
Abstract
As source waters for many drinking water treatment plants are increasingly subject to intermittent blooms of cyanobacteria, utilities must adapt treatment methods to remove associated cyanotoxins. There is concern that treatment methods applied to remove dissolved extracellular toxins may damage whole cells, leading to the release of intracellular toxins and other secondary metabolites. The objective of this study was to determine whether sodium hypochlorite and potassium permanganate preoxidation treatments cause increased susceptibility of cyanobacterial cells to mechanical shear using a method that can be easily adapted by utilities. Bench-scale simulations examined Microcystis sp., Planktothrix sp., and Dolichospermum sp., which are three types of microcystin-producing cyanobacteria that are prominent in many bloom-impacted waters. We found that chlorine led to increased susceptibility to shear in Dolichospermum and permanganate led to increased susceptibility to shear in Planktothrix, each for shear rates of 750 s−1 (a typical rate for in-line mixers used in drinking water treatment). We found that shear does not cause immediate critical damage to cells, regardless of preoxidation conditions. However, shear appears to cause a stress response in cyanobacteria, indicating the possibility of delayed toxin release. The results of this study will help water treatment utilities to manage stressors within the treatment process that could lead to release of cellular compounds during cyanobacterial blooms. Additionally, utilities can adapt the applied method of intracellular phycocyanin analysis to independently evaluate real-time cell response to their treatment processes.
Introduction
Many drinking water utilities must respond to harmful algal blooms of cyanobacteria in their source water. The situation often requires removal of cyanotoxins and taste and odor compounds before distributing water to customers. Oxidation, adsorption, and physical removal by coagulation/flocculation are generally strong barriers against harmful algal bloom impacts on finished drinking water. For cyanotoxin removal, the use of each barrier depends on the form of the toxin. Source waters can contain both extracellular toxins that are dissolved in the water and intracellular toxins that are contained within living cells. Oxidation and adsorption are employed for extracellular toxin removal, whereas coagulation/flocculation is used for intracellular toxin removal through whole-cell removal (Newcombe et al., 2010). There is concern that treatment processes may damage intact cells, thereby leading to release of additional dissolved cyanotoxins (Wert et al., 2019). It is easier to remove toxins in the intracellular form rather than extracellular, so utilities should choose treatment strategies that minimize damage to cells.
Oxidation has received the most attention as a treatment that can cause damage to cyanobacteria. Most concern is specific to the use of preoxidants because they are commonly employed before cyanobacteria are removed from water. It has been shown that many oxidants, such as chlorine, permanganate, and ozone, pose risk to cyanobacteria (Fan et al., 2013; Ma et al., 2012; Wert et al., 2013). Studies have reported that chlorine causes lysis to a majority (more than 50%) of a cyanobacteria population at 1 mg-hypochlorite (OCl−)/L, even with short contact times of 30 min (CT = 30 mg-OCl−-min/L) (Ma et al., 2012; Qi et al., 2016; Wert et al., 2013). Reports for permanganate oxidation indicate that 3 mg-permanganate (MnO4−)/L causes lysis to most cyanobacteria species, particularly when contact times above 2 h are used (CT = 360 mg-MnO4−-min/L) (Ding et al., 2010; Greenstein et al., 2020; Li et al., 2021; Li et al., 2014; Ou et al., 2012). Other oxidation processes, such as the use of ozone or advanced oxidation processes, may cause damage to cyanobacteria but they are less common treatments (Thompson and Drago, 2015).
Response to chemical oxidation varies based on cyanobacteria type, which many studies attribute to morphological characteristics. Physical characteristics include cell membrane thickness, presence of a mucilage coating, and the tendency to form a colony or filament and the size of that formation (Clemente et al., 2020; Pestana et al., 2019; Wert et al., 2013). Chemical characteristics, such as the composition of the cell membrane, are also suspected to play a role in oxidant susceptibility (Oliveira et al., 2021). Thus, there is much evidence that oxidants damage cyanobacteria, but the extent of damage varies by oxidant type and cyanobacteria species.
The complicated nature of oxidative cell damage poses a challenge to providing concrete guidance to utilities for avoiding toxin release during treatment. Generally, guidance suggests stopping preoxidant use when intact cells are present (Newcombe et al., 2010) or ensuring that cells are removed before oxidative treatments are applied (Wert et al., 2019). However, preoxidation is often used for other treatment goals (Cherry, 1962), so suspending its use when cyanobacteria are present is not always practical. A third option for managing toxin release is to apply sufficient oxidant to completely lyse cells and destroy resulting toxins (Wert et al., 2019). The risk of downstream toxin concentrations remains if complete lysis and destruction of toxins is not achieved. It can be challenging to successfully apply this strategy because utilities must consider characteristics of the dominant cyanobacteria and the type of cyanotoxin when determining oxidant type and dosage (Wert et al., 2019). If cyanobacteria lysis is not intended during preoxidation, guidance recommends using permanganate rather than chlorine because permanganate causes less damage at low dosages.
It has recently been recommended to limit permanganate dosing to 1 mg-MnO4−/L or less (GLUMRB, 2022). Newer guidance is more specific than that which had existed previously, but it fails to consider how effects of other treatment processes may play a role in cyanobacteria damage. Guidance documents express concern over cell damage and the immediate release of toxins but do not address how treatment sequences may progressively weaken cells, thereby increasing potential for delayed toxin release.
Cyanobacteria exposure to low dosages of oxidants is more concerning when the cells are subjected to downstream stressors. Cells may become more vulnerable to damage by mechanical abrasion, such as through pumping, mixing, or filter processes, after they have been weakened by preceding oxidant exposure. Studies have reported varying levels of shear damage to different species of cyanobacteria, likely because of varying morphological characteristics (Chow et al., 1999; Clemente et al., 2020; Oliveira et al., 2021; Sun et al., 2012). Cyanobacteria morphologies that affect susceptibility to shear include cell shape and cell wall constituents, such as polysaccharide content (Clemente et al., 2020; Oliveira et al., 2021).
