the effect of biochar on carbon mineralization/sequestration is related to unstable OM content, soil type, and natural SOM state (Purakayastha et al., 2016).
In addition, bioturbation affected SOM mineralization. Earthworms are common soil animals that play an important role in soil ecology because of their impact on soil physical and chemical properties, soil structure, and the global carbon cycle through feeding, digestion, excretion, and burrowing (Sharma et al., 2017). Moreover, earthworms disrupt soil aggregates and expose OM to mineralization through digging, burrowing, and feeding activities in the soil (Mbau et al., 2015). However, recent studies have found that earthworms improve SOM stabilization. For example, earthworm mucus contributes to the formation of organomineral associations and promotes the formation of macro- and microaggregates (Guhra et al., 2020; Le Mer et al., 2020).
In addition, Zhang et al. (2013) reported that earthworms facilitate carbon sequestration through the unequal amplification of carbon stabilization compared to mineralization. The presence of earthworm casts improves soil aggregation and aggregate-associated carbon stability in soil (Li et al., 2021). Therefore, the effects of earthworms on SOM mineralization remain unknown.
Earthworm abundance increases with intensified organic fertilization or conservation tillage, which amplifies their importance to SOM turnover in agroecosystems (Angst et al., 2019; Lubbers et al., 2013). Biochar has been widely used in soil remediation because of its desirable properties, such as its carbon-rich and abundant porous structure. However, the combined effects of earthworms and biochar on SOM mineralization, particularly at different soil depths, are poorly understood. Therefore, this study focused on exploring the effect of exogenous biochar on SOM mineralization with the interference of earthworms to clarify the following problems: (1) the effect of biochar on SOM mineralization in surface and deep soils; (2) the effect of earthworms on SOM mineralization in surface and deep soils; and (3) the effect of biochar on SOM mineralization with the interference of earthworms.
Materials and Methods
Sample collection and pretreatment
The corn straw was collected, air-dried, ground, and pyrolyzed at 550°C for 4 h. After pyrolysis, the corn straw-derived biochar was ground and sieved through a 100-mesh. The soil samples were collected in Laoyuhe Park next to Kunming University of Science and Technology in Chenggong District, Kunming City, Yunnan Province (N: 24°51′38.53″, E: 102°52′1.99″), trees were planted on the land (Pinus yunnanensis), the ground surface was covered with a large amount of dead leaves, and the trees had a well-developed root system. A plot of land was randomly selected with a 2 × 2 × 1 m3 excavated soil pit and sampling depths of 10–20 cm and 60–80 cm from the surface, which were marked as top soil (B) and deep soil (S), respectively.
Top and deep soils were selected to explore the effects of biochar and earthworms on soil mineralization under different SOM contents. Plant litter was manually removed from the soil, and the soil samples were stored in sterile sealed bags in the laboratory at −40°C in a freezer. A portion of the soil sample was air-dried and passed through a 100-mesh sieve to measure its physicochemical properties. The earthworm used in the experiment was Eisenia fetida, which were starved on a moist filter paper for 24 h to void their guts.
Soil culture experiment
Approximately 150 g of dry soil and 0.75 g (0.5% of total weight) of biochar were added to each group of flasks and incubated at 25°C in the dark, with the soil moisture maintained at ∼30%. Eight culture groups were established with three replicates (N = 3): top soil (B), top soil with biochar (B-BC), top soil with earthworms (B-E), top soil with biochar and earthworms (B-E-BC), deep soil (S), deep soil with biochar (S-BC), deep soil with earthworms (S-E), and deep soil with biochar and earthworms (S-E-BC). After the end of experiment, the earthworms were released into the field.
Detection of CO2 release
The CO2 concentration was measured once weekly, and the detailed operation was as follows: First, the air in the jars was replaced by CO2-free air using an inflatable pump, and then the jars were closed with rubber stoppers for 12 h. Then, 1 mL of collected CO2 was detected using a Gas Chromatography-Flame Ionization Detector with the following parameters: CH4 (10.10%); CO (10.00%); CO2 (9.93%); air pressure (101 kPa); and temperature (273 K + 20°C).
