
Abstract
Excessive nitrogen (N) deposition alters plant–soil interactions and biogeochemical processes in forest ecosystems. Fine roots, which function as critical interfaces between plants and soil, exhibit morphological, anatomical, and mycorrhizal plasticity in response to nitrogen availability. This study examined species- and root-order-specific responses to nitrogen addition across a gradient and evaluated their ecological implications. The research, conducted in northeastern China’s Fenglin National Nature Reserve, investigated three tree species across plots subjected to four N treatments. Root samples were analyzed for morphological traits, anatomical traits, and mycorrhizal characteristics. General linear models and three-way analysis of variance were employed to assess the effects of N treatments, tree species, and root orders. Nitrogen addition significantly influenced fine root morphology, with high-N (TH) increasing root diameter and medium-N (TM) enhancing specific root surface area. Anatomical traits exhibited dose-dependent changes, including increased cortex thickness and stele diameter under TH. Mycorrhizal traits showed divergent responses, with low-N promoting colonization and total hyphae length, while TM enhanced hyphae diameter. Species-specific and root-order-specific variations underscored the complexity of belowground functional responses. Fine root and mycorrhizal adaptations to nitrogen addition reflect trade-offs between structural reinforcement and absorptive efficiency, driven by species identity, root developmental stage, and soil nutrient status. These findings provide critical insights into nitrogen-induced root plasticity and its role in shaping forest ecosystem processes, offering valuable perspectives for sustainable forest management under global change scenarios.
Introduction
Excessive nitrogen (N) deposition, typically defined as nitrogen inputs exceeding 10–20 kg N·ha−1·yr−1, has become a pressing concern in forest ecosystems worldwide, driven by industrial activities and agricultural intensification (Trebak et al., 2010). When nitrogen inputs surpass this threshold, forest ecosystems may undergo nitrogen saturation, leading to alterations in biogeochemical cycles and plant–soil interactions. In such ecosystems, fine roots and their symbiotic associations play a critical role in regulating water and nutrient uptake, carbon allocation, and overall plant fitness (De Vries et al., 2009). Despite numerous studies examining aboveground dynamics, our understanding of how varying levels of nitrogen addition alter belowground traits, including fine root morphology, anatomy, and mycorrhizal structures, remains incomplete (Smith et al., 2015). Elucidating these responses is essential, as roots are the primary interface between plants and soil, mediating resource acquisition and influencing soil biogeochemical processes (Wu et al., 2025).
Recent studies have demonstrated that fine roots exhibit substantial morphological plasticity underchanging nutrient conditions, adjusting traits such as root diameter (RD), specific root length (SRL), and specific root surface area (SRSA) to optimize resource foraging (Nadelhoffer, 2000). Recent findings further suggest that high nitrogen (TH) levels generally promote root thickening and reduce SRL, whereas moderate nitrogen levels can enhance root branching and surface area to maximize nutrient uptake efficiency (Ostonen et al., 2007). Similarly, anatomical features such as cortex thickness (CT) and stele diameter (SD) have been shown to respond dynamically to soil nitrogen availability, influencing root longevity and transport efficiency (Gan et al., 2021). For instance, a study in temperate forests found that excessive nitrogen inputs increased cortical thickness, potentially as an adaptive mechanism to enhance structural stability under nutrient-rich conditions (Wang et al., 2022). Furthermore, mycorrhizal associations are known to modulate plant nutrient uptake by extending the effective absorptive surface and facilitating symbiotic exchange (Campo et al., 2020). However, recent research indicates that mycorrhizal colonization can decline under TH conditions due to host plants shifting carbon allocation away from symbiotic fungi (Garcia et al., 2008). Yet, the exact ways in which distinct N addition treatments—ranging from low to moderate and high—shape these belowground traits, and how species-specific strategies modulate these outcomes, remain poorly understood. Additionally, interspecific variations in nitrogen-induced root responses highlight the necessity for species- and root-order-specific investigations (Kumar et al., 2020).
This gap in knowledge arises partly from the complex interplay among plant species, root developmental stages, and soil fertility conditions (Yetgin, 2024). While some research has highlighted that increased nitrogen often leads to thicker roots, higher SDs, and reduced mycorrhizal colonization due to altered carbon allocation patterns, few studies have separately examined the relationships between nitrogen addition and each of these traits—fine root morphology, anatomical structure, and mycorrhizal characteristics—across multiple nitrogen levels (Wada et al., 2019). Such insights are pivotal for predicting plant performance under scenarios of global change and for informing sustainable forest management strategies (Jackson et al., 2000).
Building upon these considerations, we formulated three hypotheses based on previous findings and theoretical expectations: (1) Nitrogen addition significantly affects the morphological characteristics of fine roots. TH levels are expected to increase RD due to enhanced carbon allocation to structural reinforcement (Yu et al., 2018). Medium nitrogen levels may optimize nutrient uptake by promoting finer roots with greater SRSA, as observed in previous studies on root plasticity in response to nutrient gradients (Ostonen et al., 2017). Low nitrogen (TL) conditions are likely to maintain higher SRL, favoring resource exploration over structural reinforcement (Chen et al., 2018). (2) Nitrogen addition significantly influences fine root anatomical traits. TH levels have been shown to promote cortex thickening and stele expansion, possibly as a strategy to enhance root longevity and nutrient transport efficiency under nutrient-rich conditions (Eissenstat et al., 2000). Medium nitrogen levels may increase the stele-to-diameter ratio (SDR), facilitating vascular efficiency in species with moderate nitrogen demands (Nacry et al., 2013). (3) Nitrogen addition significantly alters mycorrhizal morphology. Under TL conditions, mycorrhizal colonization and total hyphae length (THL) are expected to increase due to greater host carbon investment in symbiotic fungi to enhance nutrient foraging (Pena et al., 2023). Medium nitrogen levels may instead promote hyphal diameter and total hyphal surface area, as fungal networks optimize nutrient exchange in environments with moderate nitrogen availability (Parrent and Vilgalys, 2007).