Shear damage also depends on the fluid shear rate or velocity gradient (G). For example, damage to Microcystis aeruginosa cells has been reported at high shear rates (G = 1,000 s−1) (Oliveira et al., 2021), but the same species was not damaged in fluids moving at low rotational speeds (Chow et al., 1999; Sun et al., 2012). Typical drinking water operations use velocity gradients between 750 and 1,000 s−1 (Davis and Cornwell, 2013), presenting shear conditions that could be damaging to cyanobacteria.
Furthermore, conclusions regarding cell susceptibility to shear have been drawn from studies which did not consider effects of upstream chemical oxidation. Pestana et al. (2019) assessed how preoxidation affects cyanobacteria susceptibility to shear conditions in the coagulation process and found that Planktothrix cells were the only species of several studied that became more vulnerable to shear following oxidant exposure. Their study included the use of coagulant chemicals, leading the authors to hypothesize that cells may have been protected from oxidative damage by flocs formed during the coagulation step (Pestana et al., 2019). The inclusion of coagulants and the formation of flocs make it difficult to draw conclusions about how preoxidation directly affects the response of cyanobacteria to shear conditions. The question remains to understand how cyanobacterial cells respond to successive chemical and mechanical treatments as drinking water utilities regularly employ preoxidants as part of their process operations.
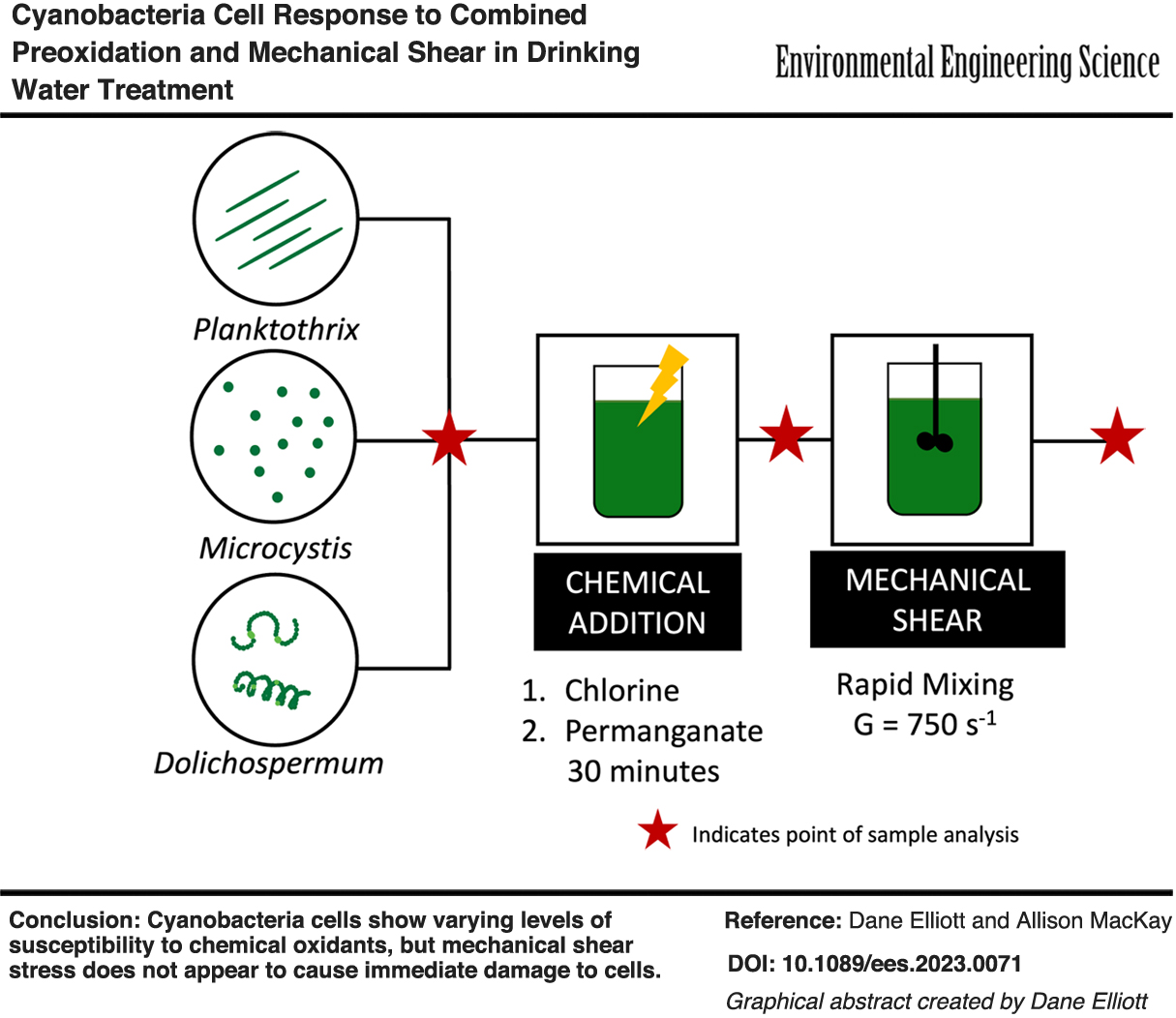
The objective of this study was to determine whether chemical preoxidation increases cyanobacterial cell susceptibility to mechanical shear. We studied Microcystis sp., Planktothrix sp., and Dolichospermum sp. because they are each toxin-producing cyanobacteria that are prominent in coastal and inland lakes in Ohio. The study used measures of intercellular phycocyanin, a photosynthetic pigment produced by cyanobacteria, to approximate the number of whole cells that remain following oxidation and shear treatments. We developed the intracellular phycocyanin method considering that it could be applied as a quantitative tool for approximating cell viability in a modestly equipped laboratory setting. We note that we did not measure cyanotoxins directly in this study because our accessible cell cultures were nontoxin-producing strains. Nonetheless, results of this study will assist utilities to manage water treatment operations with the goal of minimizing damage to whole cells so as to avoid release of intracellular toxins and other unwanted metabolites, such as taste and odor compounds.
