
Abstract
Purpose:
The purpose of our study was to determine if a hydroxyproline (HP) or gelatin diet could induce long-term hyperoxaluria in the porcine model.
Materials and Methods:
A total of 18 gravid crossbred sows (Large White × Landrace) were randomly allotted into three treatment groups: 5% HP, 10% HP, and gelatin diet. All sows were catheterized 1 day before starting treatment diet. Catheters were left in place for 5 days before being removed. Sows were recatheterized for urine collections on days 11 to 12 and days 21 to 22. Urine was collected for each entire 24-hour period, and urinary oxalate was determined by ion chromatography.
Results:
Urinary oxalate concentrations for all three diets peaked within the first 5 days of the diet. The sows fed the 5% HP, 10% HP, and gelatin diets had an early peak in urinary oxalate concentration (mg/L) at day 2 (158% increase), day 5 (316% increase), and day 5 (830% increase), respectively. The day 21 to 22 time points in all three diets demonstrated markedly increased urinary oxalate concentrations in comparison with baseline, with some concentrations higher than the early time point peaks (day 22: 5% HP=1906% increase, P=0.12; 10% HP=640% increase, P=0.02; gelatin=501% increase, P=0.01).
Conclusion:
Although both the 10% HP and gelatin diets induce significant short- and long-term hyperoxaluria in the porcine model, the gelatin diet is more cost-effective. The ability to induce long-term hyperoxaluria has important implications in establishing a porcine model for oxalate urolithiasis.
Introduction
Oxalate metabolism is critical to understanding calcium-based urolithiasis. Alhough urinary oxalate is believed to be derived equally from dietary intake and endogenous synthesis, the pathways leading to oxalate synthesis have not been fully elucidated. 4 Studies on type I and II primary hyperoxaluria have shown that high oxalate levels can be from deficiencies in the glycolate metabolizing enzymes alanine-glyoxylate aminotransferase and glycolate reductase. 5 Although glycolate is a precursor to endogenous oxalate, the precursors of glycolate and glyoxylate have not been well elucidated. 6 Hydroxyproline (HP) is a derivative of the amino acid proline and a component of collagen and thus found in animal proteins. 6 HP is metabolized to both pyruvate and glycolate in hepatic mitochondria and in the renal proximal tubule. 6,7 In humans, collagen is metabolized at a rate of 2 to 3 g daily leading to the metabolism of 240 to 420 mg of HP and the formation of 140 to 240 mg of glycolate. 8
The rat has been the primary animal model used to induce hyperoxaluria, calcium oxalate crystalluria, and urolithiasis (Table 1). Although the rat model has allowed us to test various techniques of inducing urolithiasis, including various dietary manipulations, there are significant dietary, anatomic, metabolic, and physiologic differences between rats and humans. Mandel and associates 9 demonstrated that young growing pigs fed HP became hyperoxaluric; crystalluria developed as did calcium oxalate plaques on the tips of renal papillae.
Ref=reference number; CaOx=calcium oxalate; HP=hydroxyproline.
Sustained hyperoxaluria is important in an animal model to induce histologic renal crystal deposition. 9 Our previous study demonstrated that a 10% HP diet could lead to the short-term induction of hyperoxaluria with a decrease in urinary oxalate by day 5. 10 Because of uncertainty regarding decreased feed consumption with a 10% HP diet, we investigated the hyperoxaluric effect of both a 5% and 10% HP diet in our current study. 10 Knight and colleagues 6 demonstrated that a gelatin diet fed to healthy human subjects resulted in elevated levels of urinary oxalate and glycolate. 6 HP is costly ($185/kg), so we chose to pursue the hyperoxaluric effect of a gelatin diet in the porcine model because it would be more cost-effective. We hypothesize that both HP and gelatin diets induce short and long-term hyperoxaluria in the porcine model.
Materials and Methods
The study included a total of 18 gravid crossbred sows (Large White × Landrace). Sows were housed at the University of Wisconsin Swine Research and Teaching Center. The experimental protocol was approved by the University of Wisconsin Research Animal Resource Committee. The sows were randomly allotted into one of three treatment groups: 5% HP, 10% HP, and gelatin diet.
