
Abstract
The molecular basis and evolution of multidrug resistance were established for 54 isolates of Salmonella enterica serovar Ohio, recorded between 1991 and 2005 in Asturias, a northern region of Spain. All isolates were closely related, as shown by cluster analysis of XbaI-BlnI combined profiles. Of these, 33.3% were resistant to one or more unrelated agent(s). Sulphonamides, streptomycin, tetracycline, and trimethoprim, encoded by sul1, aadA1, tet(A) or tet(B), and dfrA1, respectively, were the most common resistances, but ampicillin (bla TEM-1), gentamicin (aacC2 or aacC4), kanamycin (aphA1), and chloramphenicol (catA1) were also detected. Two types of complex genetic elements, carried by large conjugative or mobilizable plasmids, were found in isolates resistant to four or more unrelated agents (multidrug resistant), which accounted for 18.5% of the total: (i) a class 1 integron (1600 bp/dfrA1-aadA1) close to a defective Tn10, both inserted within a Tn21-like element that was carried in some cases by Tn9; (ii) Tn3-bla TEM-1, which was inserted within a defective Tn1721. These elements have been involved in the development and spread of multidrug resistance in S. enterica Ohio, which was detected between 1994 and 2001. The absence of multidrug resistance in later years could have been connected with the European Union strategies for combating antimicrobial resistance and controlling nontyphoid S. enterica in food-producing animals.
Introduction
M
In a previous study we detected a relatively high incidence of multidrug resistance in S. enterica Ohio, with 25.5% of the clinical isolates tested being resistant to from four to seven antimicrobial agents. The isolates have been recovered from different Spanish regions, including Asturias, between 1990 and 1998 (Soto et al., 2000). Although the number of studies on this serovar are scarce, multidrug resistance has also been observed in S. enterica Ohio from pigs, foods, and environmental samples (Nastasi and Mammina, 2001; Agustín et al., 2005; Zhao et al., 2006). Our investigation was aimed to establish the molecular basis of multidrug resistance in the S. enterica Ohio isolates recovered in Asturias during the 1991–1998 period and to track the trends of resistance by extending the study to all isolates of the same serovar recorded from 1999 to 2005 in the same region.
Materials and Methods
Bacterial strains
Fifty-four isolates of S. enterica Ohio all recorded at the Laboratory of Public Health (LSP) of Asturias, Spain, were included in this study (Table 1). Thirty two of them, corresponding to the 1991–1998 period, had been partially characterized in a previous study (Soto et al., 2000). The remaining 22 were all the S. enterica Ohio isolates recorded during 1999–2005 in the indicated region. The isolates were collected from the faeces of 49 patients with gastroenteritis, of which 25 were males and 24 females (Table 2); 55.1% were from infants or children (1 month to 13 years of age), and 14.3% were from elderly people (67–85 years old). Three infants suffered from episodes of persistent diarrhoea, and Salmonella was obtained in 2 and 4 consecutive months, and in 2 nonconsecutive months, respectively. The latter two isolates, recovered from a child with less than 1 year of age, and one isolate from his 2-year-old brother were associated with a family outbreak that occurred in May 1995. A second outbreak, undetected at the time (January 2003), was revealed by our study.
pUO-SoR, plasmid University of Oviedo–Salmonella Ohio Resistance.
Small (ca. 5 kb or less) cryptic plasmids were detected in some isolates (Soto et al., 2000).
Recovered from a child under 1 year of age in 4 consecutive months.
Recovered from a child under 1 year of age in 2 consecutive months.
Not registered outbreak.
Recovered from a child under 1 year of age in 2 nonconsecutive months.
Family outbreak
R, resistance; N, number of isolates with the indicated profile; PFGE, pulsed-field gel electrophoresis; LSP, Laboratory of Public Health (Asturias, Spain); AMP, ampicillin; CHL, chloramphenicol; GEN, gentamicin; KAN, kanamycin; STR, streptomycin; SUL, sulphonamides; TET, tetracycline; TMP, trimethoprim; −, negative for the indicated element(s).
ND, no data available.
Antimicrobial susceptibility testing and detection of resistance determinants
Susceptibility to antimicrobial agents was determined by the disc diffusion assay on Mueller–Hinton agar with commercially available discs (Oxoid, Madrid, Spain), according to Clinical and Laboratory Standards Institute (CLSI, 2005, 2006). The following antimicrobials were tested: ampicillin, chloramphenicol, gentamicin, kanamycin, nalidixic acid, streptomycin, sulphonamides, tetracycline, and trimethoprim. Resistance determinants [aadA1-like, aphA1, aacC2, aacC4, bla TEM, catA1, cmlA1-like, dfrA1-like, dfrA12, strA, strB, sul1, sul2, sul3, tet(A), tet(B), and tet(G)] were screened by polymerase chain reaction (PCR) amplification using previously described primers and conditions (Rodríguez et al., 2006a; Martínez et al., 2007). The primers used for aadA1-like, dfrA1-like, and bla TEM are not each specific for a single gene, but the identity of the obtained amplicons was established by sequencing (Secugen, Madrid, Spain).
