
Abstract
Introduction:
The pathogenesis of infectious kidney stones is poorly understood. Bacteria have been implicated in promoting infectious stones via urease production; however, there is mounting evidence indicating the relationship is more complex. The aim of our study was to characterize suspected biotic and abiotic extrinsic factors that may modulate the formation of infectious stones.
Materials and Methods:
A high-throughput experimental model with Griffith's artificial urine was used to test a wide variety of urinary modulators and cytoplasmic enzymes present in crude cell-free extracts (CFEs) from bacterial strains to investigate how they impact struvite and calcium (Ca) phosphate crystal production. Crystal formation was evaluated with spectrophotometry and growth curve analysis. Light microscopy and scanning electron microscopy/X-ray diffraction was used for crystal structure and composition identification.
Results:
The acidic urinary modulators used in this study prevented crystal formation, whereas osteopontin had a significant inhibitory effect. Addition of CFEs from Proteus mirabilis 175A and 177A resulted in Ca phosphate and struvite crystals. Of interest, Klebsiella pneumoniae and Klebsiella oxytoca produced crystals including Ca phosphate and Ca oxalate, respectively. Pseudomonas aeruginosa had no urease production detected and produced Ca phosphate crystals.
Discussion:
Urinary modulators can have a wide variety of effects on infectious stone formation and the role of pH is important but does not guarantee robust crystal formation. Bacterial strains can produce Ca oxalate, Ca phosphate, and struvite stones with and without urease activity.
Conclusion:
Various urinary modulators appear to influence the process and are worthy of further evaluation as a potential therapeutic strategy to prevent infection-related urinary stone formation. Stones formed from urinary tract infections may be a result of multiple encoded metabolic pathways and discovering these would improve our understanding of the stone-bacterial relationship.
Introduction
Nephrolithiasis is a common condition that affects 10%–11% of people during their lifetime. 1,2 In the United States in 2000, the cost of kidney stones to health care was almost $2.1 billion dollars and there are estimates that this could balloon to $4.1 billion dollars by 2030. 3 Not only are kidney stones prevalent, but they can be a recurrent condition. Without any intervention there is 50% chance of developing another stone within 10 years for all those who had stones. 4 However, patients with calcium (Ca) phosphate stones can have a 75% chance of recurrence. 5 Increased health care resources are required for patients with infectious-based stones because of high stone recurrence rates, infections, sepsis, comorbidities, and the higher rate of surgical intervention. There is a major clinical focus on evaluating stone-forming patients and initiating lifestyle modifications or pharmacotherapies to minimize the risk of recurrence. Despite moderate success with these clinical approaches, there is a significant amount that we do not understand regarding the pathogenesis of urinary stone disease, specifically infectious stones. 6
Struvite stones are traditionally thought to form as a result of infection with urease-splitting bacteria, which produce ammonium ions and reduce urinary pH. The ammonium ions combine with phosphate and magnesium to form struvite crystals. However, infection with the lack of high magnesium ammonium phosphate molar product has been shown to lead to Ca phosphate stones. 7,8 In the absence of infection, Ca phosphate can be associated with specific metabolic conditions, namely distal renal tubular acidosis (low urinary citrate) and primary hyperparathyroidism (high urinary Ca). Two hypotheses have been proposed to explain why Ca phosphate stones form in the absence of clinical infection and without any struvite component. The first theory is that a clinically asymptomatic urinary tract infection (UTI) was responsible for stone formation. 6 The second theory is that an infectious process was involved in the initial steps of the stone formation and disappeared secondarily. 6 Carpentier et al in their review of 39 patients with Ca phosphate stones showed that three of the stones analyzed had bacterial imprints without any history of UTI. 7
Lack of pathophysiologic understanding in infectious urinary stones has led to paucity of preventative strategies with a proven track record of clinical benefit. Antimicrobial agents cannot fully penetrate the crystal lattices of the stone to treat all bacteria, so its benefit is limited once a stone has formed. 9 Antimicrobial stewardship is vitally important because there are long-term risks with prolonged antibiotic use including bacterial resistance and secondary infections. 9 Acetohydroxamic acid is a urease inhibitor that has shown success in small studies but has a significant side effect profile that has prevented its widespread use. With better understanding of the mechanisms behind infectious stone formation, more reliable prevention of their occurrence may lead to improvement in patient care and quality of life.