Methods
Cyanobacteria cultures
Three types of cyanobacteria were selected for this study based on their presence both in Ohio and globally and because of their ability to produce toxins in the environment. These three genera also represent different cell morphologies: coccoidal colonial organisms (Microcystis), cylindrical filamentous organisms (Planktothrix), and coccoidal filamentous organisms (Dolichospermum).
A nontoxin-producing strain of Microcystis was obtained from Nicholas Dugan (U.S. Environmental Protection Agency, Water Infrastructure Division). Microcystis cells were maintained in BG-11 medium. Raw water samples containing Planktothrix cells were collected from Grand Lake St. Mary's in Celina, Ohio. Planktothrix cells were maintained in modified Bristol Solution containing sediment from the reservoir where they were obtained. Both species were maintained under a photoperiod of 12-h light/12-h dark with irradiance of 5 ± 1 μmol photons/m2·s (measured through LI-COR LI-190R Quantum Sensor) and under constant temperature (25°C ± 2°C) conditions. Dolichospermum cells were not cultured in the laboratory due to challenges associated with obtaining and maintaining pure cultures. Instead, Dolichospermum samples were collected from Buckeye Lake, Ohio during a bloom and experiments were performed within 24 h of collection.
Cell density
Optical density at 680 nm (OD680) was used as a proxy to express cell density because this measure is readily adoptable by utilities for monitoring cell density, compared with other cell counting techniques. We validated OD680 measurements by checking correlations between direct cell counts for Microcystis (Supplementary Fig. S1 in the Supplementary Data) and between biovolume for Planktothrix (Supplementary Fig. S2 in the Supplementary Data) so our results could be compared with other studies. Dolichospermum was only measured through OD680 because we did not have an established counting method for this cyanobacterium type at the time of collection.
Oxidation experiments
Batch oxidation experiments were conducted individually for each of the three cyanobacteria species to assess the impacts of chlorine or permanganate addition on cell integrity. For Planktothrix cells, an OD680 of 0.2 ± 0.05 Abs was used, which corresponded to a biovolume of 17 ± 4.6 mm3/L. Planktothrix samples naturally maintained themselves at this density and thus, they were not diluted before experimentation. Microcystis culture samples were diluted in 0.01 M phosphate-buffered saline (PBS; 1 × PBS) to an OD680 of 0.2 ± 0.05 Abs, which is ∼2.7 ± 1.25 × 105 cells/mL, to provide a comparable density to Planktothrix. Dolichospermum cells were collected at an optical density of 0.1 ± 0.01 Abs and used unaltered in experiments.
Water quality parameters were collected before each experiment to examine whether they may cause variations in the oxidant activity between experiments (e.g., loss of oxidant through competing reactions with organic matter). Solution pH was measured using a benchtop pH meter (Thermo Scientific Orion 2-Star Benchtop pH Meter), which was calibrated weekly. A portion of each sample was filtered (Millex-LCR Syringe Filter, 0.45 μm hydrophilic PTFE), and the absorbances at 254 and 365 nm were measured for E2/E3 (Sharpless et al., 2014) and SUVA-254 calculations. Dissolved organic carbon (DOC) was measured on filtered samples using a Shimadzu TOC-V CSN total organic carbon analyzer according to the 5310B combustion-infrared method (APHA et al., 2017).
Sodium hypochlorite (11–15% available chlorine, No. 7681-52-9; Alfa Aesar) and potassium permanganate (99%+ ACS reagent grade; Acros Organics) were used for oxidation experiments. Stock solutions for chlorine (1.0 M free chlorine) and permanganate (0.1 M permanganate) were kept in the dark at 4°C. The concentration of free chlorine was verified before each use by measuring the OCl− concentration through absorbance at 292 nm (ɛ = 362 M−1/cm) (Chen, 1967). The permanganate (MnO4−) concentration was verified by measuring the absorbances at 526 and 546 nm before each use (ɛ = 2,400 and 2,380 M−1/cm, respectively) (Den Boef et al., 1958). Working solutions of each chemical were prepared daily in ultrapure water (18.2 MW·cm, Millipore Milli-Q deionized water system).
For each batch experiment, stock oxidant solutions were added to prepared cell solutions to achieve concentrations from 0 (negative control) to 2 mg-OCl−/L or from 0 (negative control) to 20 mg-MnO4−/L in separate muffled glass vials. Triplicate samples were prepared at each oxidant concentration. The vials were sealed and maintained in darkness to limit photodegradation of the oxidant. Evaporative losses were considered negligible because of the pH and short reaction times compared with those reported of chlorine evaporation (Len et al., 2002). The vials were placed on an orbital platform shaker and gently rotated (<100 rpm) throughout the oxidation experiment. In initial experiments, each sample was quenched after 30 min of reaction time to a concentration of 80 mg/L sodium thiosulfate (No. S445-500; Fisher). In secondary experiments evaluating extended contact times, aliquots were obtained at 0.5, 1, 2, and 4 h and then quenched. The remaining cell suspension was then used in shear experiments, also performed in triplicate.
Shear experiments
The effect of shear on cell integrity was evaluated using a small-volume (45 mL) baffled glass vessel and impeller designed for these experiments to simulate rapid-mix tank conditions. Cell samples not exposed to chemical oxidants were prepared at the same optical density as noted above for oxidation experiments and added to the tank. Each sample was subjected to a velocity gradient (G) of 750 s−1 for 2 min. We note that, while 750 s−1 is a typical G value for rapid mixing (Davis and Cornwell, 2013), the residence time of 2 min included in this study is longer than typical for in-line mixing. This procedure was repeated for solution aliquots obtained at the end of each oxidation experiment to assess effects of prior oxidant concentration exposure on cell integrity. After shear exposure, the cell integrity of each sample was then analyzed within 60 min through intracellular phycocyanin analysis.
Intracellular phycocyanin extraction
Measurements of intracellular phycocyanin were used to quantify viable cells remaining in each sample before and after experimental manipulation. Cells for each of the three cyanobacteria genera were processed using the same method for intracellular phycocyanin extraction. Each extraction was performed in triplicate. Cell suspensions were centrifuged at 3,150 g for 15 min. The supernatant was gently removed and discarded so that phycocyanin could be removed from the remaining intact cells. The cell pellet was resuspended in 1 mL 1 × PBS and frozen at −20°C for 24 h before thawing at 4°C. Each sample was refrozen at −80°C for 15 min and thawed two more times for a total of three freeze–thaw cycles without changing the solution phase.