Integron and transposon analysis
Integrons were screened by PCR amplifications performed with degenerate primers (hep35 and hep36) designed for conserved regions of classes 1, 2, and 3 integrase genes (intI1 to intI3), followed by digestion of the generated products with HinfI (White et al., 2001). The variable regions of class 1 integrons were amplified with the 5' conserved segment (CS) and 3'CS primers that anneal with sequences flanking the attI1 site. Gene cassettes in the variable regions were identified by nested PCR with primer pairs selected on the basis of the R-genotypes of the integron-containing isolates, followed by sequencing. The presence or absence as well as the arrangement of genes outside the variable region of the integrons, including intI1, qacEΔ1, orf5, istA and istB (of IS1326), orfA and orfB (of IS1353), tniBΔ1, and tniA (Liebert et al., 1999), were also determined by PCR (Rodríguez et al., 2006b; Martínez et al., 2007). Transposon-related sequences were screened with primers targeting IS1, the insertion sequence flanking Tn9 (Liebert et al., 1999); tnpA, tnpR (encoding transposition functions), and mer genes (involved in mercury resistance) of Tn21 (Liebert et al., 1999); ybdA (encoding a protein with homology to transcriptional repressors of metal resistance operons), ybfA (for a possible sodium/glutamate transporter), tetC, tetD (the latter specifying a transcriptional activator of genes that confer resistance to redox-cycling compounds and antibiotics, negatively regulated by the product of tetC), tetR [encoding the tet(B) repressor], and tnp (for the transposase of IS10), all belonging to Tn10 (Coleman et al., 1983; Griffith et al., 2005); mcp (which encodes a polypeptide with features of a methyl-accepting chemotaxis protein), tnpR, tnpA (transposase functions), and pecM (putative regulatory component), specific of Tn1721 (Allmeier et al., 1992; Schlüter et al., 2003); and tnpA of Tn3 (which encodes the transposase; Heffron et al., 1979). Physical associations between resistance genes, integrons, and/or transposons were established in all isolates in which the latter elements were detected (Table 1; Figs. 1 and 2) (Rodríguez et al., 2006b; Martínez et al., 2007).