Testing a wide variety of these modulators under consistent experimental conditions may allow further understanding of the processes involved in infectious stone modulation. This knowledge could potentially lead to the development of novel preventative or therapeutic strategies. The aim of this project was to test urinary modulators in infectious kidney stone formation using a validated high-throughput experimental model. Factors potentially promoting or inhibiting the formation of struvite and Ca phosphate stones were studied. Secondary aims included using the model to explore the role of different bacterial strains on stone formation.
Materials and Methods
Experimental model
A validated experimental model was created in our laboratory. Review ethics board approval from Western University was attained for urine for testing biomaterials. In brief, our model was developed by testing various artificial urine media and comparing them with human urine. Griffith's artificial urine 10 was identified as the most reproducible environment and was associated with comparable infectious crystal formation with the human model. The ideal Jack Bean urease (Cat No. 94280-5G; Sigma-Aldrich, Oakville, Canada) concentration was identified as 0.1 U/mL. A 1.0 M Tris buffer (pH buffering = 7.8 at 20°C) with a buffering capacity of 10 mM was added to control the pH elevation to within physiological range while still allowing crystal formation to occur. Finally, the presence of struvite and Ca phosphate crystals was confirmed with scanning electron microscopy (SEM; Zeiss/Leo 1540XB FESEM, Oberkochen, Germany) and X-ray diffraction (EDX; Oxford Instruments X Max 50, England, United Kingdom).
Testing urinary modulators
Urinary modulators were tested at concentrations that would be typically found in human urine. For compounds not normally found in human urine, a concentration of 1 mM was used. Ten milliliter of Griffith's artificial urine was mixed with 0.1 U/mL urease. Control reactions included Griffith's artificial urine with urease and 1.0 M Tris buffer (pH buffering = 7.8) and Griffith's artificial urine with Tris buffer only. Technical triplicates of 200 μL were pipetted into a 96-well plate (Sarstedt, Nümbrecht, Germany) and placed in the Biotek Eon Microplate Spectrophotometer (Winooski, VT) at 37°C and OD600 measurements were taken every 5 minutes for 24 hours using the Gen 5 v2.0 (Winooski).
After 24 hours, the pH levels were measured using the Mettler-Toledo FiveEasy Plus pH meter F20 (Gerifensee, Switzerland). If there was no evidence of crystal growth, then no further testing was performed on these modulators. If there was evidence of crystal growth, then a small sample was collected for microscopic evaluation and examined under a Nikon Eclipse Ts2R (Melville, NY) microscope.
Testing bacterial strains
Bacterial species tested for infectious stone formation were chosen based on a list curated by the Endourology Disease Group for Excellence consortium on the rates of bacterial strains present in struvite stones (Table 1). 11 After the bacteria were streaked and incubated at 37°C for 24 hours, a small amount of the bacteria was placed in 1.5 mL of Griffith's artificial urine in a microcentrifuge tube. The bacteria were then bead beaten with 100 μL of 100 μm glass beads for two cycles of 1-minute intervals at 3450 oscillations/minute while being placed on ice for 1 minute in between. After bead beating, the tubes were centrifuged at 20,817 × g for 5 minutes and aliquots of 200 μL were placed into a 96-well plate in technical triplicate.
List of Bacterial Strains Tested for Infectious Stone Formation with Experimental Model
The same postexperimental analysis was carried out with the bacterial strains including pH measurements and assessment of crystals under light microscopy. Crystal composition was determined with SEM and EDX. A qualitative test for urease productivity was performed with Christensen's agar as described hereunder.
Determining urease-producing bacteria
Christensen's agar is formulated to detect and differentiate urea degrading microorganisms. 12 The initial ingredients required to make Christensen's agar 12 was added to 900 mL of distilled water and was autoclaved for 15 minutes at 121°C. The solution was cooled down to 50°C–55°C and sterilized urea (final concentration 20 g/L) was added, and solution volume was adjusted to 1000 mL with sterilized distilled water. After the components were adequately mixed, the solution was poured out into individual agar plates to solidify and was stored at 4°C. To determine if the bacteria produced urease, a single colony was suspended in 1 mL of phosphate-buffered saline. A 10 μL aliquot was pipetted onto the agar plate and the plates were incubated at 37°C for 48 hours.