Three-dimensional fluorescence measurement
The measured total organic carbon (TOC) concentration was diluted to below 10 mg/L and measured using a PerkinElmer LS55 instrument with an excitation wavelength range of 200–550 nm and excitation sampling interval of 10 nm, emission wavelength (Em) range of 200–550 nm and emission sampling interval of 10 nm. The excitation and emission slit widths were 200 nm, the scanning speed was 8,000 nm/min, the number of measurements was 70; and the three-dimensional fluorescence was measured at 680 V.
According to a previous method (Gao et al., 2016; Li et al., 2019), the excitation-emission matrix was divided into five regions (I, II, III, IV, and V) according to the distribution of different components of soluble OM in the excitation/emission (Ex/Em) wavelength region of the fluorescence spectrum. Region I (220–250 nm/280–330 nm) was a tryptophan-like protein, region III (220–250 nm/380–480 nm) was a fulvic acid-like protein, region IV (250–360 nm/280–380 nm) was a soluble microbial metabolite, and region V (250–420 nm/380–520 nm) was a humic acid-like protein. An emission wavelength of 520 nm was selected, and an excitation wavelength of 300–500 nm was scanned to obtain the excitation spectrum, and the fluorescence intensity ratios at 436 and 383 nm (I436/383) were calculated (Provenzano et al., 2004).
Sample characterization experiments
The prepared biochar materials and cultured samples were characterized for their physicochemical properties, including pH determination (pHS-3; Leica Instruments, Wetzlar, Germany), TOC analysis (Vario TOC Cube; Elementar, Langenselbold, Germany), and elemental analysis (Vario Max; Elementar) to determine their pH, dissolved organic carbon (DOC), and elemental content, respectively.
Data analysis
Statistical analyses were performed using SPSS 16.0 software package for Windows (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance with a 5% significance level was used to detect significant differences in soil CO2 emission rates and cumulative CO2 emissions. The pH, elemental analysis, DOC content, emission rate, and cumulative emissions of soil CO2 were averaged.
The CO2 concentration represented by the gas chromatography peak was obtained from the CO2 marker, and the CO2-O release rate was calculated as follows:
where c1 and c2 are the concentration of CO2 at time 0 and the concentration of CO2 in the incubator at 12 h, respectively. Mc is the molecular mass of elemental carbon (12 g/mol), V is the volume of the space in the upper part of the bottle (m3), m is the dry weight of the soil (kg), MVcorr is the molar volume of the gas after correction for temperature (0.0242 m3/mol), and t is time (h).
The relationships between the cumulative soil respiration C(d), g CO2/g soil·days, and soil respiration rate C(S), and g CO2/g soil·days were as follows:
where C(l) is the accumulation of the previous soil respiration and the first C(l) value is 0.
Results and Discussion
Effect of earthworms and biochar on elemental composition of soil
The elemental compositions of the original and cultivated soils are listed in Table 1. Before incubation, the bulk organic carbon (OC) in the top soil was slightly higher than that in the deep soil. After incubation, except with the addition of biochar, the bulk OC content decreased to a certain extent, particularly in the presence of earthworms. The nitrogen (N) did not decrease or increase, which may be related to the secretion of nitrogenous compounds by the microorganisms during the incubation process. Therefore, the OC/N ratio decreased significantly after incubation. The OC/N ratio in the presence of biochar was relatively high, which was mainly attributed to the input of biochar-derived carbon.
Changes in Soil Elemental Composition Before and After the Incubation
B, topsoil; B-BC, top soil with biochar; B-E, top soil added with earthworms; B-E-BC, top soil added with biochar and earthworms; N, nitrogen; OC, organic carbon; S, deep soil; S-BC, deep soil with biochar; S-E, deep soil added with earthworms; S-E-BC, deep soil with biochar and earthworms; YB, original top soil; YS, original deep soil.