Accordingly, the objectives of this study were to (i) clarify how fine root morphological and anatomical traits respond to gradient levels of nitrogen addition, (ii) elucidate species- and root-order-specific adjustments in these traits, and (iii) determine how mycorrhizal morphological characteristics change under different N treatments. By integrating morphological, anatomical, and mycorrhizal parameters, this research provides novel insights into the adaptive strategies of tree species within heterogeneous forest environments and contributes to a deeper mechanistic understanding of nitrogen-driven changes in belowground ecosystems.
Materials and Methods
Study sites and experimental design
The investigation was carried out in Fenglin National Nature Reserve, located in northeastern China’s Heilongjiang Province, within the administrative boundaries of Yichun City (48°02′–48°12′ N, 128°58′–129°15′ E). This reserve represents a typical temperate mixed forest ecosystem, dominated by Pinus koraiensis, Abies nephrolepis, and Acer tegmentosum, and has been widely studied in relation to forest dynamics and nutrient cycling (Sheng et al., 2011).
The reserve experiences a temperate continental monsoon climate, characterized by cold winters and brief but warm summers, with an annual precipitation of ∼650 mm, most of which falls between June and August. The mean annual temperature is around 1.8°C, and the elevation ranges from 230 to 560 m above sea level (Jobbágy and Jackson, 2000). These climatic conditions are representative of the broader temperate forest regions of Northeast Asia and have been shown to influence fine root traits and mycorrhizal associations in previous studies (Mridu et al., 2024).
The soil in the study area is classified as dark brown forest soil, with a pH of 6.5 and a bulk density of 1.3 g/cm³, supporting a rich variety of plant species (Dastager et al., 2009). Previous studies have highlighted that this soil type provides a stable yet nitrogen-limited environment, which can significantly shape root morphological plasticity and mycorrhizal symbiosis under nitrogen addition scenarios (Johnson et al., 2010).
The forest ecosystem in this reserve supports high biodiversity, comprising >120 tree species. Canopy coverage approaches 85%, providing a dense habitat for understory vegetation that includes about 35 shrub species and 20 herbaceous species. The study targeted three dominant tree species in the area: P. koraiensis, A. nephrolepis, and A. tegmentosum.
To evaluate the ecological impacts of nitrogen deposition, twelve 20 × 20 m experimental plots were set up on a forested slope with an eastern aspect. Key criteria such as vegetation structure, slope gradient, and orientation were considered when selecting the plots (Fig. 1). A buffer zone of 10 m separated adjacent plots to mitigate nitrogen contamination between treatments. Nitrogen was supplied in four treatments: control (CK, 0 kg N·ha−1·yr−1), TL (25 kg N·ha−1·yr−1), medium-N (TM, 50 kg N·ha−1·yr−1), and TH (75 kg N·ha−1·yr−1), with each treatment replicated three times. These nitrogen addition levels were selected based on previous studies on nitrogen deposition effects in temperate forests and their relevance to global nitrogen deposition scenarios (Wang et al., 2017).
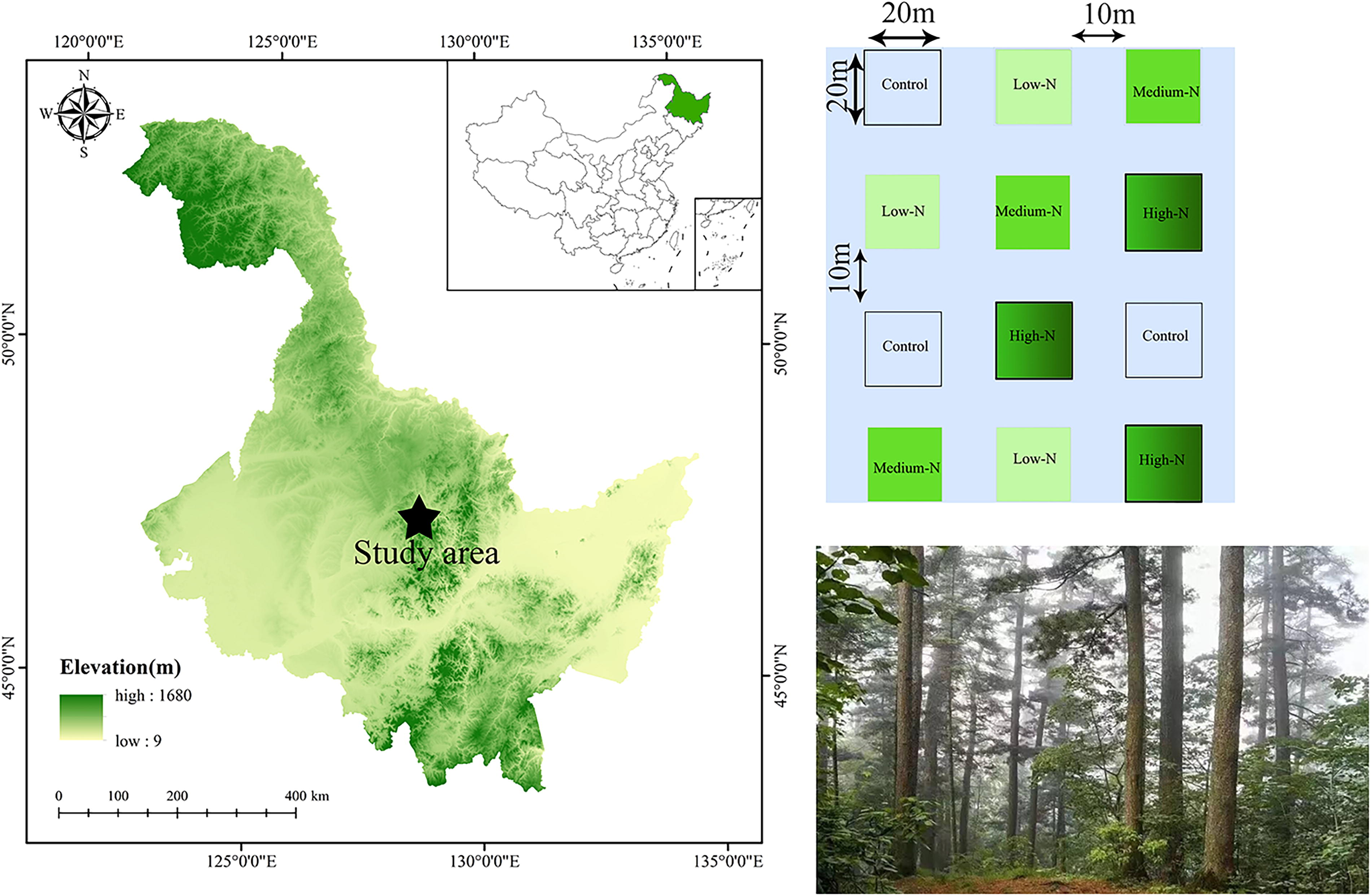
Sample plot and diagram of the experimental design in Fenglin National Nature Reserve, Yichun City, Heilongjiang Province, northeastern China.