Statistical analysis
RStudio version 2022.02.1 (Boston, MA) using the Growthcurver package version 0.3.1 was used to analyze crystal growth curves. 13 Important output from growth curve analysis includes r, which measures the slope of the curve, t_mid, which corresponds with the time to reach 50% of the reaction, and area under the curve (AUC) is a surrogate for total crystal volume. Descriptive statistics and analysis of variance (ANOVA) with the growth curve results were performed using Prism (GraphPad, San Diego, CA). Prism—GraphPad was also used for all figures and graphs.
Results
The urinary modulators etidronic acid, ibuprofen, metformin, salicylic acid, citric acid, zinc sulfate, aspartic acid, and sodium citrate (Na citrate) yielded no crystals. Osteopontin (OPN) had a significant inhibitory effect on infectious crystal formation. Whereas indole, chondroitin sulfate, sodium bicarbonate (Na bicarbonate), vanillic acid, sodium pyrophosphate (Na pyrophosphate), polyaspartic acid (PASP), methionine, and heparin promoted infectious crystal formation, P-cresol was the only modulator that did not significantly impact overall crystal formation (Fig. 1).
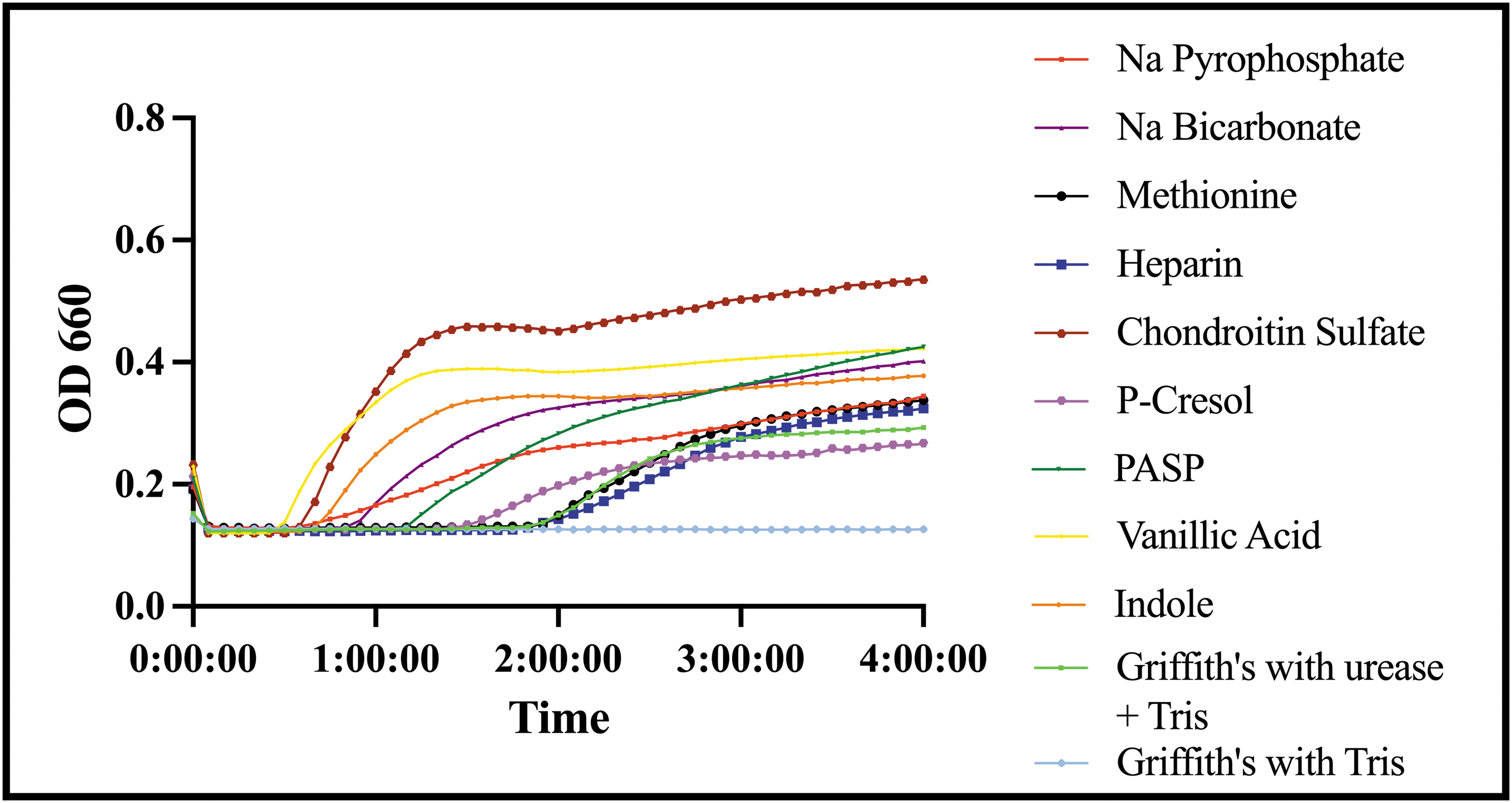
Crystal growth curves for modulators that promote crystal formation measured by absorbance at a wavelength of 660 nm over time in hours. Color images are available online.