The soil pH ranged from 6.27 to 7.12 (Table 2), indicating a neutral bias. The addition of biochar increased soil pH, whereas the presence of earthworms slightly decreased it. The pH of the biochar was ∼10.1, therefore, the pH of the soil increased to a certain extent after the addition of biochar. Earthworms secrete organic acids, leading to a decrease in soil pH (Valdez-Perez et al., 2011). In addition, the DOC content in the soil decreased after incubation, and the trend was B/S-BC > B/S > B/S-E-BC > B/S-E (Table 2). The most significant decrease was observed in the presence of earthworms alone, which may be because earthworms accelerate the decomposition of DOC through the indirect or direct stimulation of microbial activity (Ahmed and Al-Mutairi, 2022; Fujimaki et al., 2017).
Changes in Soil Dissolved Organic Carbon and pH (Mean ± Standard Error) Before and After Incubation
DOC, dissolved organic carbon.
However, the degree of decrease in the DOC content with the addition of biochar was the lowest. On one hand, this phenomenon may be due to the leaching of DOC from the biochar itself; on the other hand, it is possible that the adsorption of DOC by biochar inhibited the degradation of DOC. Considering that the biochar was repeatedly washed before addition and added in smaller amounts, DOC released from the biochar may contribute less to its DOC within a short incubation time (7 days). Therefore, it is more likely that the small reduction in DOC content with biochar addition was due to its adsorption by the biochar, which inhibited its degradation and thus retained a higher DOC content. This was confirmed by the release of CO2.
Compared with the top soil, the DOC content in deep soil, generally with higher mineral fractions and relatively lower microbial activity, was reduced to a lesser extent after incubation (Rukshana et al., 2013; Zhao et al., 2018). The composition of DOC may further influence this phenomenon, which can be elucidated from the three-dimensional fluorescence map of DOC. This may lead to a relatively slow decomposition of DOC in the deep soil, which is further evidenced by the release of CO2.
Effect of earthworms and biochar on dissolved organic fraction of soil
To further investigate the differences in DOC composition, we analyzed its composition with three-dimensional fluorescence spectroscopy (Fig. 1). Before incubation, the DOC in both the top and deep soil layers was dominated by tyrosine and tryptophan. The response degrees of tyrosine and tryptophan were higher in the top soil. In general, tyrosine and tryptophan decompose easily (Tremblay et al., 2007). Thus, the top soil had more easily decomposed substances than the deep soil. In contrast, the DOC in the deep soil showed a response at 420 nm, which belonged to the fulvic acid region. It was more difficult to degrade because of its large molecular weight compared to tyrosine and tryptophan, further indicating that the composition of DOC in the deep soil was more difficult to degrade than that in the top soil.