The TL treatment (25 kg N·ha−1·yr−1) represents a moderate increase in atmospheric nitrogen deposition, which aligns with reported deposition rates in many temperate forest regions of China and North America (Zhang et al., 2012). The TM treatment (50 kg N·ha−1·yr−1) simulates an enriched nitrogen input scenario, reflecting potential future increases in nitrogen deposition due to industrial and agricultural activities (Liu et al., 2022). The TH treatment (75 kg N·ha−1·yr−1) was designed to examine potential nitrogen saturation effects, as previous studies have suggested that nitrogen saturation thresholds in temperate forests range between 50 and 80 kg N·ha−1·yr−1 (Hales et al., 2009).
The nitrogen addition levels in this study (25, 50, and 75 kg N·ha−1·yr−1) were chosen to represent moderate to TH deposition scenarios relative to the widely accepted nitrogen saturation threshold of 10–20 kg N·ha−1·yr−1 (De Schrijver et al., 2008). This ensures that the study effectively captures the potential impacts of excessive nitrogen inputs on fine root traits and mycorrhizal associations.
The nitrogen addition experiment was conducted continuously for 12 years (2011–2023) to assess the long-term impacts of nitrogen deposition on fine root morphology, anatomical traits, and mycorrhizal characteristics. Monthly applications of NH4NO3 dissolved in 32 L of deionized water were performed using backpack sprayers from May to October each year, following standardized nitrogen enrichment protocols used in temperate forest ecosystems (Frey et al., 2004). Control plots received an equal volume of deionized water without nitrogen to maintain consistent soil moisture levels across all treatments.
Sample collection
Within each plot, three trees of the designated species were randomly selected to obtain representative data, resulting in a total of 36 sampled trees per species across all treatments (3 trees per plot × 3 replicate plots × 4 nitrogen treatments). This sampling approach has been widely used in forest ecology studies to balance replication and logistical feasibility while maintaining statistical power for detecting treatment effects (Block et al., 2001). To further enhance data robustness, root and soil samples from each tree were collected from three separate points within a 3-m radius and subsequently pooled to form a composite sample per tree. This method reduces within-tree variability and ensures that the obtained data represent the local belowground environment more effectively (Valverde‐Barrantes et al., 2013). The selection of three trees per plot has been shown to capture species-specific variability while maintaining sufficient replication for statistical analyses. Root samples were collected around each tree using a 10 cm diameter soil auger to a depth of 10 cm, which corresponds to the soil layer containing the majority of roots. Root samples from three cores were combined into a composite sample per tree. These samples were immediately placed in insulated containers with ice and transported to the laboratory within 3 h, where they were stored at −20°C for subsequent analyses of morphological and chemical traits (Table 1). Soil samples from the same cores were also collected and stored in similar conditions to analyze mycorrhizal traits (Table 1).
Symbols
CT, cortex thickness; MHD, mean hyphae diameter; MRC, mycorrhizal root colonization; RD, root diameter; SD, stele diameter; SDR, stele-to-diameter ratio; SRL, specific root length; SRSA, specific root surface area; THL, total hyphae length; THS, hyphae surface area.
In the laboratory, root samples were gently cleaned using deionized water to remove soil particles while preserving structural integrity. Visual and tactile criteria, such as elasticity, color, and texture, were employed to distinguish live roots from dead ones. The samples were then divided into two portions: one for assessing mycorrhizal colonization and the other for morphological analysis.
Morphological analysis
Roots were classified into orders 1–3 based on Fitter’s classification system (Freschet et al., 2021). Scans of the cleaned roots were obtained at a resolution of 600 dpi using an Epson 11000XL scanner, and morphological parameters—including root length, diameter, number, and surface area—were quantified using Regent Instruments root analysis software. The samples were then oven-dried at 75°C until reaching a constant weight. SRL (cm/g) was calculated by dividing total root length by dry weight, while SRSA (dm2/g) was derived by dividing root surface area by dry weight.
Root anatomy traits
Fine roots (first to third orders) reserved for anatomical analysis underwent a series of dehydration steps using ethanol concentrations of 70%, 85%, 95%, and 100%, followed by clearing in xylene and embedding in paraffin. Thin sections (8 μm) were prepared using a microtome, stained, and observed under a compound microscope. Root parameters such as cortical thickness, overall diameter, and SD were measured using WinRHIZO TronMF 2012 software. The stele-to-root diameter ratio was then calculated to analyze structural adaptations.