Samples were placed in the dark at 4°C for an extraction period of 96 h to allow phycocyanin to be released from cell materials into the solution phase. Then, samples were centrifuged at 3,150 g for 5 min to separate cell debris from the supernatant. Extracted phycocyanin in the supernatant, referred to subsequently as “remaining phycocyanin,” was quantified using fluorescence spectroscopy (BioTek Synergy HTX Multimode Microplate Reader).
The use of “remaining” intracellular phycocyanin to assess impacts of chemical and/or mechanical stressors on cyanobacterial cells assumes that increased cell damage or death will result in less phycocyanin being recoverable from the cell pellet by our extraction methodology. Cells that are impacted by oxidants that permeate cells and degrade intracellular components before other signs of cell lysis appearing (Ding et al., 2010) are counted as damaged cells by our method if phycocyanin is oxidized in the experimental process.
We validated the representativeness of the intercellular phycocyanin approach for a subset of samples by comparing the results to those obtained concurrently from a live/dead cell staining method (see Supplementary Data for details). We note that our goal was to employ a robust cell viability approach that would be consistent across cyanobacterial cell morphologies and that could be deployed readily in a modestly equipped laboratory. Alternative approaches of cell observation of live/dead cell staining or flow cytometry require specialized training or equipment, which limits their use by utilities in a standard water quality laboratory, especially as bloom conditions can change frequently over seasons.
Data analysis
Fluorescence intensity (arbitrary units [a.u.]) of remaining phycocyanin was used to evaluate effects of chemical oxidation and mechanical shear on the cyanobacterial cells. Remaining phycocyanin (%) for each sample, i, was calculated using Equation (1):
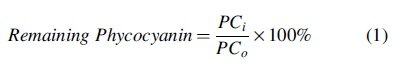
where PCi is the fluorescence intensity (a.u.) of the sample after oxidation and/or shear and PCo is the fluorescence intensity of the sample exposed to 0 mg/L oxidant (negative control). Each experimental sample included three replicates of phycocyanin extractions to quantify methodological error. Three replicates were conducted for each oxidant to quantify experimental variation. A one-way analysis of variance was used to analyze results of oxidation experiments and student's t-tests for paired samples were used to analyze shear experiments. A value of p < 0.05 was used to determine whether differences were significant.
Results and Discussion
Experimental variation
We evaluated both methodological and experimental error to determine the source of variation in reported experimental results of remaining phycocyanin after sample manipulation. The methodological error was 6% ± 6%, indicating strong reproducibility of the phycocyanin extraction method. Variation between experiments was 11% ± 10% across the three species, implying that factors between experiments have a greater contribution to reported error. The oxidation of Microcystis cells showed particularly high error (Fig. 1a) and is discussed in the following section. Variations between experiments could not be explained by correlations with water quality parameters (OD680, E2/E3, pH, DOC, and SUVA-254, data not shown) as could be the case if oxidant reactions with cells were limited by other side reactions, such as dissolved organic matter (Laszakovits and MacKay, 2019). We suspect that biological characteristics that were not directly tracked in this study, such as cell viability, may be the underlying source of variability between replicates.
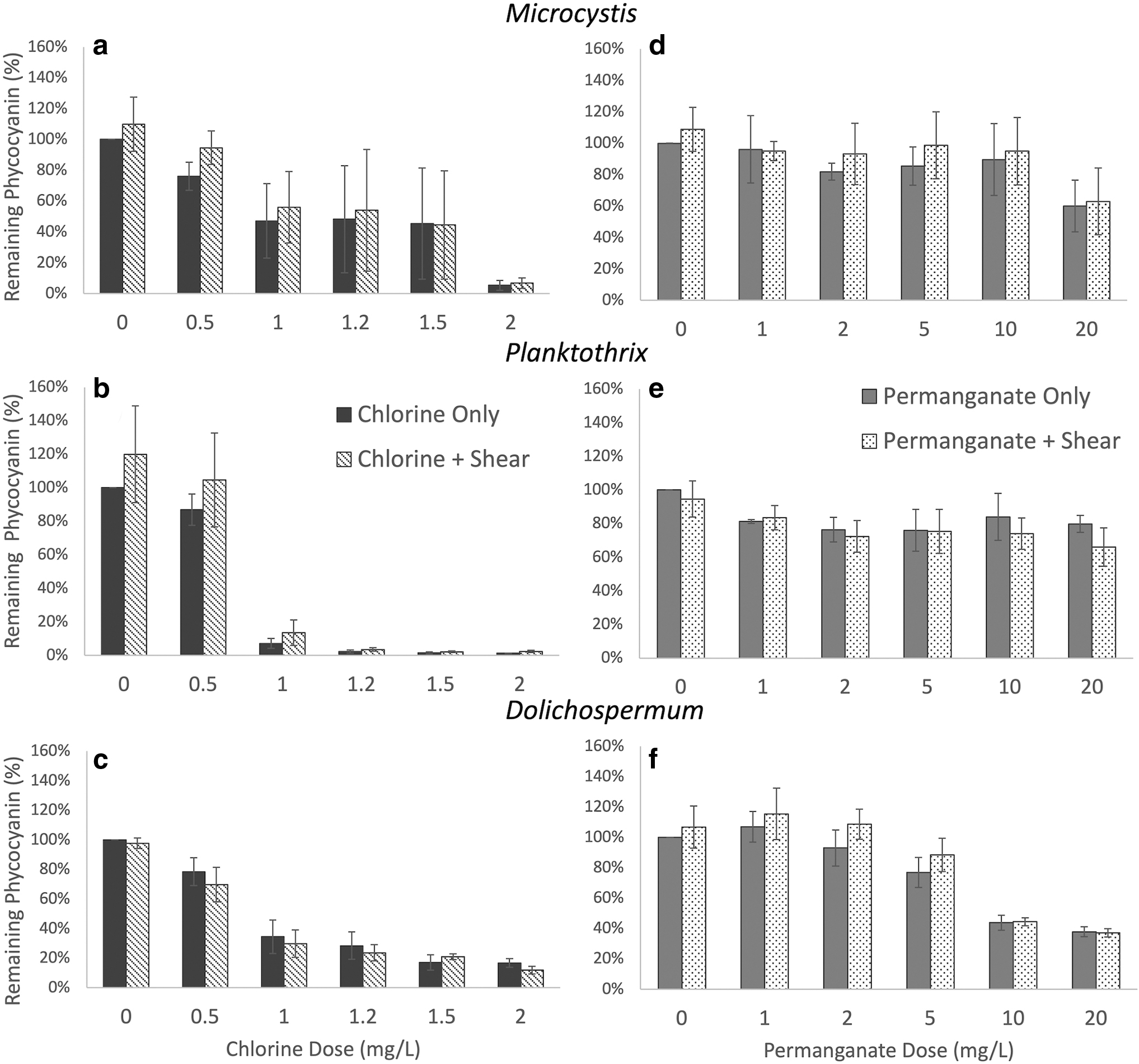
Chlorine and permanganate oxidation (30-min contact time) and shear results for Microcystis (top,
Oxidation damage to healthy cells
We first evaluated the effects of chemical oxidation independent of shear to confirm findings reported in literature and to serve as method of validation. We found that all three species of cyanobacterial cells sustained damage by chlorine with critical dosages, which we define as dosages causing damage to more than 50% of the cell population, ranging between 1 and 2 mg-OCl−/L during a 30-min contact time. Following exposure to chlorine, remaining phycocyanin decreased, which suggests that chlorine caused cell damage resulting in immediate phycocyanin release, leaving less remaining phycocyanin in the sample.