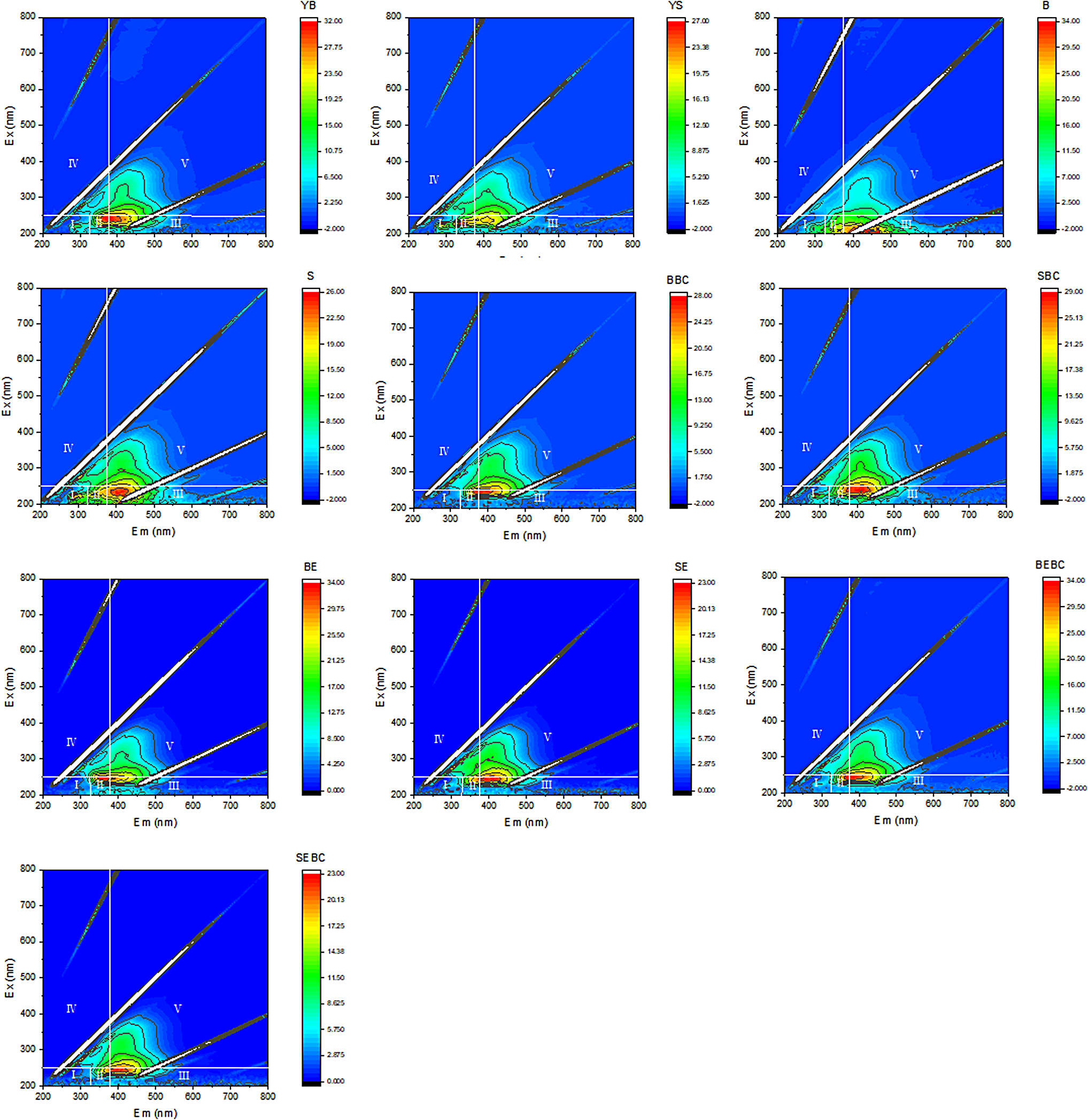
Three-dimensional fluorescence of different soils before and after incubation. B, top soil; B-BC, top soil with biochar; B-E, top soil with earthworms; B-E-BC, top soil with biochar and earthworms; S, deep soil; S-BC, deep soil with biochar; S-E, deep soil with earthworms; S-E-BC, deep soil with biochar and earthworms; YB, original top soil; YS, original deep soil.
After incubation, the DOC compositions of the top and deep soil layers were dominated by fulvic acid, which indicated that tyrosine and tryptophan were depleted during incubation. Notably, the response intensity of the fulvic acid region of the top soil DOC was higher than that of the deep soil (approximately three times higher than that of the deep soil), indicating that more fulvic acid fractions were present in the top soil after incubation. This may be because the SOM in the top soil was more easily decomposed into soluble fulvic acid by microorganisms.
Furthermore, the response to fulvic acid was more pronounced in the presence of earthworms, and the fulvic acid fraction of DOC in the top soil was approximately five times greater than that in deep soil. This further demonstrates that earthworms promote the decomposition of SOM and alter the composition of DOC in the soil leachate. In contrast, after biochar addition, the responses of tryptophan, tyrosine, and fulvic acid remained strong in the soil. Previous results showed that the DOC content decreased less after biochar addition. This result further indicates that the addition of biochar inhibited the decomposition of easily degradable fractions.
Effect of earthworms and biochar on mineralization of SOM
CO2 release rate
According to the changes in the soil respiration rate, the respiration rate of soil CO2 decreased with incubation time, which was mainly due to the decrease in microbial activity resulting from a reduction in food sources. There was a decrease in the soil respiration rate after the addition of biochar, which was consistent with the earlier DOC and fluorometric measurements. In addition, the respiration rate in the top soil was slightly higher than that in the deep soil after biochar addition (Fig. 2a, b). When the OC/N was >20, the soil microorganisms had more pronounced nitrogen fixation to maintain an OC/N ratio suitable for their survival, and the decomposition of OM by microorganisms may become lower (Sigua et al., 2014). However, the deep soil clearly contained a lower content of readily decomposable SOM.