Mycelium extraction and morphological traits
To evaluate mycorrhizal infection, ∼200 root tips were selected randomly from each sample. Observations of macroscopic infection indicators, such as swelling and fungal structures, were made using a dissecting microscope. Infection rates were expressed as the percentage of infected tips relative to the total number observed.
Extraradical hyphae were isolated from 5 g of fresh soil following the method described by Toljander et al. (2006). Soil samples were homogenized in water using an electric blender, and the resulting suspension was filtered through a membrane. Hyphae were stained with lactoglycerol containing 0.05% trypan blue and examined under a microscope. Hyphal traits, including total length, surface area, and average diameter, were quantified using the same root analysis software.
Statistical analysis
The Kolmogorov–Smirnov test was used to assess data normality, while Levene’s test evaluated homogeneity of variance. If the data did not meet the normality assumption (p < 0.05), a nonparametric Kruskal–Wallis test was applied to compare differences among treatments, followed by Dunn’s post hoc test for pairwise comparisons. For datasets with unequal variances, a Welch’s analysis of variance (ANOVA) was performed instead of standard ANOVA to account for heteroscedasticity. A general linear model and three-way ANOVA were used to assess the effects of nitrogen treatments, tree species, and root order on various root and mycorrhizal traits. Pairwise comparisons were conducted using the Least Significant Difference method. All statistical analyses were performed in R version 3.2.3.
Results
Root morphological traits
The effects of different treatments on mean RD, SRL, and SRSA varied significantly among tree species and root orders (Fig. 2). For RD, TH significantly increased values compared to CK in most cases, particularly in higher root orders. Specifically, in P. koraiensis, RD was significantly higher under TM and TH compared to CK in second and third root orders (Fig. 2A). In A. nephrolepis, TH resulted in significantly higher RD across all root orders compared to other treatments (Fig. 2A). In A. tegmentosum, RD was significantly higher under TH in all root orders, with the most pronounced effect observed in third-order roots (Fig. 2A).
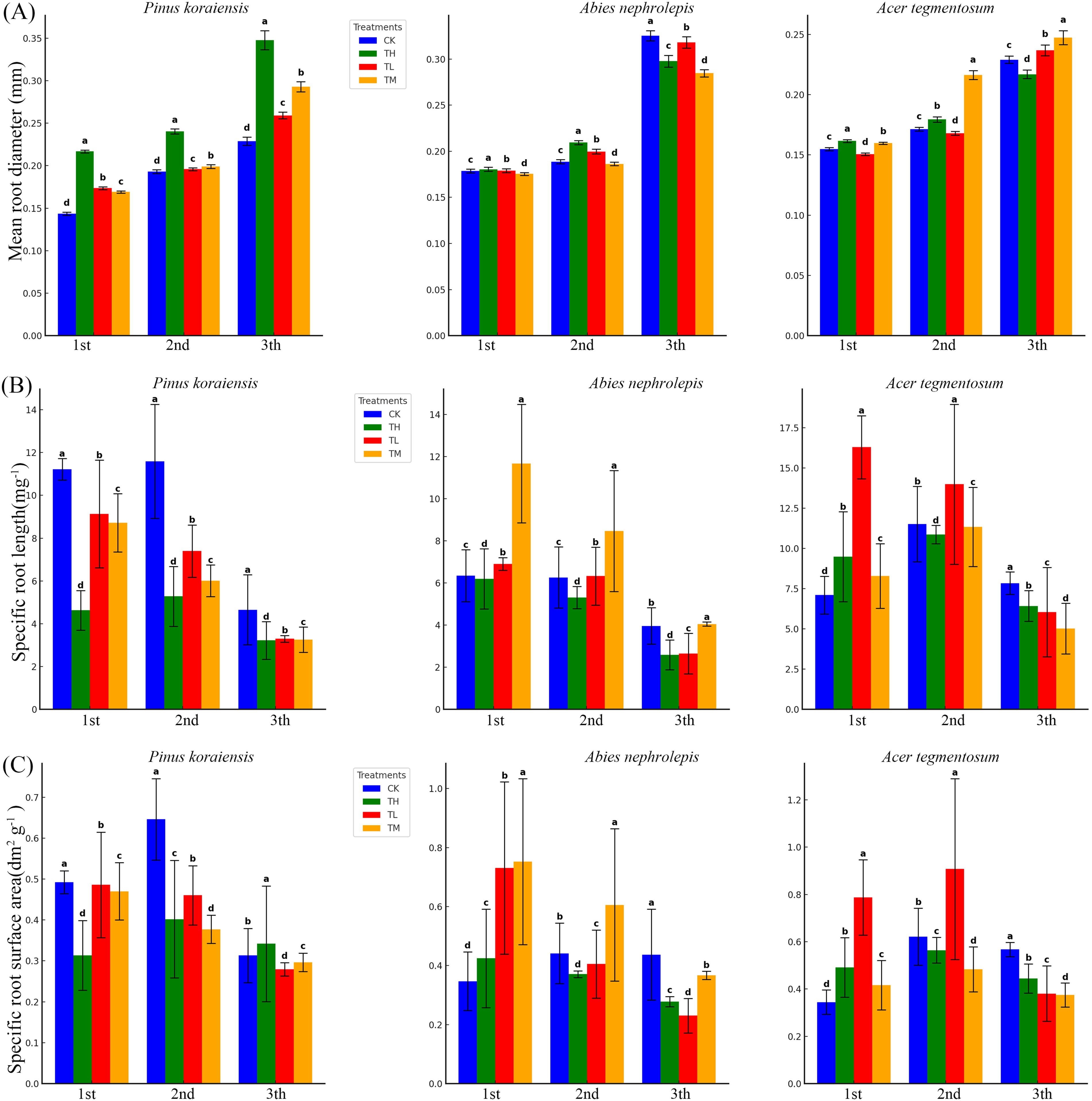
Mean root diameter
SRL exhibited contrasting patterns among species and root orders. In P. koraiensis, SRL under CK was significantly higher than under TM and TH in first-order roots (Fig. 2B). In A. tegmentosum, SRL was significantly higher under TM compared to CK in first- and second-order roots (Fig. 2B). In A. nephrolepis, SRL exhibited a mixed response, with significant increases under TM in some root orders but decreases under TH (Fig. 2B).
