
Abstract
Introduction:
Swine models have proven useful for many different disease processes, especially for urologic research. In this study, we sought to create a model of hyperoxaluria in the adult sow by feeding hydroxyproline (HP). The development of an adult porcine model for calcium oxalate stone disease would represent a significant contribution to stone research as previous animal models have been developed only for rats and baby pigs.
Methods:
The experiment included a total of 12 multiparous, gestating sows (Large White × Landrace). Sows were randomly allotted to one of the two treatment groups. Treatments involved basal diets that were either control diet (CD) or acidogenic diet (AD). Urine was collected for 6 consecutive days. On days 1 and 2, each sow was fed 2 kg of the assigned basal diet (CD or AD). On days 3, 4, and 5, 200 g of L-hydroxyproline (Wilshire Technologies, Princeton, NJ) was added to each basal diet for all the 12 sows. The HP was evenly mixed with the basal diets before feeding. On day 6, each sow was fed the basal diet originally assigned without HP (Fig. 1). Urine was collected for each entire 24-hour period to control for differences in the diurnal and postprandial variations in the renal handling of oxalate and glycolate.
Results:
The addition of HP to the diet increased urinary oxalate excretion. Overall, there was a 192% (CD) and 187% (AD) increase in urinary oxalate between days 1 and 3. The increase peaked on day 3 and gradually returned to baseline by day 6. Student's paired t-test was performed and it confirmed that oxalate on days 3 and 5 was significantly different than baseline (p = 0.009 and p = 0.03, respectively). Urinary glycolate also increased as a result of adding HP to the diet. Overall, there was a 12,340% (CD) and 14,400% (AD) increase in urinary glycolate between days 1 and 3. The increase peaked on day 3 and then declined, although remained more than 10 × greater than baseline at day 6. Student's paired t-test confirmed that glycolate levels on days 3, 5, and 6 were significantly different than baseline (p < 0.001, p = 0.01, and p = 0.03, respectively).
Conclusion:
The role of oxalate in the formation of kidney stones cannot be understated. Medical prevention and management of calcium oxalate nephrolithiasis will require a comprehensive understanding of oxalate metabolism in humans. A model for human hyperoxaluria can be reliably created in the adult sow. Such a model is necessary to further our understanding of oxalate metabolism and ultimately aid in the prevention of calcium oxalate calculi.
Introduction
Oxalate metabolism is an area of research that remains of great interest. Urinary oxalate is believed to be derived equally from dietary and endogenous pools. However, pathways leading to endogenous synthesis of oxalate have not been fully elucidated. 3 From studies on types 1 and 2 primary hyperoxaluria, high levels of oxalate are known to result from deficiencies in the glycolate metabolizing enzymes alanine-glyoxylate aminotransferase and glycolate reductase. 4 Thus, glycolate is implicated as a precursor to endogenous oxalate. The endogenous precursors of glyoxylate and glycolate are to date not known, but hydroxyproline (HP) has been implicated as part of the pathway. 5
HP is metabolized to pyruvate and glycolate in both hepatic mitochondria and renal proximal tubular cells. 5,6 In humans, collagen is metabolized at a rate of 2–3 g daily, leading to metabolism of 240–420 mg of HP and formation of 140–240 mg of glycolate. 5
Hyperoxaluria has been induced in rat, porcine, and human models by various dietary manipulations. In the rat, oral intake of ethylene glycol or HP increased urinary oxalate. 7 Mandel et al 2 demonstrated that young growing pigs fed HP became hyperoxaluric, developed crystalluria, and developed calcium oxalate plaques on renal papillae. Knight et al 5 fed gelatin, a food ingredient that contains HP, to healthy human subjects and created elevated levels of urinary oxalate and glycolate. HP is a natural source of animal protein.
