
Abstract
Background and Purpose:
We previously reported hyperoxaluria and calcium oxalate calculi in adult pigs (sows) fed hydroxyproline (HP). The purpose of this study was to grossly and histopathologically characterize intrarenal effects in this model.
Methods:
In the swine facility at our campus, we maintained 21 gestating sows, of which 15 received daily treatment (5% HP mixed with dry feed) and 6 received no treatment (controls). Nine were sacrificed at 21 d (three control, six HP). All kidneys were extracted and examined grossly and for radiographic evidence of stones (GE CT scanner, 80kV, 400MA, 1 sec rotation, 0.625 mm slices). Papillary and cortical samples were processed for histologic analysis.
Results:
Kidneys from treated sows showed significant calculi distributed within the renal papilla on CT, appeared mottled in the renal cortex and papillary areas, and had less distinct corticomedullary borders. Tiny crystals and mucinous debris lined the papillary tips, calices, and pelvis in kidneys from four of six treated sows, and multiple stones were noted at the papillary tips. Hematoxylin and eosin stain revealed crystals in collecting tubules and papillary tips in treated kidneys and none in controls. Yasue staining confirmed crystals in proximal periglomerular tubules of treated but not control animals. Tubular dilation and inflammatory/fibrotic changes were identified in kidneys from treated animals; none of these changes were evident in control kidneys.
Conclusions:
We report renal damage as a result of dietary-induced hyperoxaluria in adult sows. Specifically, we found crystalluria in proximal periglomerular tubules and collecting ducts, with tubular damage at all segments.
Introduction
N
Oxalate excreted in the urine is derived in approximately equal proportions from diet and endogenous synthesis. 7 Hydroxyproline (HP) is a precursor for endogenous oxalate synthesis, and the metabolism of HP primarily occurs in the mitochondria of hepatocytes and renal proximal tubule cells. 8 Collagen degradation is the major source of HP and has a daily turnover in humans of 2 to 3 g/day. 9 We have previously demonstrated sustained, predictable, and reproducible hyperoxaluria through dietary manipulation in our ongoing development of a porcine model. 10,11 In this study, we sought to characterize crystal formation with HP feeding and to describe associated renal changes in gestating sows. As a secondary objective, we sought to verify that nongestating sows responded similarly to HP feeding with respect to stone formation.
We hypothesized that (1) 21 days is sufficient for studying HP-induced stone formation and associated urinary and renal changes in this model, and (2) kidneys from sows forming stones would have histologic evidence of renal damage.
Methods
The study was approved by the University of Wisconsin (UW)-Madison Research Animal Resource Committee. Animals were housed at the UW Swine Research and Teaching Center, an Association for Assessment and Accreditation of Laboratory Animal Care-accredited swine facility with full-time staff support.
Experimental design
The overall experiment had two parts: study #1 was designed to study the renal effects of the dietary intervention in gestating sows and was carried through to 42 days; study #2 was a secondary experiment to verify our model in nongestating sows.
Study #1 – dietary intervention, gestating sows
The treatment diet (TD) consisted of 5% HP (100 g in 2 kg) evenly mixed with the standard UW gestation diet comprised of corn, soybean meal, mineral and vitamin supplements, while the control diet (CD) consisted of 5% (100 g in 2 kg) starch mixed with the standard UW diet. The gestating sows were initially acclimated to a 2 kg basal diet for 2 weeks before day 1 of the experiment. At day 1, 21 sows were randomly allotted to the TD (n=15) and CD (n=6) groups. The TD group was further divided into 21 days (TD-21) and 42 days (TD-42) of dietary intervention, with six and nine sows, respectively. Animals in each group were sacrificed at the end of the intervention using standard U.S. Department of Agriculture methods (USDA Food Safety and Inspection Service Directive 6900.2, revision 2; 8/15/2011), and their kidneys were extracted for imaging and evaluation.
Study #2 – verification of model in nongestating sows
We assessed stone formation in four nongestating sows of similar age and weight (two CD and two TD sows) as above. Treatment sows were maintained on the TD for 21 days, and all animals were sacrificed at 21 days, and kidneys were extracted as described above.