Three days before starting the treatment diet, each sow was fed 2 kg of a basal diet. 10,11 Starting on day 1, each sow was fed 2 kg of the treatment diet and allowed continuous access to water. The treatment diet consisted of the basal diet evenly mixed with 100 g of L-HP (5% HP diet) (Wilshire Technologies, Princeton NJ), 200 g L-HP (10% HP diet), or 1000 g gelatin (Milligan and Higgins, Johnston, NY) (Fig. 1.) Amino acid analysis of the gelatin revealed a 12% HP and a 20% glycine content by weight. All sows were catheterized 1 day before starting the treatment diet. Each sow was catheterized with an 18F Foley catheter and the balloon inflated with 30 mL sterile water. Tubing was attached to the catheter and placed into the cap of a 20-L collection pail (Fig. 2). Each collection pail was weighed before being placed in ice-filled plastic tubs. 10,12 Sows were closely monitored for signs and symptoms of cystitis, and catheters were removed in cases of infection.

Gelatin (left) and L-hydroxyproline (right) crystals that were used to create the sow treatment diets.

Sows with Foley catheters in place attached to tubing leading into collection pails.
Catheters were left in place for 5 days before being removed. Sows were then recatheterized for urine collection on days 11 to 12 and days 20 to 21. Urine was collected for each entire 24-hour period to control for differences in the diurnal and postprandial variations in the renal handling of oxalate. An overview of the study timeline is described in Figure 3. For each 24-hour collection, total urine weight was recorded and subsampled. Fresh urine samples were collected and stored at 4°C until analysis. Urinary oxalate concentration was determined by ion chromatography using a Dionex DX-500 Ion Chromatograph (Dionex 4×250 mm Ionpac AS14 analytical column; 4×50 mm Ionpac AG14 guard column; eluant:100% 1.0 mM NaHCO3/3.5 mM Na2CO3; pressure: 800–1200 psi; flow rate: 2 mL/min). 6 To further confirm that the treatment diets had the expected impact on urinary oxalate, mass spectrometry (Dionex Corp., Sunnyvale, CA) was used to confirm increased urinary glycolate in a subset of the samples.

Study timeline.
All results are presented as mean values±standard deviation. Percent change was calculated for each of the time points compared with the baseline urinary oxalate concentration from day 0. On days in which there were missed urine collections (because of urinary tract infection and removal of the catheter), the percentage change from baseline was calculated by comparing the mean urinary oxalate concentration for the animals present at the time point compared with the mean baseline concentration of the same animals at day 0. The statistical significance of the results was assessed via paired and unpaired t tests for comparisons between each of the three diets and for comparisons made for time points between animals in the same diet group, respectively. Statistical significance was set for P values≤0.05.
Results
Of the 18 sows that began the study, 5 sows were excluded because of infection and/or catheter-related problems leaving 3 sows on the 5% HP diet, 6 sows on the 10% HP diet, and 4 sows on the gelatin diet. The catheter had to be removed, and thus urine collections were missed for urinary tract infections or catheter-related problems for some of the time points throughout the study (Table 2, a and b). All sows in the study, however, had a minimum of urine collections on days 0 to 2 and one set of urine collections at the later time points (days 11–12 and/or days 21–22) to allow us to evaluate for short- and long-term hyperoxaluria.
HP=hydroxyproline; SD=standard deviation; % inc=percentage increase from baseline diet (day 0); **=P<0.05 when compared with baseline urinary oxalate for the diet.
Two missed urine collections because of urinary tract infection at time point.
One missed urine collection because of urinary tract infection at time point.
On days with missing urine collections, % inc was calculated by comparing urinary oxalate concentrations of the same animals at the time point and from day 0.
Urinary oxalate concentrations for all three diets peaked within the first 5 days of the diet. The sows fed the 5% HP, 10% HP, and gelatin diets had an early peak in urinary oxalate concentration (mg/L) at day 2 (158% increase), day 5 (316% increase), and day 5 (830% increase), respectively. Urinary oxalate concentrations at the day 11 to 12 time points demonstrated an elevation in oxalate concentration (mg/L) compared with baseline (day 12: 5% HP=190%; 10% HP=315%; gelatin=64%) but had lower oxalate concentrations (mg/L) when compared with the earlier time point peak (Fig. 4). The day 21 to 22 time points in all three diets demonstrated markedly increased urinary oxalate concentrations in comparison with baseline with some concentrations even higher than the early time point peaks (day 21: 5% HP=1906%; 10% HP=640%; gelatin=501%).