The western-type diet, which has an abundance of animal protein, has been implicated as an increased risk factor for the formation of kidney stones in humans. 8 Consumption of the western-type diet results in endogenous oxidation of excess cationic and sulfur-containing amino acids which impose a chronic metabolic acidosis that leads to bone demineralization and hypercalcuria. 9
In this study, we sought to create a model of hyperoxaluria in the adult sow by feeding HP. The development of an adult porcine model for calcium oxalate stone disease would represent a significant contribution to stone research as previous animal models have been developed only for rats 10 and baby pigs. 2 It is preferable to use the adult pig as a model because most research confirming anatomic and physiologic similarities with humans has been done in adult pigs. We hypothesized that adding HP to the diets of adult sows will increase the urinary excretion of both oxalate and glycolate. We tested this hypothesis in sows fed both a normal gestating diet, formulated to meet their nutritional needs, 13 and a higher acid load diet, similar to the typical western diet of adults in the United States.
Methods
The experiment included a total of 12 multiparous, gestating sows (Large White × Landrace). Sows were housed at the University of Wisconsin Swine Research and Teaching Center. The experimental protocol was approved by the University of Wisconsin–Madison Research Animal Resource Committee. Sows were randomly allotted to one of the two treatment groups. Treatments involved basal diets that were either control diet (CD) or acidogenic diet (AD). The CD diet consisted of the standard University of Wisconsin (UW) gestation diet, a corn soybean meal-based diet. The AD diet consisted of 2.5% addition of CAD-MATE to the UW gestation diet. CAD-MATE is a commercial source (Granco Minerals, Petersburg, VA) of a blend of inorganic anionic salts which provide excess sulfate and chloride relative to cations. Animals were fed 2 kg diet per day and allowed continuous access to water.
Each sow with catheterized with a 20F Bard Foley catheter. Balloon was inflated to 30 cm3, and Tygon tubing was attached to the catheter and placed in the cap of a 20-L collection pail. Collection pails were weighed before being placed in the ice-filled plastic tubs. Ice was used to stabilize urine collected over each 24-hour collection period rather than attempts to stabilize urine by acidification because urine pH was a critical measure. This procedure allowed accurate assessment of urine pH and cation–anion excretion in earlier experiments with young pigs. 11 Sows were closely monitored for signs and symptoms of cystitis, and they were removed from the trial in cases of infection.
Urine was collected for 6 consecutive days. On days 1 and 2, each sow was fed 2 kg of the assigned basal diet (CD or AD). On days 3, 4, and 5, 200 g of L-hydroxyproline (Wilshire Technologies, Princeton, NJ) was added to each basal diet for all the 12 sows. The HP was evenly mixed with the basal diets before feeding. On day 6, each sow was fed the basal diet originally assigned without HP (Fig. 1). Urine was collected for each entire 24-hour period to control for differences in the diurnal and postprandial variations in the renal handling of oxalate and glycolate.
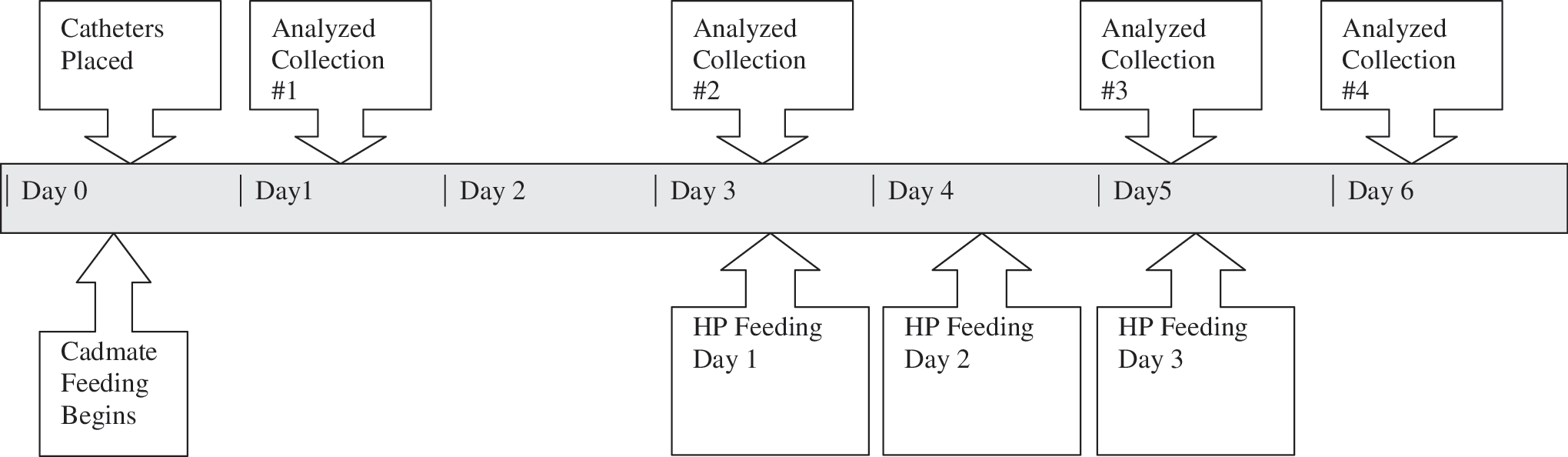
Study timeline. HP = hydroxyproline.