These findings are consistent with studies which report that chlorine works almost instantaneously and damages the majority (more than 50%) of a cyanobacterial population at concentrations around 1 mg-OCl−/L (Ma et al., 2012; Qi et al., 2016; Wert et al., 2013). The remaining intracellular phycocyanin levels decreased to 5% ± 3%, 1% ± 0.1%, and 17% ± 3% for Microcystis, Planktothrix, and Dolichospermum of initial concentrations, respectively, after exposure to a dosage of 2 mg-OCl−/L (Fig. 1). These results suggest that Planktothrix is the most susceptible to chlorine damage, followed by Microcystis and then Dolichospermum.
Our results also suggested that cyanobacteria have a range in which critical damage, which we define as damage to more than 50% of the population, occurs. This is most apparent in the response of Microcystis to chlorine as there is high experimental variation between 1 and 1.5 mg-OCl−/L. The percentage of remaining phycocyanin after 1 mg-OCl−/L ranged from 21% to 65%, indicating that this dosage of chlorine may not always cause critical damage. Biological characteristics may contribute to the range in oxidative response at a given dosage. For example, cell viability, which can be measured by photosynthetic capacity, may dictate how susceptible a culture is to oxidation (Li et al., 2020; Zhou et al., 2013), but it was not tracked in our study. Our results suggest that Microcystis has a critical range between 1 and 2 mg-OCl−/L. Planktothrix appears to have a range of 0.5–1 mg-OCl−/L. The range for Dolichospermum starts at 1 mg-OCl−/L but may go beyond 2 mg-OCl−/L because we did not observe damage to 90% or more of the Dolichospermum population at 2 mg-OCl−/L.
The responses to chlorine oxidation correspond well with others reported in literature (Li et al., 2020; Ma et al., 2012; Qi et al., 2016), although there are slight variations in sensitivity by cyanobacterium type.
We found that permanganate oxidation is less damaging to cyanobacterial cells than chlorine at low dosages and short contact times. These observations support the guidance provided by the Ten States Standards (GLUMRB, 2022). We first examined permanganate within a contact time of 30 min. At applied permanganate concentrations up to the maximum recommended dosage of 1 mg-MnO4−/L, cells showed only a slight difference in remaining phycocyanin compared with the 0 mg-MnO4−/L control. Permanganate concentrations were extended to higher dosages to examine oxidant regimes under which cell damage might occur within 30 min. We found that Dolichospermum cells sustained critical damage at high dosages, but Microcystis and Planktothrix sustained only minimal damage at permanganate concentrations up to 20 mg-MnO4−/L (Fig. 1). Remaining intracellular phycocyanin levels decreased to 60% ± 16%, 80% ± 5%, and 38% ± 3% for Microcystis, Planktothrix, and Dolichospermum, respectively, compared with untreated controls, after exposure to a dosage of 20 mg-MnO4−/L for 30 min.
The observed damage to Planktothrix was not statistically significant compared with the control dosage of 0 mg-MnO4−/L, but those of Microcystis and Dolichospermum were. We see that cyanobacteria respond differently to chlorine and permanganate during the same contact time. Li et al. (2021, 2020) evaluated Microcystis aeruginosa cells and found that the rate of membrane damage due to chlorine (410 M−1s−1 at 2 mg-OCl−/L) was faster than the membrane damage rate by permanganate (27 M−1s−1 at 2 mg-MnO4−/L). Reaction rates likely explain why chlorine caused critical damage to our cell populations within a 30-min contact time while permanganate did not.
Given the lack of observed cell damage by permanganate at typical dosages, we examined the effect of longer contact times on Microcystis and Planktothrix (Fig. 2). We found that critical damage occurred at high dosages during a contact time of 2 h or more. A dosage of 10 mg-MnO4−/L for 4 h led to a decrease in remaining intracellular phycocyanin to 17% ± 6% and 3% ± 0.6% for Microcystis and Planktothrix, respectively. These responses indicate that Microcystis shows less sensitivity to permanganate than Planktothrix. Microcystis showed higher resistance to the permanganate, even at longer contact times, as critical damage only occurs at a dosage of 10 mg-MnO4−/L after 2 h. Planktothrix showed considerable, but not critical, damage from a dosage of 5 mg-MnO4−/L at a contact time of 1 h.