SRSA showed distinct trends depending on species and root orders. For P. koraiensis, SRSA in first-order roots was significantly higher under CK compared to TM and TH, while in third-order roots, TM resulted in significantly higher values (Fig. 2C). In A. nephrolepis, SRSA increased significantly under TM across all root orders compared to CK and TL (Fig. 2C). Similarly, in A. tegmentosum, TM consistently led to significantly higher SRSA compared to CK in second-order roots, while TH showed the most significant effect in third-order roots (Fig. 2C).
The ANOVA (Table 2) confirmed significant effects of tree species, treatments, root orders and their interactions on RD, SRL, and SRSA. These findings highlight the differential responses of fine root traits to nitrogen addition across tree species and root orders, emphasizing the complexity of belowground functional traits under varying environmental conditions (Fig. 2 and Table 2).
Analysis of Variance Results for Effects of Species, Treatments, Root Order, and Interactions on Fine Root Traits
Shown are degrees of freedom (df) and the p value of the respective variables and the model itself. S represents the species (Pinus koraiensis, Abies nephrolepis, and Acer tegmentosum); T represents the treatments (control, low-N [25 kg N·ha−1·yr−1], medium-N [50 kg N·ha−1·yr−1], high-N [75 kg N·ha−1·yr−1]); and R represents the root order (first- to third-order fine roots).
p < 0.001.
RD, root diameter; SRL, specific root length; SRSA, specific root surface area.
Root anatomy traits
The effects of different treatments on CT, SD, and SDR varied significantly among tree species and root orders (Fig. 3). For CT, TH resulted in significantly higher values compared to other treatments across all root orders in P. koraiensis (Fig. 3A). In A. nephrolepis, CT was significantly higher under TH compared to TM and TL in first- and second root orders, while in third-order roots, TM was significantly higher than CK. Similarly, in A. tegmentosum, TH significantly increased CT compared to TL, particularly in second and third root orders.

Cortex thickness
For SD, P. koraiensis showed significantly higher values under TH in first root order compared to CK and TL, while in second and third root orders, TM was significantly higher than other treatments (Fig. 3B). In A. nephrolepis, TH consistently resulted in significantly higher SD across all root orders compared to CK. In A. tegmentosum, SD was significantly higher under TM in second and third root orders compared to TL. For SDR, no significant differences were observed among treatments for P. koraiensis and A. nephrolepis across all root orders. However, in A. tegmentosum, SDR was significantly higher under TM compared to TL in second root order (Fig. 3C).
The ANOVA indicated that tree species, treatments, root orders, and their interactions had significant effects on CT, SD, and SDR (Table 3). These findings underscore the significant variations in fine root anatomical traits among different treatments, tree species, and root orders (Fig. 3 and Table 3).
Analysis of Variance Results for Effects of Species, Treatments, Root Order, and Interactions on Root Anatomy Traits
Shown are degrees of freedom (df) and the p value of the respective variables and the model itself. S represents the species (Pinus koraiensis, Abies nephrolepis, and Acer tegmentosum); T represents the treatments (control, low-N [25 kg N·ha−1·yr−1], medium-N [50 kg N·ha−1·yr−1], high-N [75 kg N·ha−1·yr−1]); R represents the root order (first- to third-order fine roots).
*p < 0.05.
p < 0.01.
p < 0.001.
CT, cortex thickness; SD, stele diameter; SDR, stele-to-diameter ratio.
Mycorrhizal morphological characteristics
The effects of different treatments on mycorrhizal root colonization (MRC), THL, total hyphae surface area (THS), and mean hyphae diameter (MHD) varied significantly among tree species (Fig. 4). For MRC, CK resulted in significantly higher colonization rates compared to TL and TM in P. koraiensis, while no significant differences were observed between CK and TH (Fig. 4A). In A. nephrolepis, colonization rates were significantly higher under CK compared to TM and TH. Similarly, in A. tegmentosum, CK and TL showed significantly higher colonization rates compared to TM, while no significant differences were observed between CK and TH.