For each 24-hour collection, total urine excreta was recorded and subsampled. Urine pH was recorded. Fresh urine samples were collected and stored at 4°C until analysis. Oxalate and glycolate analysis were determined by ion chromatography as described by Knight et al. 5
Results
All the 12 sows completed the 24-hour urine collection on day 1. (Table 1) Because of the infection and/or catheter-related problems, two sows were excluded on day 3, one sow on day 5, and three sows on day 6. The addition of HP to the diet increased urinary oxalate excretion. Before feeding HP (day 1), the mean oxalate excretion was 3.1 meq/d and 1.98 meq/d in the CD and AD groups, respectively. On day 3, the mean oxalate excretion rose to 5.92 meq/d (CD) and 3.44 meq/d (AD). On day 5, the mean oxalate excretion was 7.385 meq/d (CD) and 2.66 meq/d mg/L (AD). On day 6, the mean oxalate excretion had declined to 3.58 meq/d (CD) and 1.68 meq/d mg/L (AD) (Table 1). Overall, there was a 145% (CD) and 74% (AD) increase in urinary oxalate. The increase peaked on day 5 in the CD group and day 3 in the AD group and gradually declined in each group by day 6. Student's paired t-test was performed for both the groups and confirmed that oxalate on day 3 was significantly different than baseline (p = 0.04).
Urinary glycolate also increased as a result of adding HP to the diet. Mean baseline glycolate (day 1) was 0.099 meq/d (CD) and 0.099 meq/d (AD). On day 3, the mean glycolate excretion rose to 9.05 meq/d mg/L (CD) and 8.54 meq/d (AD). On day 5, the mean glycolate excretion declined to 8.07 meq/d (CD) and 5.24 meq/d (AD). On day 6, the mean glycolate excretion declined to 1.44 meq/d (CD) and 1.13 meq/d (AD). Overall, there was a 9000% (CD) and 8500% (AD) increase in urinary glycolate between days 1 and 3. The increase peaked on day 3 and then declined, although it remained more than 10 × greater than baseline at day 6. Student's paired t-test confirmed that glycolate levels on days 3, 5, and 6 were significantly different than baseline (p < 0.001, p = 0.001, and p = 0.002, respectively). As a result of the CAD-MATE feeding, the mean urinary pH over all urine collections was 6.77 (CD) and 5.84 (AD) (Figs. 2 –5).

Urinary oxalate excretion on each day of study. CTL = control.