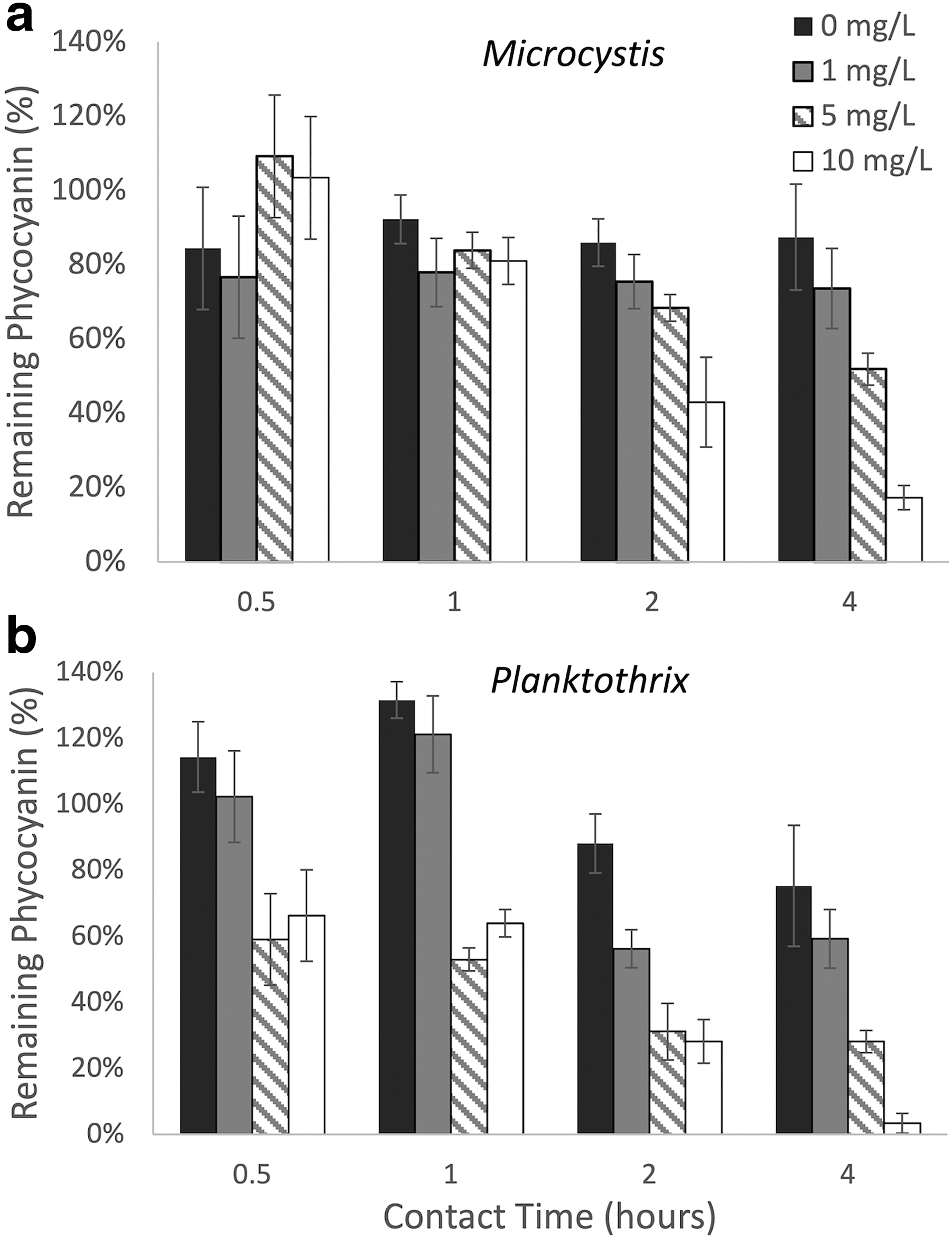
Effect of increased contact time on permanganate oxidation of
Notably, in these trials we see more damage after 30 min to Planktothrix for 5 and 10 mg-MnO4−/L samples than what was observed in the initial experiments (Fig. 1e), but still not critical damage until 2 h at a dosage of 5 mg-MnO4−/L. We also observe that Planktothrix reacts to experimental conditions, as the remaining phycocyanin in the 0 mg-MnO4−/L control also showed a decrease over the 4-h experiment (Fig. 2b).
Overall, our findings indicate that contact time is an important factor for permanganate damage to cells.
Across the three cyanobacteria, our findings correspond with findings reported by other studies. We found that 17% ± 6% of phycocyanin remained in Microcystis cells after 4 h of exposure to 10 mg-MnO4−/L, which is consistent with Fan et al. (2013) who reported that less than 20% of cells remained intact under the same oxidation conditions. Our findings are also consistent with those reported by Xu et al. (2019), who studied the effect of permanganate on Pseudanabaena galeata, a filamentous cyanobacteria similar to Dolichospermum. Their study found that only 75% of Pseudanabaena cells remained after 4 mg-MnO4−/L for 30 min, which aligns closely with our finding that 77 ± 10% of intracellular phycocyanin from Dolichospermum cells remained at 5 mg-MnO4−/L within the same contact time. Our results suggest that Dolichospermum is most sensitive to oxidative attack by permanganate, followed by Planktothrix then Microcystis, but our results may reflect slight experimental variations, such as lower cell density in Dolichospermum samples.
Overall, there is consistency between our measurements of cell damage by intracellular phycocyanin and of similar studies that employed direct measures of cell counts for assessing the impacts of chlorine or permanganate oxidation (Fan et al., 2013; Ma et al., 2012; Qi et al., 2016; Wert et al., 2013; Xu et al., 2019). Many prior studies, which quantified cell damage used cell membrane staining methods to obtain a ratio of live to dead cells (Ding et al., 2010; Fan et al., 2013; Wert et al., 2013). We analyzed several initial oxidation experiments with both live/dead staining and intracellular phycocyanin analysis. Our results demonstrated that intracellular phycocyanin analysis tends to indicate slightly more living cells than given by the live/dead stain analysis for the same sample (Supplementary Fig. S3 in the Supplementary Data). This suggests that cells may become damaged without releasing intracellular components (Ding et al., 2010). We concluded that the measure of intracellular phycocyanin is a good proxy for the number of whole cells in a population, but it yields an overestimation, compared with methods that can quantify mild membrane damage.