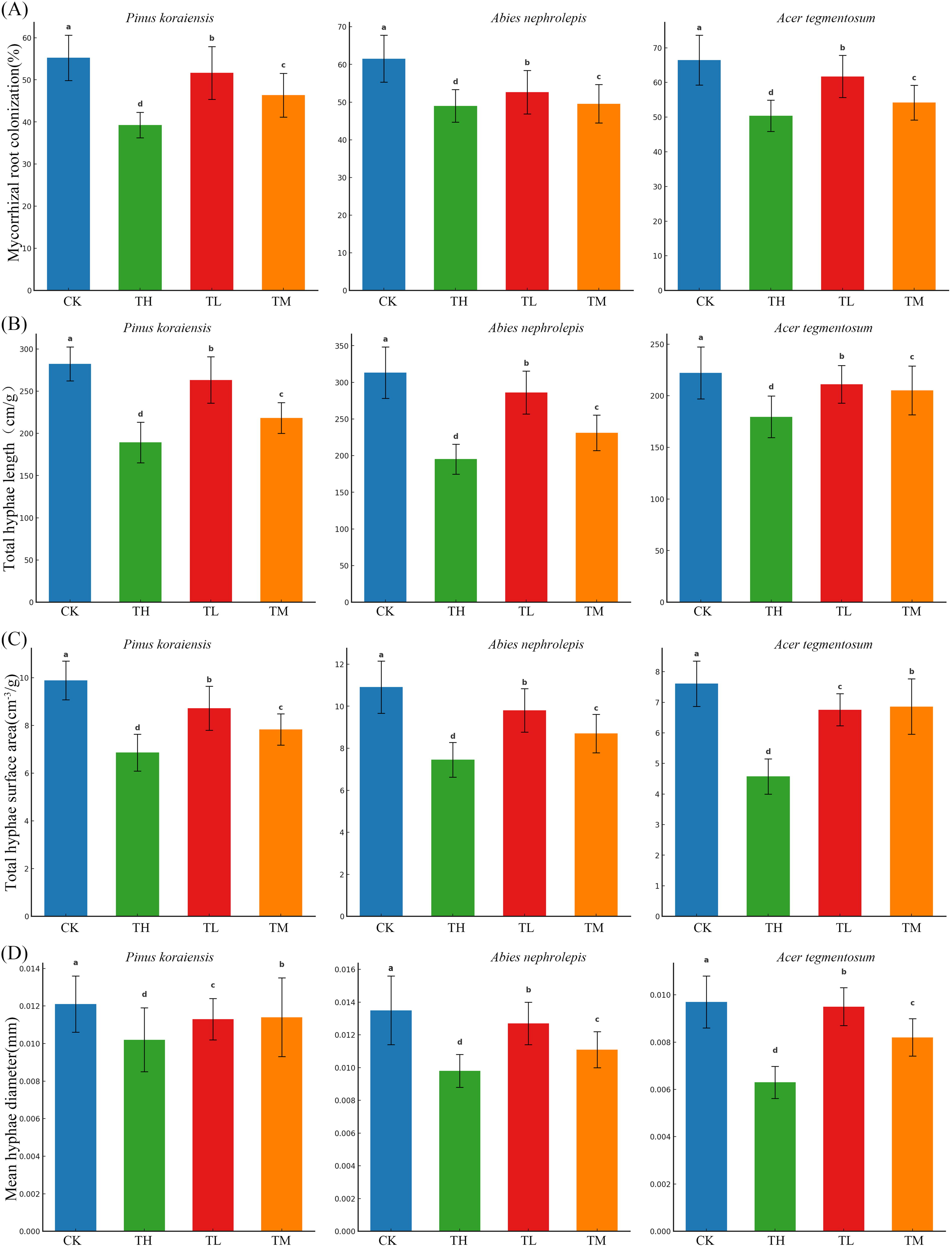
Mycorrhizal root colonization
For THL, P. koraiensis showed significantly higher THL under CK compared to all other treatments (Fig. 4B). In A. nephrolepis, TL resulted in significantly higher THL compared to TM and TH. In A. tegmentosum, CK produced significantly higher THL compared to TM, while TL was significantly higher than TH.
THS also exhibited significant differences among treatments (Fig. 4C). In P. koraiensis, CK resulted in significantly higher values compared to TL, TM, and TH. In A. nephrolepis, TL resulted in significantly higher THS compared to CK, while TM was significantly lower than TH. In A. tegmentosum, THS was significantly higher under CK compared to TM and TH.
For MHD, P. koraiensis showed significantly higher values under TM compared to CK and TH (Fig. 4D). In A. nephrolepis, TH resulted in significantly higher MHD compared to CK, while TL was significantly lower than all other treatments. In A. tegmentosum, TH resulted in significantly higher MHD compared to CK, and TM was significantly higher than TL.
The ANOVA confirmed significant effects of tree species, treatments, and their interactions on all measured parameters (Table 4). These results demonstrate the significant variability in mycorrhizal morphological characteristics among tree species and treatments (Fig. 4 and Table 4).
Analysis of Variance Results for Effects of Species, Treatments, Root Order, and Interactions on Mycorrhizal Morphological Characteristics
Shown are degrees of freedom (df) and the p value of the respective variables and the model itself. S represents the species (Pinus koraiensis, Abies nephrolepis, and Acer tegmentosum); T represents the treatments (control, low-N [25 kg N·ha−1·yr−1], medium-N [50 kg N·ha−1·yr−1], and high-N [75 kg N·ha−1·yr−1]).
p < 0.05.
p < 0.001.
MHD, mean hyphae diameter; MRS, mycorrhizal root colonization; THL, total hyphae length; THS, total hyphae surface area.
Discussion
Responses of fine root morphology to nitrogen addition
Our findings demonstrate that nitrogen addition significantly influenced the morphological traits of fine roots, including mean RD, SRL, and SRSA, with notable differences among tree species (P. koraiensis, A. nephrolepis, A. tegmentosum) and root orders (Fig. 2, Table 2). TH consistently increased RD across all species, particularly in higher root orders, whereas TL maintained higher SRL in P. koraiensis. TM enhanced SRSA in A. nephrolepis and A. tegmentosum, particularly in second-order roots, highlighting a species- and root-order-specific response to nitrogen addition (Fig. 2A–C).
These results demonstrate that nitrogen availability substantially alters the morphological traits of fine roots, reflecting a shift in resource allocation strategies. TH promoted structural reinforcement through increased RD, while TL maintained higher SRL, indicating enhanced absorptive efficiency under nitrogen-limiting conditions. The significant increase in SRSA under TM suggests a balanced response to optimize water and nutrient absorption, especially in A. nephrolepis. These findings align with the hypothesis that nitrogen addition modifies fine root morphology to maximize resource use efficiency.