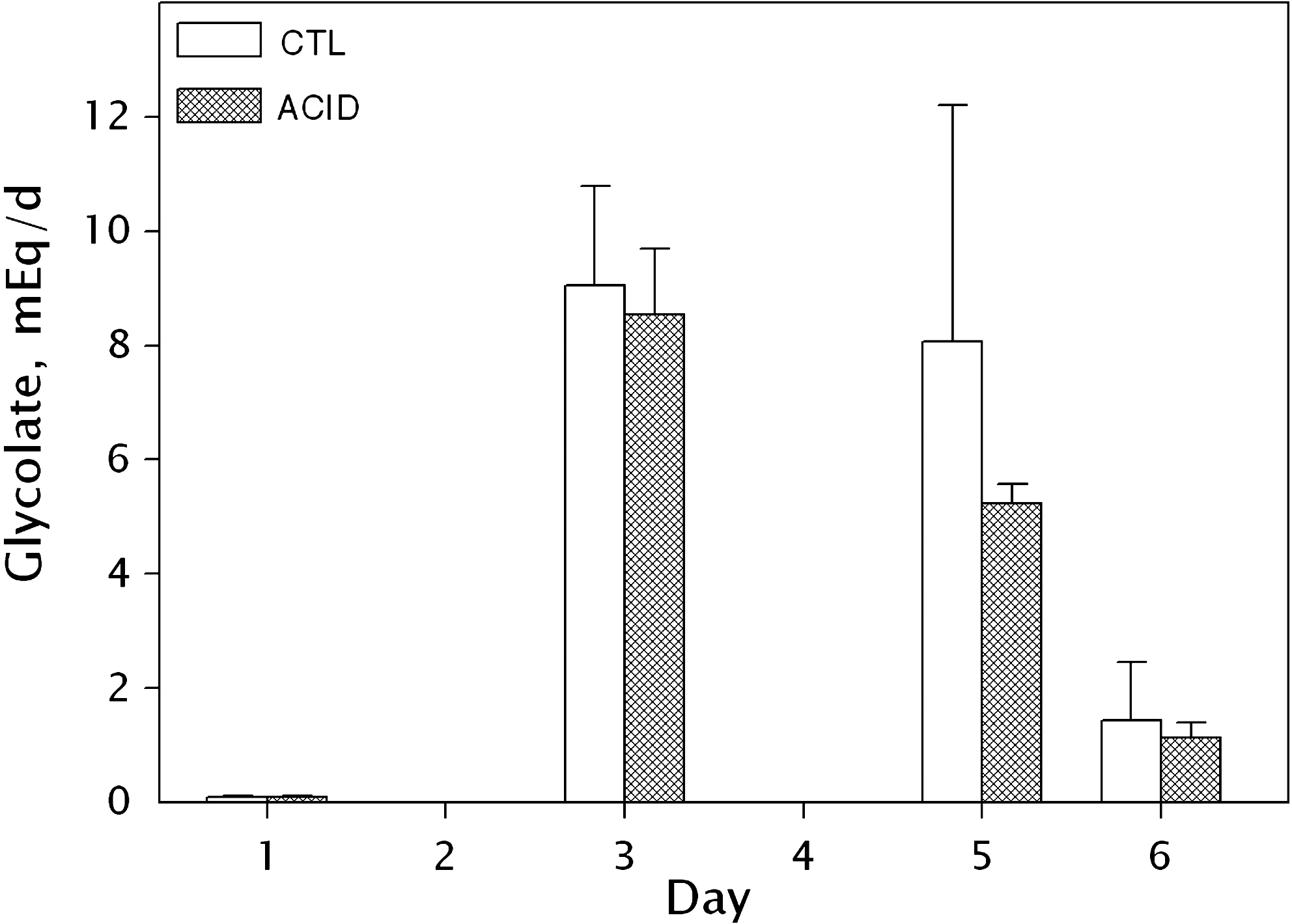
Urinary glycolate concentration on each day of study.