Shear damage on healthy cells
We next examined the effects of shear on cells with no exposure to preceding chemical oxidation. Our results showed that shear alone does not lead to immediate release of phycocyanin from cells. We found that exposure to 2 min of shear at a velocity gradient of 750 s−1 increased the remaining phycocyanin in Microcystis cells (Fig. 3). There was also an apparent increase in remaining phycocyanin for Planktothrix and Dolichospermum, although it was not statistically significant. A decrease in remaining phycocyanin after shear exposure suggests that shear is damaging cells and causing immediate release of phycocyanin. The absence of a change in the remaining intracellular phycocyanin suggests that damage that causes immediate release of cellular components is not occurring. An increase in remaining phycocyanin suggests either that there are more cells present or that increased phycocyanin production, or expression is occurring in existing cells.

Effects of shear conditions on cells with no exposure to preceding chemical oxidation.
We suspect that the apparent increase in remaining phycocyanin may be due to a cell stress response brought on by mechanical shear. Studies have reported that cyanobacteria can increase photosynthetic performance as a response to stress (Jin et al., 2020). It is feasible that an increase in phycocyanin could also be observed because it is involved in photosynthetic performance (Hazra and Saha Kesh, 2017). Although changes in phycocyanin levels in response to stress is unstudied, we question whether the apparent increase in phycocyanin expression following shear may also be a cell stress response. No study has reported a stress response to mechanical shear, but many studies have reported complex ways that cyanobacteria respond to a variety of other stressors, such as osmotic, oxidative, and temperature-induced stress (Jin et al., 2020; Rachedi et al., 2020; Yang et al., 2020; Zorina et al., 2011).
We note that there may also be evidence of stress response in the results of chemical oxidation on cells. We observe multiple instances where a low dosage of oxidant caused an increase in remaining intracellular phycocyanin, compared with the untreated control. For example, there is an increase in remaining phycocyanin after Dolichospermum was exposed to 1 mg-MnO4−/L, which was not observed at higher dosages (Fig. 1f).
We also observed a similar trend when Planktothrix was exposed to permanganate at long contact times (Fig. 2b). Within the initial hour of Planktothrix exposure to permanganate at low or zero dosages, the levels of remaining phycocyanin increased compared with the starting level. At 1 mg-MnO4−/L, the remaining phycocyanin increased from a starting level of 100% to 121% ± 6% at 1 h. At times >2 h, remaining phycocyanin decreased to 56% ± 9% or more. A similar trend was seen in the samples exposed to 0 mg-MnO4−/L. This may be an indicator of a stress response from cells, triggered by low dosages of oxidants or the stress from experimental conditions. The increase is not observed in higher dosages (above 5 mg-MnO4−/L), which show an overall dominant effect of cell damage.
The same trend was not observed for Planktothrix in the initial oxidation experiments, potentially due to the shorter contact time.
Our study is the first to report a potential stress response to mechanical shear, which may be due to the use of intracellular phycocyanin as the approach for evaluating cell integrity. Many studies that report effects of shear stress on cyanobacteria use cell staining to evaluate membrane integrity (Clemente et al., 2020; Oliveira et al., 2021; Sun et al., 2012). For example, Oliveira et al. (2021) reported membrane damage of both Microcystis and Dolichospermum cells but did not include analysis of intracellular organic matters, such as phycocyanin. Our study reports either an increase or no change in remaining phycocyanin, either way indicating no release of intracellular material during or immediately after shear. Considered together, Oliveira et al. (2021) and our study may support the notion that shear causes cell membrane damage such that they are susceptible to a viability stain, but not to an extent that cells immediately release intracellular components.
Shear damage on preoxidized cells
To determine whether preoxidation increases susceptibility to shear, we investigated the effect of shear with preceding exposure to either chlorine or permanganate. We found that chlorine preoxidation led to a decrease in remaining phycocyanin for Dolichospermum (Fig. 1c). The decrease of remaining phycocyanin from Dolichospermum cells indicates that cells were damaged by shear and released phycocyanin. This finding also suggests that chlorine preoxidation increases susceptibility to shear in Dolichospermum cells, since we did not report damage from shear on healthy Dolichospermum cells. In contrast, we observed an increase in remaining phycocyanin in Microcystis and Planktothrix (Fig. 1a, b). The increase in remaining phycocyanin from Microcystis and Planktothrix cells again implies a stress response due to shear but does not directly indicate that preoxidation increased susceptibility to shear.
In the case of permanganate preoxidation, we found that shear caused a decrease in remaining phycocyanin for Planktothrix but an increase for Microcystis and Dolichospermum (Fig. 1). This finding indicates that permanganate preoxidation makes Planktothrix more vulnerable to shear. While Dolichospermum was made more vulnerable to shear by chlorine, it does not display the same sensitivity to permanganate. Microcystis maintained similar responses regardless of preoxidation, indicating no increased susceptibility to shear from preoxidation.
Environmental implications
Generally, the potential for preoxidation to increase cyanobacteria susceptibility to shear depends on both oxidant type and cyanobacteria genera. It appears that an oxidant that is damaging to cells may not necessarily increase cell susceptibility to shear. For example, we noted that Dolichospermum was not made more vulnerable to shear by permanganate, even though Dolichospermum showed higher sensitivity to permanganate oxidation than the other cyanobacteria. Conversely, Dolichospermum was the only cyanobacteria to show higher shear susceptibility after chlorine exposure, yet this genus was more resistant to chlorine oxidation than the other cyanobacteria. A similar trend occurred with Planktothrix, which showed little damage by permanganate oxidation within a 30-min contact time for any oxidant dosages, but Planktothrix did show increased vulnerability to shear from 30 min of permanganate exposure. There are some cases in which preoxidation increases susceptibility to shear, but the effects of shear remain minimal overall.