Tissue processing
The extracted kidneys were immediately flushed with heparinized saline, maintained in ice, and taken to the CT suite. The ex-vivo kidneys were scanned on a GE Discovery CT750 HD CT scanner using a protocol optimized for stone detection (80kV, 400MA, 1 sec rotation, 0.625 mm slices). Images from the unlabeled kidneys were then examined for radiographic evidence of stones using standard bone windows.
After scanning, the kidneys were bivalved and examined. Gross appearance was recorded and digital photographs were taken; loose stone material was documented, collected in individual jars, and sent for analysis. Whole kidneys were fixed in 10% formalin for 48 hours, and subsequently, four transverse sections of each of the following were taken: Upper, midpolar, and lower pole papilla and renal cortex. Paraffin embedded blocks of each representative region were prepared, sectioned at 5 microns, and processed for the following stains: Hematoxylin and eosin (H&E), Yasue metal substitution, and periodic acid Schiff. The stained sections were then evaluated microscopically under incandescent and polarized light with the assistance of an experienced research pathologist; digital photographs were obtained.
Results
Study #1 – Dietary intervention, gestating sows
The TD kidneys had extensive hyperattenuating material distributed throughout the medullary tips and papilla and within the calices (Fig. 1A); there was no evidence of cortical calcifications. No calcifications were evident in the CD kidneys. The calculated stone size ranged from submillimeter crystals up to 2 mm. The Hounsfield unit density in the larger stones was in the range of 400 to 500.

CT scans showing calcifications in the renal collecting system of a pair of treatment diet-21 kidneys.
Kidneys from CD animals were normal in overall gross appearance (Fig. 2A). In the TD kidneys, the renal cortex and papillary areas appeared mottled (Fig 2B); corticomedullary borders were less distinct compared with controls. Tiny crystals (yellow-white in appearance) and mucinous debris lined the papillary tips and calices in 7 of 12 kidneys from treated sows (Figs. 2B, 2C). Transverse sections of the papilla showed crystals aligned in the direction of the medullary collecting ducts (Fig. 3A) and numerous stones at the papillary tips (Fig. 3B). Yellow calcified deposits were seen to project from the tips of the papilla, plugging the openings of the distal collecting ducts (Fig. 3C). There was no visible evidence of Randall plaques in any of the papilla.
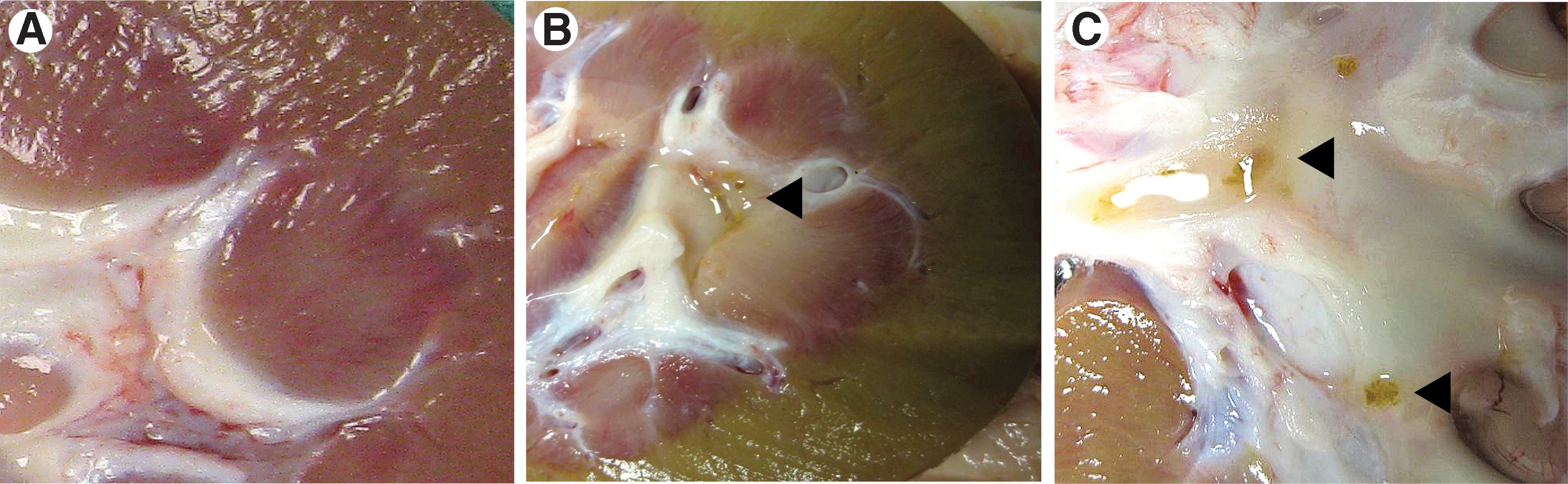
Bivalved kidneys at 21 days.