Of the urinary modulators that were not associated with crystal formation, all maintained the initial acidic urinary pH conditions. The urinary modulators that were neutral or increased infectious crystal formation had similar or higher pH levels compared with the Griffith's urine with urease and Tris buffer (pH = 8.17). OPN had an end pH of 9.23, which is significantly higher compared with the baseline pH of the Griffith's urine with urease and Tris buffer. Despite this elevation in pH, the growth curve in table 2 analysis showed a less steep curve compared with the control (r = 0.008, t_mid = 447.929). Table 3 shows the AUC was substantially decreased with the addition of OPN (p < 0.0001). The crystals seen under light microscopy were struvite crystals with no obvious Ca phosphate crystals, which was unique compared with other modulators (Fig. 2).

Growth Curve Analysis for Urinary Modulators Allowing Crystal Formation
auc_e = empiric area under the curve; auc_l = logistic area under the curve; CS = chondroitin sulfate; k = growth capacity; n0 = degrees of freedom; OPN = osteopontin; PASP = polyaspartic acid; r = growth rate; Sigma = goodness of fit; t_gen = doubling time; t_mid = time to inflection point.
One-Way Analysis of Variance Comparing Empiric Area Under the Curve for Griffith's with Urease and Tris Compared with Each Urinary Inhibitor
p < 0.05 compared with Griffith's with urease and Tris.
CI = confidence interval.
The strains of bacteria that did not produce any crystals were Escherichia coli UTI89, Staphylococcus aureus USA300, Enterobacter cloacae A4-RS-21, Citrobacter freundii RCA +1, Staphylococcus agalactiae A3-MS-19, Enterococcus faecalis A2-FV-11, and Staphylococcus epidermidis A2-FV-12. Both Proteus strains caused the pH levels to rise well above 8 after 48 hours. In contrast, Klebsiella oxytoca, Klebsiella pneumoniae, and Pseudomonas aeruginosa had a pH range from 6.77 to 7.02 (Table 4).
pH Values of Crystal-Producing Bacterial Strains in Experimental Model
Both Proteus mirabilis strains had the steepest reaction curves with an early initiation of crystal formation supported by their greater r values and lower t_mid values (Fig. 3; Table 5). Both Klebsiella species had a similar shaped curve, but K. oxytoca (r = 0.006) had a lower r value compared with K. pneumoniae (r = 0.009), which means the development of crystals requires more time for K. oxytoca (Table 5). The empiric AUC for K. oxytoca was 52.266, which was the second highest of all bacterial species tested. Pseudomonas aeruginosa had a distinctive growth pattern with a bimodal curve. Owing to the earlier initiation of crystal formation, P. aeruginosa demonstrated the highest t_mid (1382.57), the greatest AUC, and highest growth capacity of all bacterial strains tested. An ANOVA comparing crystal growth from the bacterial strains showed a significant increase compared with controls with P. mirabilis 175A, P. aeruginosa, and K. oxytoca, but not K. pneumoniae and P. mirabilis 177A.

Crystal growth curves from crystal-producing bacterial strains measured by absorbance at a wavelength of 660 nm over time in hours. Color images are available online.
Growth Curve Analysis for Crystal-Producing Bacterial Strains
auc_e = empiric area under the curve; auc_l = logistic area under the curve; n0 = degrees of freedom; r = growth rate; Sigma = goodness of fit; t_gen = doubling time; t_mid = time to inflection point.