In comparison to previous studies, our findings are consistent with Garnett et al. (2009), who reported increased RD and decreased SRL under TH conditions in temperate forest ecosystems. This alignment highlights a common physiological adaptation of fine roots to nitrogen enrichment, wherein thicker roots are favored to improve structural stability and reduce turnover rates. However, our observation of increased SRSA under TM diverges from Zhang et al. (2019), who found no significant changes in SRSA across nitrogen treatments. This discrepancy may arise from differences in experimental conditions, such as soil nitrogen saturation levels or tree species with distinct nutrient acquisition strategies (Van Sundert et al., 2020). Additionally, the species-specific response observed in our study, particularly the enhanced SRSA in A. tegmentosum, could be attributed to variations in root plasticity and symbiotic interactions with soil microbiota (Lee et al., 2014).
The results are largely consistent with our hypothesis. TH significantly increased RD, TM improved SRSA, and TL maintained higher SRL. These trends likely reflect a trade-off between structural reinforcement and absorptive efficiency under varying nitrogen conditions. The increased RD under TH can be explained by a shift in carbon allocation to root thickening, which enhances nutrient transport efficiency and reduces the metabolic cost associated with fine root turnover (Tabares et al., 2020). Similarly, the enhanced SRSA under TM may reflect a strategy to optimize water and nutrient acquisition in moderately enriched environments (Paez-Garcia et al., 2015). However, the reduced SRL under TH, particularly in P. koraiensis, deviates from our hypothesis, potentially due to species-specific limitations in root elongation under nitrogen-rich conditions or competitive interactions with soil microorganisms (Weemstra et al., 2021). A potential explanation for the observed reduction in SRL is that TH availability may shift carbon allocation from root elongation to structural reinforcement. Increased nitrogen levels often enhance shoot growth, potentially reducing the relative investment in fine root elongation as trees prioritize aboveground biomass accumulation (Tian et al., 2022). This reallocation could result in shorter, thicker roots with lower SRL, as observed in P. koraiensis. Additionally, nitrogen enrichment can alter soil microbial communities, leading to increased competition between roots and microbial decomposers for essential resources. TH inputs may promote the proliferation of fast-growing bacteria while suppressing mycorrhizal associations, thereby reducing the plant’s reliance on extensive root elongation for nutrient acquisition (Xun et al., 2015). Furthermore, root exudation patterns may change under TH, influencing rhizosphere interactions and potentially limiting root elongation (Kou et al., 2015). These results emphasize the importance of nitrogen availability and species-specific adaptations in shaping fine root morphology.
Recent studies highlight that nitrogen-driven changes in fine root morphology depend on species traits, soil nutrients, and microbial interactions. Long-term nitrogen enrichment may alter root lifespan and turnover, impacting nutrient cycling and soil carbon storage (Wilts et al., 2004). However, key questions remain: (1) Do these changes persist over time? (2) How do they influence rhizosphere microbes? (3) How do nitrogen and other global change factors interact? Addressing these will enhance understanding of plant–soil feedback and forest ecosystem resilience.
Impacts of nitrogen addition on fine root anatomical traits
The findings of this study demonstrate that nitrogen addition significantly influenced fine root anatomical traits, including CT, SD, and SDR, with notable variations among tree species and root orders (Fig. 3, Table 3). TH resulted in significantly higher CT and SD across most species and root orders, while TM led to increased SDR in specific tree species such as A. tegmentosum in the second root order (Fig. 3A–C). These observations underscore the differential responses of fine root anatomy to varying nitrogen levels.
These observations underscore that TH availability exerts a pronounced influence on root anatomical traits, particularly by thickening the cortex and broadening the stele, while TM stimulates proportional changes in internal structure, reflected by SDR. This pattern implies that fine root anatomical plasticity responds differentially to increasing N levels, presumably enabling species-specific optimization of water and nutrient transport capacity (Wang et al., 2018).
Consistent with the findings of Li et al. (2023), who observed increased CT and SD under TH in temperate forest species, our results suggest that enhanced nitrogen availability promotes root thickening and structural robustness. This similarity can be attributed to increased carbon allocation to root thickening under nutrient-enriched conditions, which strengthens roots and reduces their susceptibility to biotic and abiotic stresses (Lu et al., 2020). However, our observation of a significant increase in SDR under TM contrasts with the findings of Li et al. (2010), who reported no changes in SDR across nitrogen treatments in subtropical forest ecosystems. This discrepancy may arise from differences in experimental conditions, such as soil nitrogen saturation levels or species-specific responses to nitrogen addition. In particular, A. tegmentosum exhibited a pronounced increase in SDR under TM, potentially due to its reliance on fine-tuned root plasticity to optimize nutrient transport efficiency under moderate nutrient availability (Yan et al., 2018).
Our results align closely with the hypothesis that TH generally promotes cortex and stele expansion, whereas TM promotes a higher SDR in certain tree species and root orders. The reason for such congruence is that, under TH conditions, enhanced nitrogen assimilation can escalate the biosynthesis of structural compounds, thereby thickening root tissues (Krapp, 2015). Meanwhile, the phenomenon of increased SDR under TM likely reflects an intermediate nitrogen status that balances tissue differentiation and vascular capacity (Gadallah, 2000). However, partial divergence among species and root orders may arise from differences in root ontogeny, nutrient acquisition strategies, or preexisting soil fertility baselines (Tobner et al., 2013). Consequently, our findings confirm that nitrogen-induced anatomical plasticity varies with both species identity and root developmental stage, thereby offering novel insights into root functional adaptations in heterogeneous forest ecosystems.
Recent studies highlight that nitrogen-induced changes in root anatomy vary by species, root order, and environmental conditions, affecting water and nutrient transport efficiency (Wang et al., 2023). While TH promotes cortical thickening and stele expansion, moderate nitrogen influences vascular differentiation, yet long-term effects remain unclear. Key questions include (1) How do these anatomical changes persist over time? (2) What are the implications for root hydraulic conductivity? (3) How do multiple environmental factors interact with nitrogen to shape root anatomy? Addressing these will improve the understanding of plant adaptation to nitrogen enrichment.