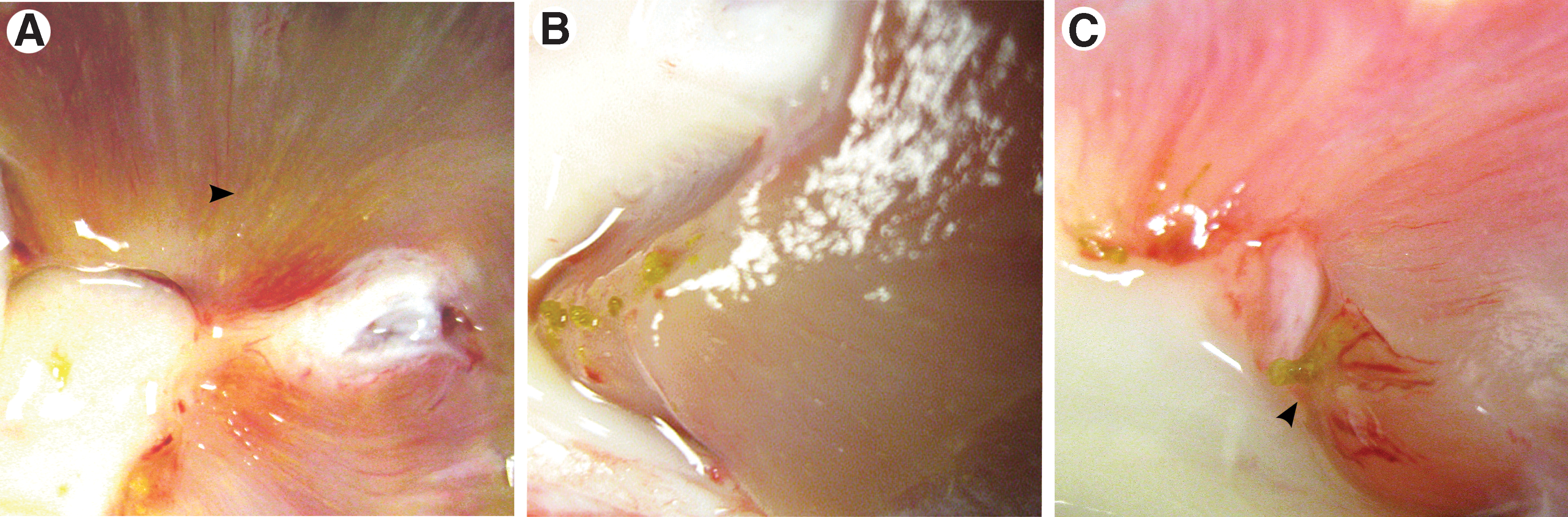
Gross dissection of treatment diet kidneys.
Study #2 – Verification of model in nongestating sows
While the two TD sows were initially fed 10% HP diets, their intake declined after 5 days. Therefore, the HP concentration was reduced to 5%, and their tolerance and regular feed consumption was reestablished. After 21 days, all animals were sacrificed and their kidneys were extracted. CT scanning of the kidneys demonstrated calculi near the papillary tips of all four kidneys from the TD sows, which were up to 1.6 mm in maximal diameter; no cortical calcifications were identified. Kidneys from the CD sows had no identifiable stones.
Stone analysis
The retrieved stone material ranged in size from <1 mm crystals to 2.1 mm with a pale white-yellow appearance. Five separate specimens from five of the treated sows were sent for analysis by micro CT (James Williams, PhD, Indiana University). Each appeared as a cluster or aggregate of calcium oxalate particles and crystals. Micro-CT examination of the largest of these (1.9 mm in length) revealed polyhedral crystals, consistent with calcium oxalate dihydrate, and a smaller area of round lumps, which had an x-ray attenuation consistent with calcium oxalate and which were presumed to be calcium oxalate monohydrate (Fig. 4).