Histogram of soil respiration rate and cumulative soil carbon emissions in the experimental group of biochar addition culture in the presence of earthworms.
The CO2 respiration rate was markedly higher in the presence of earthworms than in the earthworm-free treatment group. At the beginning of the incubation period, the CO2 respiration rate reached 100 g/(g·day). Earthworms have a respiratory effect on CO2 emissions (Binet et al., 1998). Moreover, the addition of earthworms may increase the activity of microorganisms, resulting in the mineralization of most SOM in the soil (Hoang et al., 2017). Considering the significant decrease in easily degradable DOC after the addition of earthworms, as previously reported, earthworms accelerated the mineralization of SOM, particularly for easily degradable DOC. In addition, the respiration rate of the top soil was higher than that of the deep soil. This result was attributed to the higher SOM content of the top soil and a higher number of microbial populations (Zhao et al., 2018).
Meanwhile, the SOM in the deep soil was more stable and difficult to utilize. The addition of biochar inhibited the release rate of CO2 in the earthworm group, which was partly explained by the protection of SOM by the biochar. Moreover, earthworm activity in the experimental group where earthworms were cocultured with biochar was reduced relative to that in the experimental group where only earthworms were added. Thus, a decrease in earthworm activity may be another important reason for this inhibition.
Cumulative release of CO2
The cumulative release of CO2 after earthworm treatment was significantly higher than that after the earthworm-free treatment, which was consistent with the release rate of CO2. In the earthworm-free treatment, the addition of biochar did not change the cumulative release of soil CO2. However, under the earthworm treatment, the addition of biochar resulted in a lower cumulative release of soil CO2 than earthworm activity alone, which was more pronounced with incubation time. Most studies have suggested that biochar has insignificant effects on soil carbon sequestration in the short term and may even have positive priming effects (Major et al., 2010).
However, our study found that biochar inhibited the short-term release of CO2 in the presence of earthworms, particularly in deep soils with low SOM content. This may be because earthworms promote the interaction between biochar and SOM, thus avoiding the further decomposition of SOM by adsorption and even formation of agglomerates, which complied with the composition of three-dimensional fluorescent DOC. In addition, biochar may also be toxic to earthworms (Sohi et al., 2010), and it was found that earthworms had a certain tendency to avoid biochar at the beginning, and the earthworm activity was weaker in the earthworms with biochar added, which may also contribute to reducing cumulative soil respiration.
Conclusion
This study investigated the effects of biochar and earthworms on mineralization and SOM components in top and deep soil during a 43-day incubation experiment. After incubation, the bulk OC and DOC contents decreased. The addition of earthworms significantly enhanced CO2 emissions; however, biochar inhibited the emissions of CO2 compared to the control (B/S groups). A similar result was observed for the change in DOC content. Notably, the soil respiration rate in the earthworm culture group decreased with time after the addition of biochar, and there was a partial tryptophan and tyrosine response at the end of the experiment. This indicates that biochar reduced the mineralization caused by earthworms, protecting the easily decomposable SOM in the soil.
In addition, the mineralization response of the top soil was more evident than that of the deep soil, which was mainly due to the higher content of easily decomposed SOM in the top soil. This study demonstrates the important role of biochar in soil carbon sequestration, particularly in the presence of earthworms in short-term cultures.
Footnotes
Acknowledgments
We express thanks to Xiaobo Yang for her help in writing this article; and Xiaoyu Gui for her support in the experimental manipulation.
Authors' Contributions
F.L.: investigation, data curation, methodology, and writing—review and editing. J.Z.: investigation, data curation, and writing—review and editing. X.G.: methodology and writing—original draft. S.F.: modification. L.W.: modification. H.P.: modification and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by Yunnan Major Scientific and Technological Projects (202202AG050019), the National Natural Science Foundation of China (42267028, 41907300), the Basic Research Program of Yunnan Province (202001AU070088).