There is no observation of shear causing critical damage, with or without preoxidation, which indicates that shear does not pose a risk for immediate release of cellular components, such as toxins or taste and odor compounds, under conditions typical of a rapid mixer. There may be, however, a concern for delayed release of cellular components downstream from shear exposure, despite no immediate critical damage to cells.
Cyanobacteria responses to oxidation and shear were somewhat varied but generally indicated that Microcystis cells are less susceptible to chemical and mechanical stressors compared with Planktothrix and Dolichospermum. Microcystis showed high resistance to chlorine and permanganate and did not show particular susceptibility to shear conditions, with or without preceding oxidation. Of the few studies, which compare the effects of stressors on multiple genera of cyanobacteria, there is conflicting information about which genera are the most sensitive. Wert et al. (2013) examined the effects of oxidants on Microcystis and two filamentous cyanobacteria. They found that Microcystis was more susceptible to oxidation than the filamentous cyanobacteria, which were of the same order as Planktothrix. Their finding is conflicting with ours, as we observed that Microcysitis was generally less susceptible than Planktothrix.
On the other hand, our findings match those of Mucci et al. (2017), who examined the effects of chitosan as a coagulant on Microcystis, Planktothrix, and Cylindrospermopsis cells. Cylindrospermopsis is a chained filamentous cyanobacteria of family Aphanizomenonaceae, like Dolichospermum. The study found that Microcystis was less sensitive to the chemical than Planktothrix and Cylindrospermopsis, likely due to the unique makeup of polymeric substances around their membrane (Mucci et al., 2017). However, it is important to note that chitosan is not an oxidative chemical, so effects of coagulation may have also been observed. Our finding also matches that of Oliveira et al. (2021), which examined the effects of shear conditions but not oxidative damage. Their study reports that Microcystis is less susceptible to shear than filamentous cyanobacteria, including Dolichospermum.
It is difficult to classify cyanobacteria by susceptibility when they have shown a range of responses to different stressors. The complexity associated with chemical damage is likely due to the exact reaction mechanism between a chemical and the substances that each cyanobacterium has in and around its membrane, such as polysaccharides (Oliveira et al., 2021) or polymeric substances (Mucci et al., 2017). This also relates to the effects of contact time, as chemical reaction rates will depend on the type of chemical and other reaction constituents that vary between cyanobacteria type, as we observed between chlorine and permanganate. Chemical composition of the membrane may also play a role in shear susceptibility, as characteristics such as toughness of the cell wall can impact response to physical stressors like shear. Thus, drinking water utilities have unique challenges in avoiding cyanobacteria damage because bloom conditions and treatment methods vary between treatment processes.
This study offers a technique that utilities can use to independently evaluate real-time cell response to treatment processes, especially as bloom and treatment conditions change. The intracellular phycocyanin analysis requires relatively accessible materials and analytical equipment, which are often available in a standard water quality testing laboratory. In-house testing can provide additional operational guidance when dealing with cyanobacterial blooms, especially as we consider the issue of delayed cellular component release. Each treatment process varies in terms of chemical dosages and contact times and residence times within treatment components, so the risk and timeline of cell damage is different between processes. The bloom conditions, such as dominating cyanobacteria species, also impacts risk of cell damage, and bloom conditions can change frequently over a season. The most direct way for utilities to avoid cell damage and release of cellular components would be to evaluate their own processes as blooms occur using bench-scale tests.
Aside from independent monitoring, there are operational precautions that utilities can take to protect the water against delayed metabolite release from cyanobacterial cells. During bloom events, utilities should ensure that cells are not left in the sedimentation basin for long periods of time, particularly if cells experienced a preoxidant before shear conditions. If cells are retained on filters, increased backwashing may be necessary to remove the cells before they release metabolites. Utilities should use caution when recycling filter backwash water because cells retained on the filter will likely be weakened further and could have released cellular components into the backwash water. We recommend that utilities remove as many cells as possible before filtration so that cell-containing waste can be fully removed from the process. Cell-containing waste from the settling basin could be a source of highly concentrated weakened cells, but it can be managed separately from the water treatment process. Utilities should use caution before reintroducing any recovered water from the waste into the system.
This study has demonstrated that shear preceded by preoxidation does not necessarily result in immediate release of intracellular components, but the long-term effects are not fully understood. Thus, the question of whether cells become stressed and release metabolites in the treatment process shifts to release of metabolites in the management of generated waste products.
Conclusions
The objective of this study was to determine whether chemical preoxidation increased cell susceptibility to mechanical shear. The study found the following:
Chlorine slightly increases susceptibility to shear conditions for Dolichospermum. Permanganate slightly increases susceptibility to shear conditions for Planktothrix. Cyanobacteria appear to show a stress response to mild stressors such as low oxidant dosages, changes in environmental conditions, and shear conditions, which does not result in immediate release of intracellular phycocyanin. The lack of immediate phycocyanin release likely indicates that immediate release of cyanotoxins and other cellular components does not occur. If cyanobacteria are removed from the treatment process shortly after exposure to mild stressors, the risk of cellular component release due to shear is likely minimal. Utilities can employ the methods used in this study to evaluate their treatment processes for likelihood to impart cell damage.
Footnotes
Acknowledgments
The authors thank Mr. Nicholas Dugan at the U.S. Environmental Protection Agency, Water Infrastructure Division for providing the Microcystis strain used in this study; Ms. Keara Stanislawczyk at The Ohio State University Stone Laboratory for processing Flowcam samples; and Dr. Elizabeth Crafton for assisting with cyanobacteria culturing methods. The authors also thank the two anonymous reviewers who helped strengthen this article with their comments.
Authors' Contributions
D.E.: methodology, investigation, formal analysis, validation, visualization, and writing—original draft. A.M.: conceptualization, writing—review and editing, supervision, project administration, and funding acquisition.
Author Disclosure Statement
The authors have no competing interests to declare.
Funding Information
Funding for this study was provided by the Ohio Water Development Authority through Research and Development Grant number AWD9165.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.