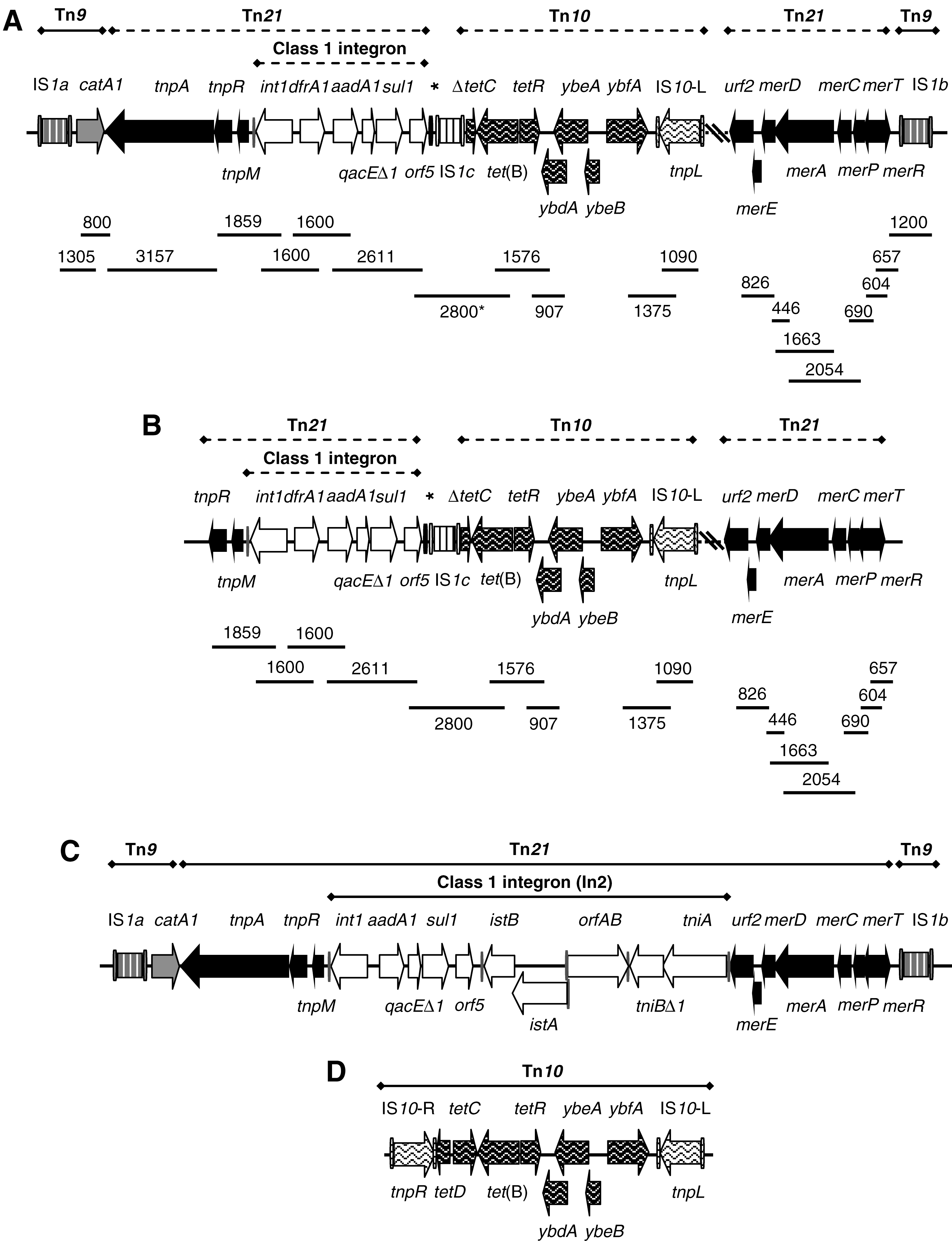
Organization of the class 1 integron, and the Tn21- ± Tn9-related transposons associated with the defective Tn10 in tet(B)-positive isolates of Salmonella enterica serovar Ohio. (
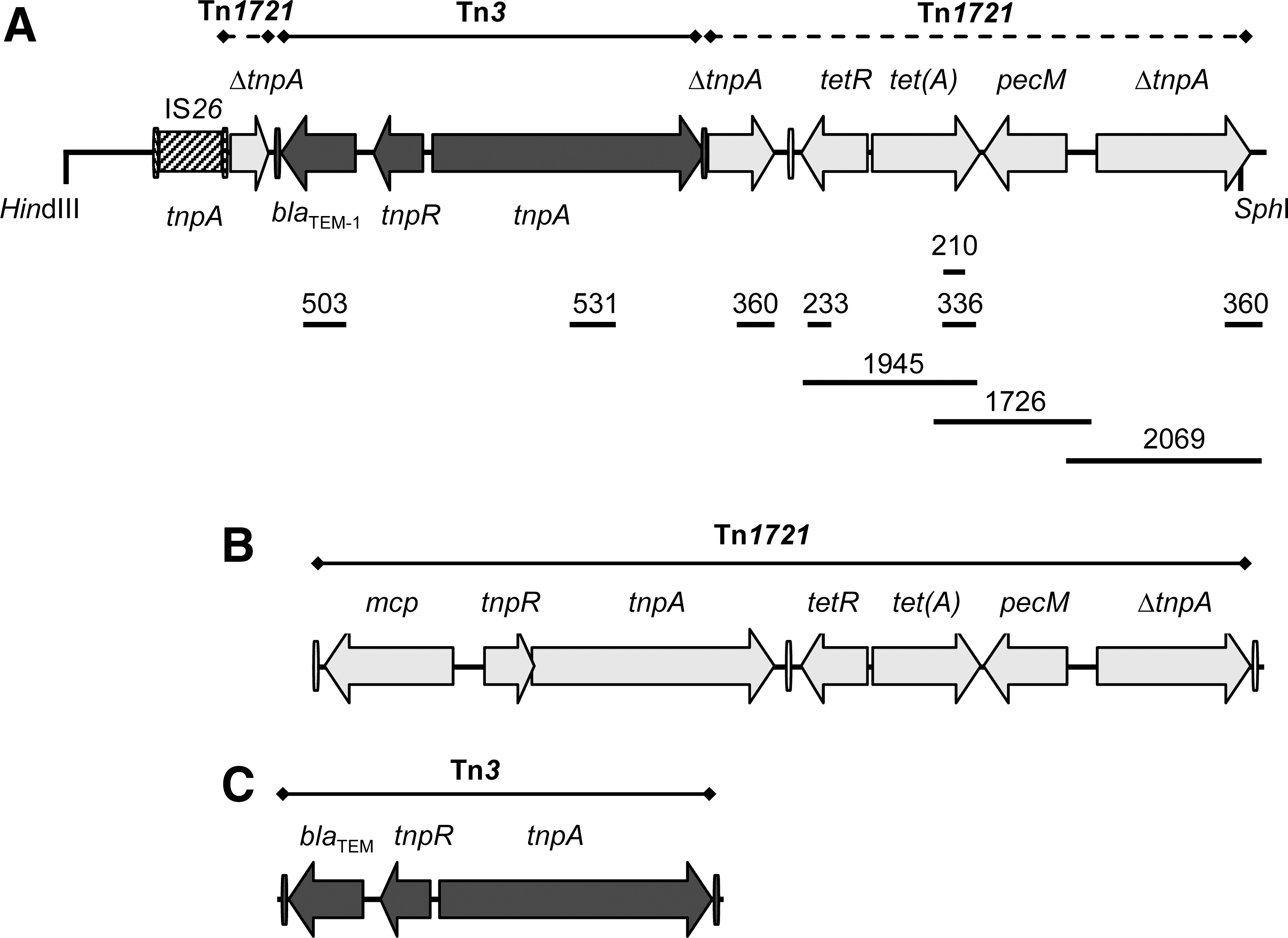
Organization of the Tn1721-Tn3-like transposons, carried by pUO-SoR1 in bla
TEM-1- and tet(A)-positive isolates of S. enterica serovar Ohio (
Plasmid analysis and conjugation experiments
To determine the number and molecular size of large plasmids, total DNA of S. enterica Ohio was digested with S1 nuclease (Amersham Biosciences, Barcelona, Spain) and subjected to pulsed-field gel electrophoresis (PFGE) (S1-PFGE) (Martínez et al., 2005). Lambda ladder PFG marker (New England BioLabs, Beverly, MA) was the size standard. Conjugation experiments were performed in Luria broth (Sambrook and Russel, 2001) at 37°C, using Escherichia coli K12 J53 (resistant to rifampicin) as recipient (Martínez et al., 2005). Two nonconjugative R-plasmids were mobilized from S. enterica Ohio into E. coli K12 J53 by triparental mating, using the E. coli HB101 helper strain with plasmid pRK600 that confers resistance to chloramphenicol, as described by Kessler et al. (1992). Transconjugants were selected on eosin methylene blue agar (Oxoid) containing rifampicin (50 mg/L) along with ampicillin (100 mg/L), kanamycin (30 mg/L), or tetracycline (30 mg/L). Transfer of resistance genes was phenotypically tested by the disc diffusion method, and drug resistance determinants were screened by PCR, as earlier. The identified R-plasmids were termed pUO-SoR (plasmid of University of Oviedo–Salmonella Ohio Resistance), followed by a serial number, and their presence in the E. coli transconjugants was also confirmed by S1-PFGE.