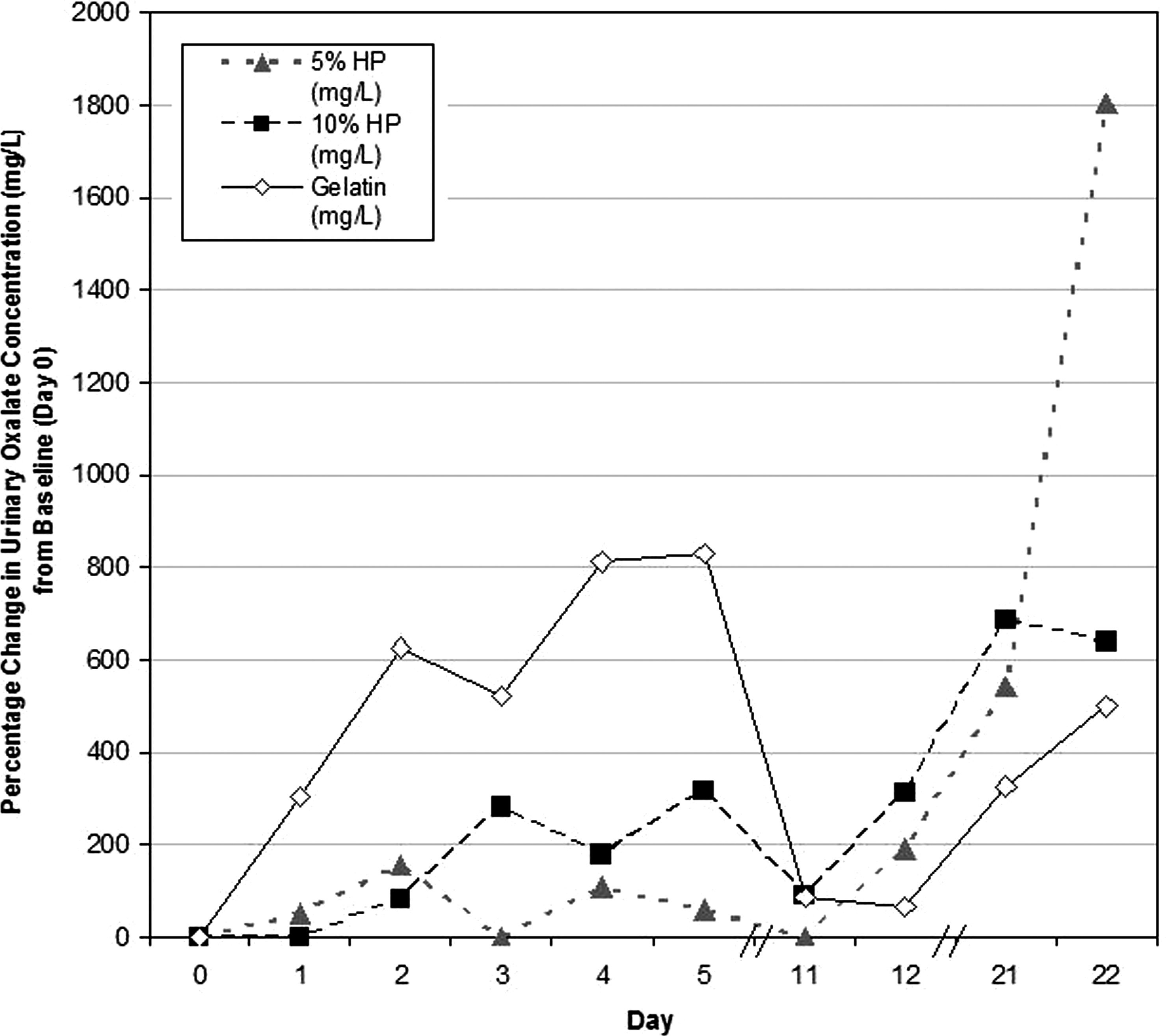
Percentage change in urinary oxalate concentration (mg/L) on each day of the study. HP=hydroxyproline.