One-Way Analysis of Variance Comparing Empiric Area Under the Curve for Griffith's Only Compared with Bacterial Strains
p < 0.05 compared with Griffith's only.
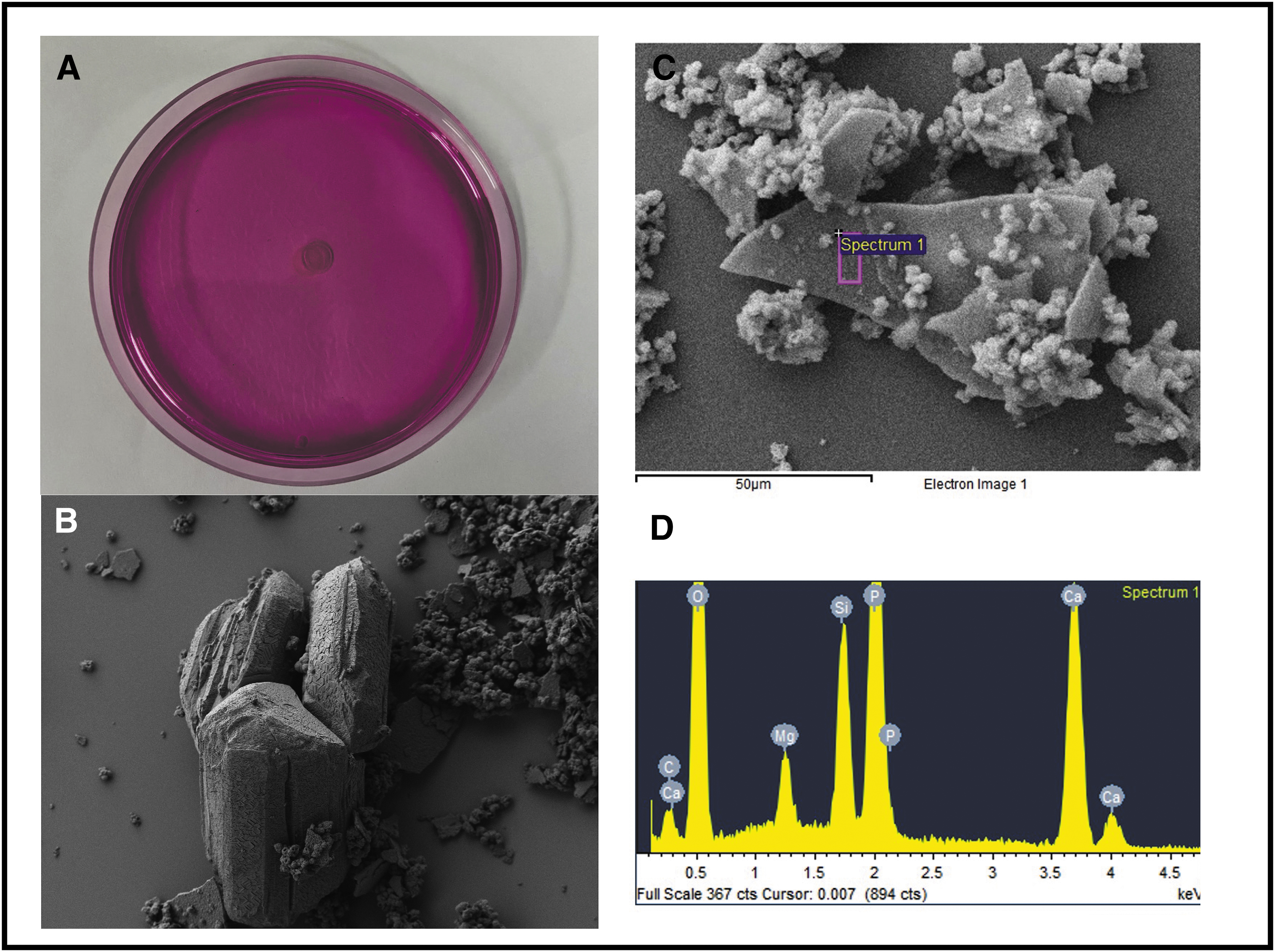
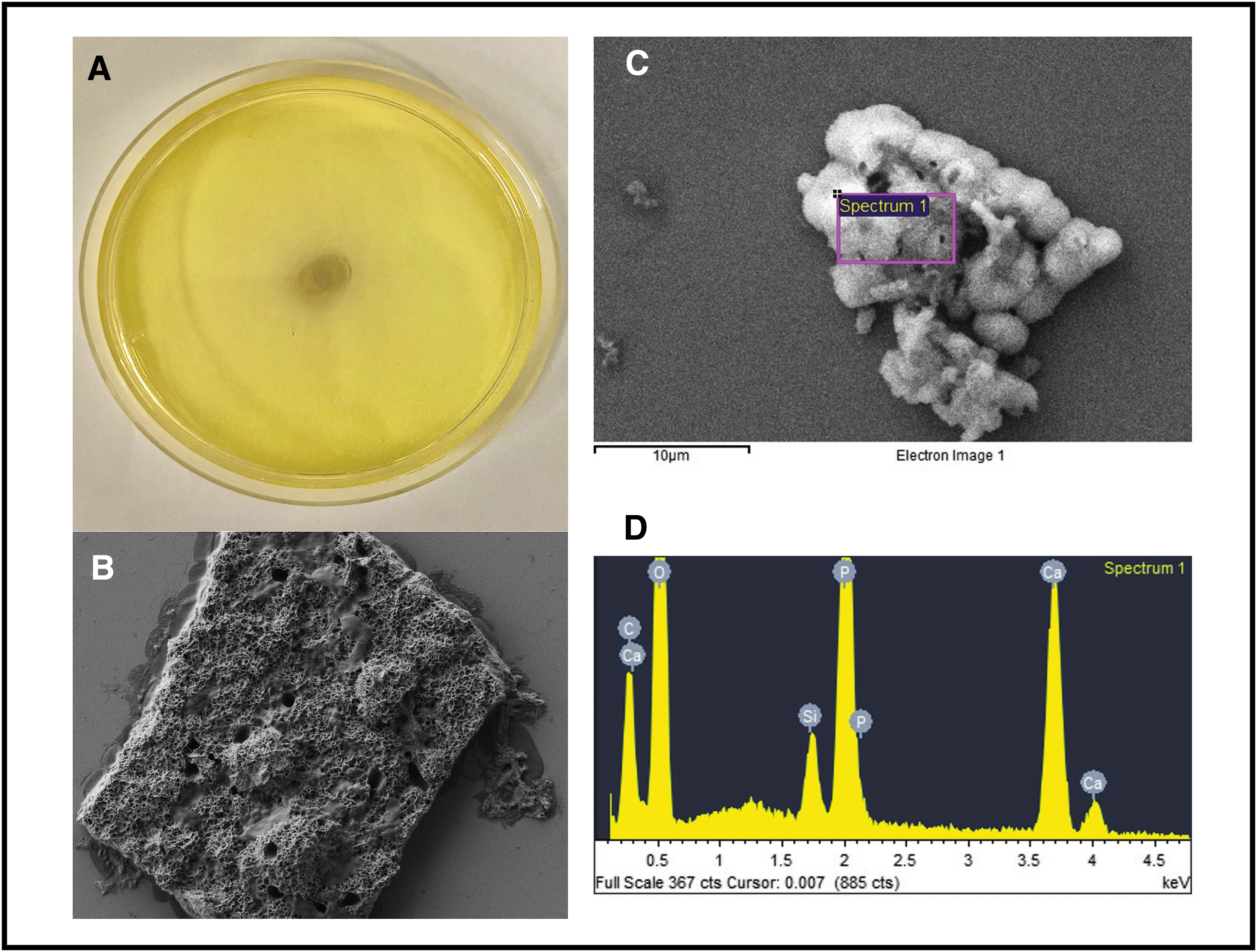
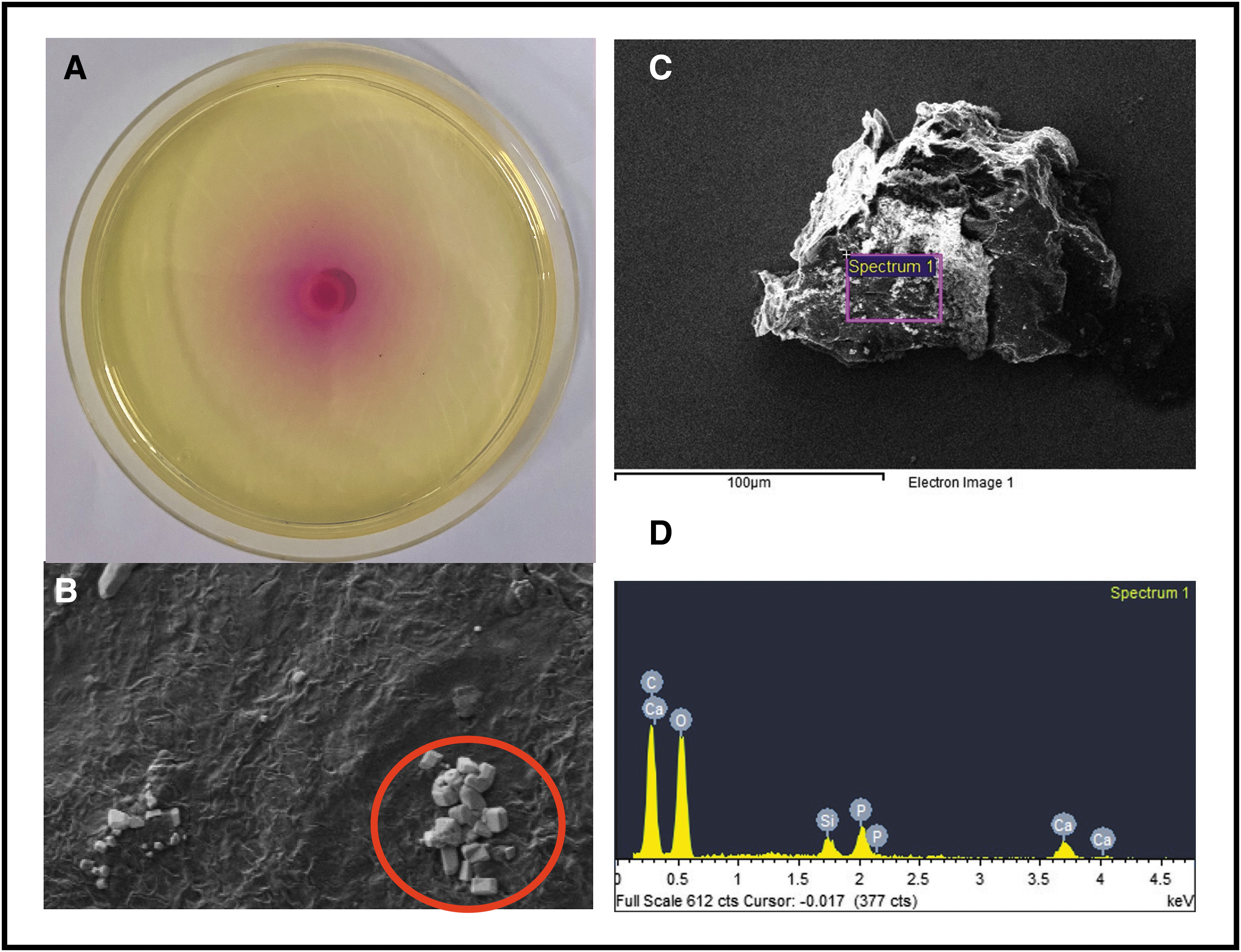
Figure 4 shows high urease productivity of P. mirabilis 175A with a completely pink Christensen's agar. The SEM for P. mirabilis 175A showed both Ca phosphate and struvite crystals, which was confirmed with EDX where there were large peaks for both phosphate and magnesium demonstrating a mixed composition of both stone types. Of interest, results from Christensen's agar showed no urease activity with P. aeruginosa; however, it still produced Ca phosphate crystals (Fig. 5). There was no magnesium present in the EDX analysis, showing a composition consistent with Ca phosphate crystals only and not struvite. Klebsiella oxytoca showed lower degree of urease activity with a smaller ring of pink around the bacterial colony. The encircled area of Figure 6B shows rectangular crystals, and analysis of crystal composition showed predominantly Ca, carbon (C), and oxygen (O) with no magnesium and very little phosphate, most consistent with a Ca oxalate stone composition. Overall, struvite crystal formation required a high concentration of urease, whereas Ca phosphate crystals formed without any urease. Ca oxalate crystals were observed to still form in the presence of urease.