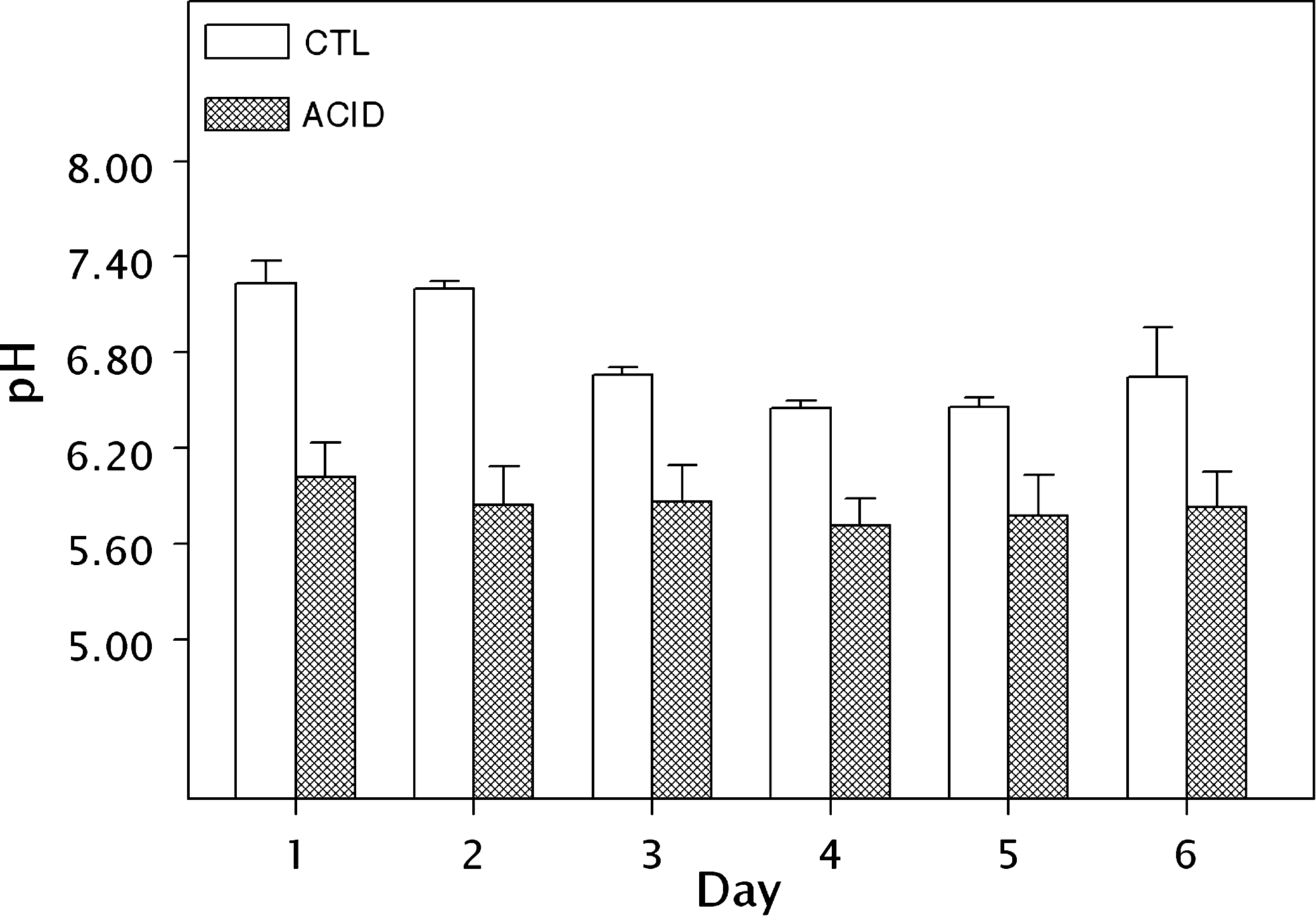
Urinary pH on each day of study.

Method of urine collection. (
Discussion
Our study confirms that hyperoxaluria and hyperglycoluria can be reliably induced by HP in an adult porcine (sow) model. Addition of 10% HP to the diet of gestating sows resulted in a rapid increase in oxalate and glycolate on the first day of feeding. Sows fed a western-type acidified diet had comparable oxalate and glycolate levels at baseline as controls, and they experienced a comparable rise in these levels with HP feeding. These findings are meaningful because they provide a reproducible model for creating hyperoxaluria in the sow using either a traditional diet or a diet that more closely resembles the human western diet.