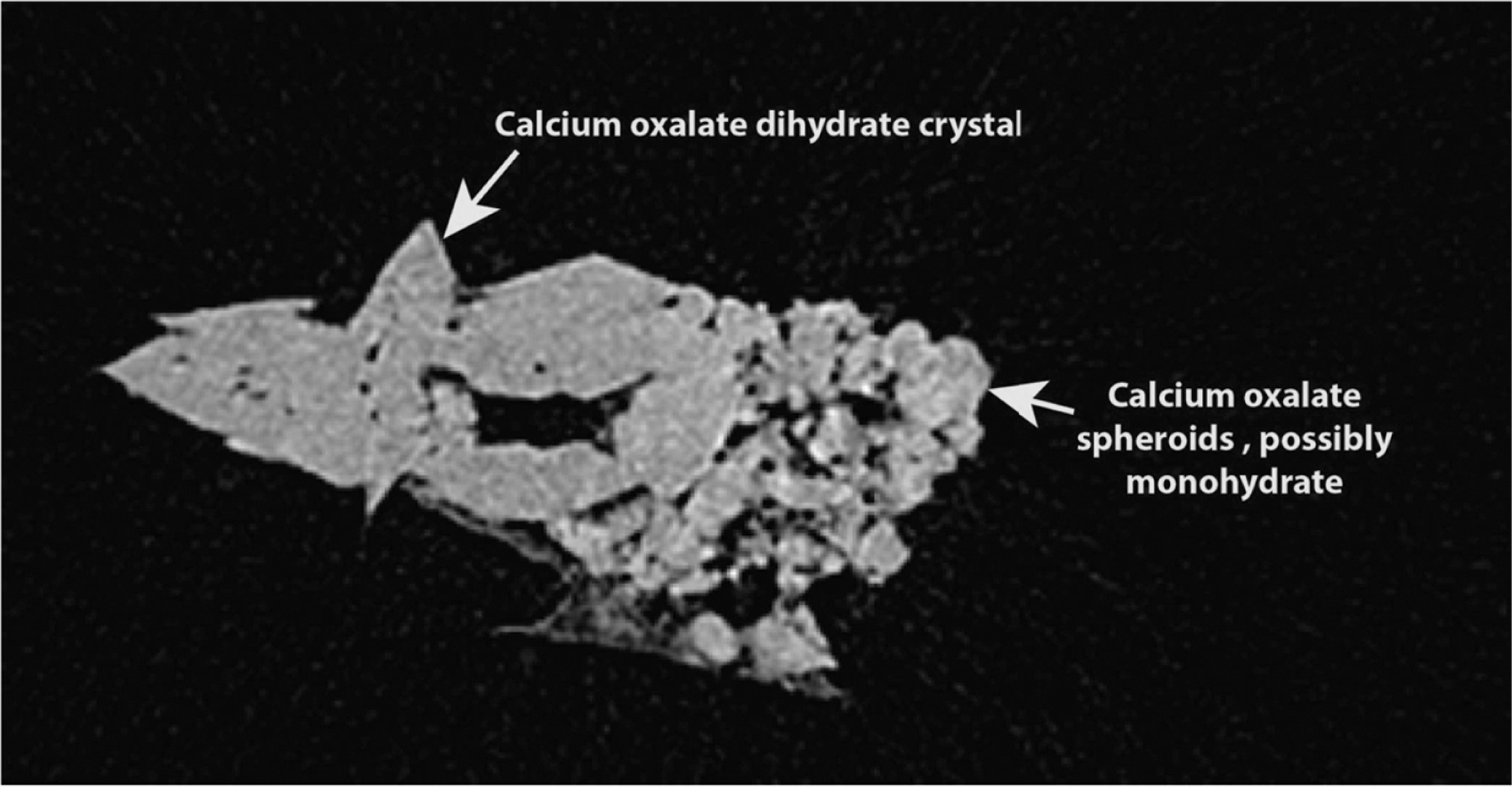
Micro-CT demonstration of an extracted stone, confirming a combination of calcium oxalate dihydrate and monohydrate crystals.