Discussion
The urinary modulators demonstrated positive, negative, and mixed effects on infectious crystal formation. The pH level appears to have an important role, but an alkaline pH does not automatically equate to crystal formation. Not all bacterial strains in our experimental model yielded crystals. As anticipated, we observed that urease-producing bacteria (e.g., P. mirabilis) led to an alkaline pH and infectious stone formation, but the more intriguing results came from the other crystal forming bacterial strains that did not reach the same pH levels. The results from K. oxytoca showed that the bacterium can promote noninfectious stones in the presence of urease and P. aeruginosa can promote Ca phosphate stones without urease.
The urinary inhibitors that completely prevented any crystal formation were predominantly acids. This effect could have been owing to the degradation of the urease enzyme or another mechanism preventing a rise in pH. Metformin in our experiment showed complete inhibition of crystal formation, and previous studies with in vitro and in vivo models have shown metformin reduces crystal formation through antioxidant effects. 14 Zinc has previously shown a propensity to improve Ca aggregation; however, in our study zinc sulfate completely inhibited crystal formation suggesting that sulfate is the more dominant modulator and prevents the formation of crystals. 15,16 Citrate has been shown to inhibit urease-induced crystallization and delay the onset of nucleation and growth of struvite crystals. 17 Our results support the importance of citrate as a stone inhibitor and somewhat counterintuitively given its effect on raising urinary pH. Citrate may be an underutilized clinical treatment strategy in infectious stones. OPN did not completely inhibit crystal formation, but delayed the onset of the formation of crystals, slowed the reaction, and significantly decreased the overall crystal volume. This is consistent with other studies showing its inhibitory effect on Ca oxalate and Ca phosphate crystal formation. 18 The challenge with OPN as a potential therapeutic strategy is that upregulation in the kidney is mediated by inflammation and healing. 19
The results from the bacterial strains within the experiment were quite intriguing. Only the P. mirabilis strains raised the pH owing to urease production resulting in subsequent Ca phosphate and struvite crystal formation. However, P. mirabilis 177A did not increase crystal volume enough to reach statistical significance in our analysis. Despite not producing a very high pH or any urease, P. aeruginosa was able to induce Ca phosphate crystals. Struvite crystals will not form with a pH <7.0, so their absence is expected. 20 Bacteria with a lower level of urease productivity that cannot raise the pH above this threshold may only form Ca phosphate stones. Klebsiella oxytoca was able to produce Ca oxalate stones even in the presence of urease, which was not shown in any other experimental conditions. Chutipongtanate et al showed a positive link with Ca oxalate crystallization and all bacteria including E. coli, K. pneumoniae, S. aureus, and S. pneumoniae. 21 Our results suggest that there may be bacterial intracellular proteins that are able to drive these crystal forming reactions besides urease and that overall urease productivity may determine ultimate stone composition.
The strengths of this study are that the experimental conditions were standardized and testing in artificial urine ensured that the effects demonstrated were solely related to the modulator. We tested many inhibitors, and we based the test concentrations of normal human urine concentrations when possible. Limitations of this study are the effect of some of the urinary modulators may be concentration dependent and we only tested one concentration for each modulator. We cannot account for urinary modulators found in human urine that were not incorporated into our model, which could alter the results of the urinary modulators. Therefore, the results of this study may not hold true in the in vivo setting and further clinical research is required.
Successful urinary acidification appears to be the most successful way to treat infection-based stones and further research will be required to identify a successful treatment strategy to be used in patients. Central to prevention of infection-related urinary stones also includes more effective strategies at reducing the occurrence of urinary tract infections without disrupting the urinary microbiome.
Conclusions
The results of this study suggest that infectious stone formation is a more complex clinical entity than historically believed. Urinary modulators appear to have an important role, and their potential therapeutic clinical impact warrants further investigation.
Footnotes
Authors' Contributions
B.W.—conceptualization, methodology, investigation, formal analysis, writing original draft, writing review and editing. J.A.C.—conceptualization, methodology, validation, writing review and editing. K.F.A.—conceptualization, methodology, validation, writing review and editing. J.B.—conceptualization, methodology, writing review and editing, visualization. J.P.B.—conceptualization, methodology, resources, writing review and editing, visualization. H.G.—conceptualization, methodology, writing review and editing. H.R.—conceptualization, methodology, writing review and editing, visualization.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.