One unexpected finding was the early peak in the urinary oxalate and glycolate levels on the first day of feeding with subsequent decline despite continued feeding. Mandel et al 2 used 10% HP in young pigs and noted that urinary oxalate levels peaked at around day 6 of feeding and did not decline until HP feeding was stopped. One explanation for this discrepancy may be the differences that exist in the metabolic capacity between the adult and young growing pigs. The adult sows, fed to meet maintenance energy requirements, may adjust faster to the HP loads and have greater metabolic capacity than the young growing pigs. Alternatively, metabolic adaptations in enzymatic capacity may have allowed a decrease in excreted oxalate on the later days of the study. A better understanding of HP metabolism will ultimately help explain these findings.
We must also consider that the decline in the urine oxalate concentrations may reflect an acquired aversion to the HP diets. Based on the observations, sows consumed less of their daily meals on subsequent days in which HP was added to the basal diets and appeared to waste more feed. No attempt was made to record the amount of food wasted, a factor that could have contributed to the declining oxalate levels over time. The amount of HP given per meal may have bearing on oxylate and glycolate levels, but it does not appear to be a strict dose–response relationship. Mandel et al 2 did not observe an increase in urinary oxalate when HP was increased to 15% and 20% of the meal. In a study in which rats were fed HP in varying proportions, no further increases in oxalate excretion were observed beyond 1% HP concentration. 7 Admittedly, feeding 10% HP in this study may have provided more than needed to create the same levels of hyperoxaluria and hyperglycoluria. Future studies will be needed to determine the threshold amount needed.
Consumption of HP -rich animal protein causes an increase in urinary oxalate. 5 This increase is the result of an endogenous pathway and multiple enzymatic reactions. Oxalate metabolism occurs largely in the liver, and to a smaller degree in the erythrocytes. 12 In the liver, glyoxalate is the immediate precursor to oxalate, and glyoxalate concentrations are relatively low as glyoxalate is rapidly converted to glycine by alanine-glyoxalate aminotransferase. Glyoxalate can alternatively be converted to glycolate in the cytosol by D-glycerate dehydrogenase. 12 Thus, the fate of glyoxalate that results from HP ingestion is reduction to glycolate, conversion to glycine, or production of oxalate via lactate dehydrogenase in the cytoplasm of liver cells. 12 Our data demonstrate a concurrent rise in both oxalate and glycolate, indicating that the pathways leading to each has been activated. Future research must investigate the relative importance and contribution of the pathways leading to glyoxalate formation, because this information can aid our understanding of hyperoxaluria.
Because our goal was to create a model of hyperoxaluria, we opted to only monitor the urinary oxalate levels. Collections and analysis to determine serum HP, oxalate, and glycolate concentrations would provide additional insights. These values may have allowed better understanding of the mechanisms driving the urinary changes. In addition, measures of other major urinary cations and pH would offer a better understanding of systemic changes in the renal adaptation to HP interventions. Also, as stated above, lower concentrations of HP may have allowed us to determine the minimal amount needed to induce a detectable hyperoxaluria. Pathologic examination of the sow's kidneys may also have been valuable, as previous studies have shown that HP ingestion may cause the formation of calcium oxalate crystal deposits. Although the short time course of our study would likely have not allowed for such formation, it is possible that precursor to lesions could be detected.
In summary, we have demonstrated the feasibility of using the adult sow as a model for hyperoxaluria. Ingestion of HP results in an almost immediate state of hyperoxaluria which is statistically significant when compared with the controls. Hyperoxaluria also results in sows fed a diet that closely resembles the acid load of a human western diet. Such a modification is easy to make and potentially very useful when extrapolating data to humans. Moreover, the larger animal model will have benefits as a model for oxalate stone disease.
Conclusion
Medical prevention and the management of calcium oxalate nephrolithiasis will require a comprehensive understanding of oxalate metabolism in humans. A model for human hyperoxaluria can be reliably created in the adult sow. Such a model will be useful to further our understanding of oxalate metabolism and ultimately aid in the prevention of calcium oxalate calculi.
Footnotes
Disclosure Statement
Daniel M. Kaplon, Kristina Penniston, Camille Darriet, and Thomas Crenshaw—none. Stephen Y. Nakada: Cook Urological Inc., Consultant/Advisor.