Histology
H&E staining revealed crystals within the collecting tubules and at papillary tips of two-thirds (67%) of TD kidneys but in none of the controls (Figs. 5A, 5B). Yasue staining confirmed that these crystals contained calcium (Figs. 5C, 5D). Tubular epithelial damage indicated by epithelial attenuation and ulceration was observed with H&E in some areas with heavy intratubular crystals (Figs. 6B, 6C). Notably, the sporadic presence of crystals in cortical tubules and interstitium was observed (Fig. 6B). In some of the TD kidneys, many crystal aggregates were not clearly associated with tubules and appeared embedded in the interstitium of the papilla (and cortex), although loss of discernible tubular structures with apparent interstitial localization of crystals is possible. The interstitially localized crystals were often surrounded by multinucleate giant cells (Fig. 6C), and occasionally the same was observed with intratubular crystals.

Papillary tips at 21 days, 4× magnification. Hematoxylin and eosin stain:

Cortical section, 21 days, H&E 20×.
Some crystals appeared to expand between the renal tubular epithelium and the underlying interstitial connective tissue. The interstitium surrounding some regions of crystal accumulation in the cortex was infiltrated by small to moderate numbers of mononuclear leukocytes; inflammation surrounding crystals in the papilla was more commonly limited to multinucleate giant cells, although sometimes lymphoid cells and polymorphonuclear leukocytes were present. The cortices of several TD kidneys exhibited narrow radiating regions of mild widening of the interstitium with extracellular matrix interpreted as fibrotic changes (Fig. 6D). These radiating zones were both associated and unassociated with crystals in or near the lesions and often had mild to moderate mononuclear inflammatory infiltrates.
In addition to crystals, several of the animals had aggregates of lamellar eosinophilic and basophilic material in the papilla interpreted as mineral. Striking polymorphonuclear and mononuclear inflammation of the papilla was noted in one animal with crystal accumulation at the papillary tip with evidence suggestive of ascending pyelonephritis. CD kidneys exhibited minimal changes generally limited to mild occasional lymphocytic interstitial infiltrates, minimal interstitial widening with fibrous tissue, rare individual inflamed or regenerative renal tubules, and occasional nonpolarizing aggregate accumulation (interpreted as mineral) in the papilla (note that only six CD kidneys were examined).
Discussion
We have developed a porcine model in which sustained hyperoxaluria and renal crystal formation is achieved with dietary intervention. 10,11 In the present study, we reproduced calcium oxalate crystal formation in our model within 21 days of HP feeding and identified renal derangements associated with hyperoxaluria. Specifically, we demonstrated renal stone formation in adult swine, crystals in proximal tubules adjacent to glomeruli and in collecting ducts, and signs of inflammatory tubular damage at all segments. This damage was characterized by giant multinucleated cells, degeneration of tubular epithelium, expansion of the interstitium with matrix deposition, and rarely the extrusion of crystal into the interstitium of TD but not CD kidneys—such changes indicate an acute inflammatory response, presumably to the stress of hyperoxaluria and crystalluria, which appeared to culminate in an early fibrotic process. Although the most apparent crystal formation was seen within expected regions of the kidney (ie, renal papilla and distal tubules), we also noted proximal tubular damage at the level of the renal cortex.
Observations similar to ours have been reported in rodent studies, 12,13 whereby a short-term induction of hyperoxaluria and crystalluria led to tubular epithelial destruction and widened interstitial spaces 14 and migration of calcium oxalate crystals from the tubular lumen into the interstitium. 15 The striking similarities of our results with those using established rodent models of urolithiasis strengthen the validity of our model.