Responses of mycorrhizal morphology to nitrogen addition
As shown in Figure 4 and supported by Table 4, CK consistently resulted in elevated MRC and THL across all three tree species, whereas TL and TM influenced these parameters in a species-specific manner. In addition, TM and TH produced significant increases in MHD for some species, although their effects on total THS remained variable (Fig. 4).
These findings indicate that TL treatments generally favor MRC and THL, whereas TM can enhance MHD. However, marked interspecific differences suggest that nitrogen-driven mycorrhizal morphological adjustments are highly dependent on each species’ physiological and ecological traits (Fig. 4 and Table 4). One possible explanation for the increased MRC and THL under TL is that TL availability promotes greater carbon allocation to mycorrhizal symbionts, enhancing fungal colonization and extraradical hyphal growth (Pan et al., 2020). Under nitrogen-limited conditions, plants rely more on mycorrhizal fungi to improve nutrient uptake efficiency, leading to higher colonization rates and more extensive hyphal networks. Conversely, TM may enhance MHD by facilitating a moderate nutrient supply that supports fungal structural reinforcement without suppressing symbiotic interactions. However, interspecific differences in mycorrhizal responses may arise from variations in host dependency on fungal symbiosis. Species with inherently high mycorrhizal reliance, such as A. nephrolepis, may maintain high MRC even under moderate nitrogen levels, while species with more flexible root foraging strategies, like P. koraiensis, may reduce symbiotic investment as soil nitrogen availability increases (Cahill and McNickle, 2011). Additionally, nitrogen enrichment can shift fungal community composition, favoring taxa that allocate more biomass to thicker hyphae rather than expanding the overall network length, which may explain the increased MHD under TM.
Comparable patterns have been reported in various forest stands subjected to changing nitrogen levels, where reduced nitrogen inputs tended to foster hyphal expansion, possibly due to improved carbon allocation from host to fungus (Bahr et al., 2013). Nonetheless, Yang et al. (2017) observed inconsistent outcomes, likely attributable to variability in edaphic conditions and symbiont assemblages. The principal explanation for such discrepancies resides in the differing capacities of mycorrhizae to adapt to nitrogen availability shaped by local soil fertility (Sherrard and Maherali, 2012).
In light of our hypothesis that nitrogen addition significantly alters mycorrhizal morphological traits, our results largely concur with the premise that TL fosters enhanced colonization and hyphal extension, while TM promotes thicker hyphae in certain species (Zhou et al., 2021). The mechanism underlying TL-induced colonization augmentation may stem from balanced carbon—nitrogen exchange, in which host trees direct more photosynthate toward fungal symbionts under TL (Cheeke et al., 2017). Likewise, the greater hyphal diameter triggered by TM could reflect metabolic shifts that prioritize structural reinforcement when nitrogen is moderately supplied (Yang et al., 2014). Although our observations corroborate these patterns, exceptions arose in A. nephrolepis, where TM and TH additions did not uniformly boost THS. One plausible reason is the species’ distinct nutrient uptake strategy, potentially shaped by root architecture or evolutionary adaptation to nutrient-poor environments (Zhang et al., 2023). Another explanatory factor may be the suppression of enzymatic functions under excess nitrogen, leading to divergent morphological outcomes (Enowashu et al., 2009). Overall, while our findings align with the hypothesis, the observed interspecific divergence underscores the necessity to consider inherent plant functional traits and soil nutrient status when interpreting nitrogen-induced mycorrhizal responses (Powell et al., 2009).
Recent studies highlight that nitrogen-driven shifts in mycorrhizal morphology are species-specific and influenced by soil conditions and fungal community dynamics (McKee, 2001). While TL enhances colonization, moderate nitrogen alters fungal structural investment, but long-term effects remain uncertain. Key questions include (1) How do nitrogen-induced mycorrhizal changes persist over time? (2) What are the functional consequences for nutrient cycling? (3) How do multiple environmental stressors interact with nitrogen to shape mycorrhizal dynamics? Addressing these will improve predictions of forest ecosystem responses to nitrogen deposition.
Conclusion
This study demonstrates that nitrogen addition significantly influences fine root morphology, anatomy, and mycorrhizal traits in a species- and root-order-specific manner. TH increased RD, while TL maintained higher SRL, and TM enhanced SRSA, highlighting trade-offs between structural reinforcement and absorptive efficiency. Anatomically, TH promoted cortex thickening and stele expansion, whereas TM increased the SDR in certain species, reflecting root structural adjustments to nitrogen enrichment. Mycorrhizal traits exhibited divergent responses, with TL increasing MRC and THL, while TM enhanced MHD, suggesting shifts in symbiotic investment strategies. These findings underscore nitrogen-driven plasticity in root traits and emphasize the necessity for future research to integrate root functional traits with ecosystem nutrient cycling. Understanding how species-specific adaptations mediate belowground responses to nitrogen deposition will improve predictions of forest ecosystem resilience and inform sustainable management strategies.
Footnotes
Acknowledgment
The authors gratefully appreciate the Chinese Academy of Sciences for the great cooperation in the experiment.
Authors’ Contributions
M.Z. designed the study, Y.L. got grants from the foundation, supervised data collection, and edited the article. M.Z., Y.L., and J.H. contributed to the whole article preparation and design and wrote the main article text. M.Z., J.H., P.Y., and T.Y. prepared all figures and field experiments. All authors reviewed the article.
Availability of Data and Material
Data are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This research was supported by grants from the Researchers Supporting Project New Era Longjiang excellent master or doctor dissertation grant program (LJYXL2023-060).