In the 10% HP diet, day 2 (P=0.03), day 21 (P=0.04), and day 22 (P=0.02), all demonstrated a significant increase in urinary oxalate concentration (mg/L) compared with baseline. In the gelatin diet, day 3 (P=0.03) and day 22 (P=0.01) demonstrated a statistically significant increase in urinary oxalate concentration (mg/L) compared with baseline. There were no significant differences in urinary oxalate concentration (mg/L) when comparing the time points between each of the three diets.
Discussion
Our study confirms that short- and long-term hyperoxaluria can be reliably induced by HP and gelatin diets in an adult porcine (sow) model. Mandel and coworkers 9 fed 10% HP to young pigs and noted that urinary oxalate levels peaked around day 6 of feeding and did not decline until HP feedings were stopped. Our earlier work with gestating sows fed a 10% HP diet showed a rapid rise in urinary oxalate by day 3 with a subsequent decline to baseline urinary oxalate concentration by day 5. 10 Given the short time course of our earlier study, it was difficult to ascertain if long-term hyperoxaluria was attainable. It was hypothesized that differences in induction of hyperoxaluria between the studies by Mandel and associates 9 and Kaplon and colleagues 10 may have been because of differences in the metabolic capacity between adult and young growing pigs.
Both the 10% HP and gelatin diets induced significant hyperoxaluria in comparison with the 5% HP diet. Our early study time points (days 0–5) demonstrated a peak in urine oxalate concentrations similar to the peak observed by Kaplon and coworkers. 10 Urinary oxalate concentrations decreased at the day 11 to 12 time points but were still higher than baseline urine oxalate concentrations on day 0. Despite a decrease in diet consumption by day 5 for all three groups (approximately 1.5 kg of diet consumed), feed consumption remained constant from day 5 to the end of the study. The cause for the decrease in hyperoxaluria at the day 11 to 12 time points may be partially explained by the decrease in feed consumption by the sows or because of metabolic adaptations in enzymatic capacity that may have led to decreased excretion of oxalate. The day 21 to 22 time points, however, did lead to significant hyperoxaluria compared with baseline for the 10% HP and gelatin groups.
Despite the similar hyperoxaluric effect between the 10% HP and gelatin diets, there is a significant difference in the cost of the two diets ($7.32/kg for 150 Bloom Gelatin, Milligan & Higgins vs $185.00/kg for hydroxy-L-proline, Wilshire Technologies). Thus, the gelatin diet is more cost-effective for the induction of hyperoxaluria in the sow.
Knight and colleagues 6 demonstrated that the consumption of HP in the form of gelatin could lead to an increase in urinary oxalate concentration in humans. Oxalate is derived from endogenous production, in the liver and in erythrocytes, as well as exogenous sources from dietary intake as well as via ascorbate metabolism. 13 The liver is the primary source of endogenous oxalate with glyoxalate as its primary immediate precursor. Glyoxylate concentration in the liver is generally suppressed because of the metabolism of glyoxalate to glycine, which is further catalyzed by alanine-glyoxylate aminotransferase. Glyoxylate can also be converted to glycolate in the liver cytosol by D-glycerate dehydrogenase. 6 Gelatin has both a high HP (12%) and glycine (20%) content; thus, besides the conversion of HP to oxalate, dietary glycine may also play a role in increasing the concentration of glyoxylate and thereby oxalate.
Further research to assess the contribution of other precursors to glyoxylate and thus oxalate formation is warranted to aid in our understanding of hyperoxaluria. Studies exploring oxalate and glyoxylate metabolism in the liver and the blood will be needed to help us further characterize their roles in hyperoxaluria. 7 In addition to urinary oxalate, measurement of other major ions and urinary pH will allow us to better understand the systemic changes that occur in the renal adaptation to HP and gelatin diets. Because both the 10% HP and gelatin diets have demonstrated the induction of significant long-term hyperoxaluria, we are currently pursuing a necropsy study to determine if the resultant elevated urinary oxalate concentrations are adequate to induce crystal deposition in porcine kidneys. From our current results, we believe that this large animal model shows promise as a more versatile model for oxalate urolithiasis.
Conclusion
Although both the 10% HP and gelatin diets induce significant short- and long-term hyperoxaluria in the porcine model, the gelatin diet is more cost-effective. The ability to induce long-term hyperoxaluria has important implications in establishing a porcine model for oxalate urolithiasis.
Footnotes
Disclosure Statement
Dr. Nakada is a consultant/advisor for Cook Urological Inc. For the remaining authors, no competing financial interests exist.