Although not a major amino acid in the human diet, HP is consumed in small amounts in meats and gelatin containing foods. 16 Ours is not the first study to use HP as a nutrition intervention. Mandel and colleagues 17 demonstrated short-term hyperoxaluria, peaking at day 6, in young swine fed 10% HP. 17 Whereas Mandel and associates 16 used young growing pigs, which have different metabolic capacity than adult pigs, we used adult sows because they may more closely resemble adult humans. In developing our model, we devised a reliable method for sustaining sows' consumption of HP-enriched feed, for obtaining 24-hour urine collections, and for measuring urinary oxalate excretion. In Kaplon and coworkers, 11 we reported hyperoxaluria in adult sows fed HP for 5 days in both their standard feed and in feed that was acidified to mimic the acid load of the typical Western diet. Thereafter, Patel and associates 10 demonstrated that both short (5 days) and long-term (21 days) hyperoxaluria (similar in scale to human values) was reliably induced with HP and also with gelatin (the latter of which was used by Knight and colleagues 16 in a similar human study). A single kidney from a single treated animal revealed a small calcium oxalate stone. 10
Using this paradigm, we now report stone formation in both gestating and nongestating sows. We observed calculus deposits (“plugs”) at the papillary tips and occasional submucosal crystal deposits at the papilla, similar to findings from human intestinal bypass as reported by Evan and colleagues. 18 Our results also showed fibrotic changes in the cortical tubules adjacent to the glomeruli of treated sows, suggesting that renal damage occurs in the initial portion of proximal renal tubules. Despite the inflammatory effects we observed, only one of the TD animals had histologic signs of infection, suggesting that these changes are sequelae of hyperoxaluria and not of infection.
Results from this model may have clinical relevance. Because our model increases endogenous oxalate, it may be relevant for studying primary hyperoxaluria. Calcium oxalate crystal formation in these patients usually occurs initially, however, in the renal parenchyma as nephrocalcinosis and frequently progresses to reduced renal function, leading to extrarenal oxalate deposits. In our study, we observed crystals primarily within the renal tubules and minimal to no evidence of intracellular crystals as would be expected with nephrocalcinosis. Accordingly, although our model is not characterized by an intestinal challenge, our results are similar to those observed in patients with short bowel, 18 whose intestinal calcium absorption is usually very low and whose urinary oxalate excretion is subsequently extremely high. Our model has potential relevance for studying enteric calcium oxalate stone disease.
A limitation of our study is the possibility that direct potential toxicity induced by renal reabsorption and cortical metabolism of HP (as suggested by Knight and coworkers 16 ) might contribute to some of the renal effects observed. This could mean that the changes may not necessarily be attributable to hyperoxaluria and/or to intratubular calcium crystal deposition. Nonetheless, we detected similar tissue architectural changes to those seen in human calcium oxalate stone formers, 18 thereby supporting the theory that these changes are truly from crystalluria. Further studies are needed to assess the direct impact HP might have on renal tissue.
In considering the use of animal models for studying urolithiasis, our study is important for a number of reasons: (1) Irrespective of how calcium oxalate lithiasis is induced, the renal effects of hyperoxaluria, stone formation and growth can be studied in this model; (2) similar urinary and renal changes, along with calculus formation, are evident in both gestating and nongestating sows, confirming that both groups respond similarly; (3) we can reliably induce stone formation with 21 days of dietary manipulation, obviating the need for longer, more expensive interventions; (4) our results (a) concur with currently upheld theories of stone formation derived from human studies, specifically regarding papillary extrusion of crystals through ducts of Bellini and submucosal crystal deposition, 18 and (b) corroborate the histologic tissue changes seen in established rodent models. 14,15,19 The overall findings of this study hold promise for further development and use of our porcine model to study the pathophysiology of calcium oxalate urolithiasis.
Conclusion
We demonstrated crystal formation and described the associated stress-induced histopathology in a porcine model of urolithiasis. This is the initial report of dietary-induced renal calculi in adult sows, with crystalluria in cortical proximal tubules and throughout the collecting ducts and with tubular damage at all segments. Further studies will allow us to elucidate the etiology of renal damage and identify potential preventive mechanisms.
Footnotes
Acknowledgments
The authors thank the University of Wisconsin Swine Research and Teaching Center staff for their exemplary care of the research animals; Ruth Sullivan, VMD, PhD, for her assistance with histology review and photos; Frank N. Ranallo, PhD, for his assistance with CT protocols and image reconstruction; and Lisa Sampson for her technical assistance with CT imaging and tissue handling.
Disclosure Statement
No competing financial interests exist.