Cloning and sequence analysis of the tet(A) region
The tet(A) gene was cloned from plasmid pUO-SoR1 into the pUC18 cloning vector, using standard techniques (Sambrook and Russel, 2001). In the successful strategy, pUO-SoR1 was extracted from an E. coli transconjugant of S. enterica Ohio LSP334/94, using the HiPure Plasmid Midiprep Kit (Invitrogen, Barcelona, Spain), simultaneously digested with SphI and HindIII (Takara Biomedicals, Saint-Germain-en-Laye, France), and then ligated to the vector previously cut with the same enzymes. The ligated DNA was transformed into chemically competent E. coli DH5a cells (Invitrogen), and direct selection was achieved in Luria broth containing both ampicillin and tetracycline, at the above indicated concentrations. The cloned DNA fragment conferring resistance to tetracycline was sequenced and analyzed by BLAST (Altschul et al., 1997).
Molecular typing by macrorestriction PFGE
Genomic variability of the isolates was assessed by macrorestriction-PFGE. DNA-containing plugs were independently digested with XbaI and BlnI endonucleases (30 U, 4 h, 37°C; Takara Biomedicals) and the fragments obtained were separated in a CHEF-DRIII system (Bio-Rad Laboratories, Madrid, Spain). Agarose plug preparation and PFGE running conditions were performed using the standardized PulseNet protocol (
Results
Antimicrobial drug resistance in S. enterica Ohio and genetic basis
When S. enterica Ohio isolates were tested for antimicrobial resistance and the responsible genes identified, 11 profiles, termed S and R1 to R10, were detected (Table 1). Isolates susceptible to all compounds tested were the most frequent (66.7%; S profile), while the remaining were resistant to one (11.1%), two (1.9%), three (1.9%), and four or more (18.5%) antimicrobial agents, the latter considered as multidrug resistant. The most common resistances were sulphonamides, streptomycin, and tetracycline (shown by 29.6, 20.4, and 18.5% of the isolates, respectively), followed by ampicillin and trimethoprim (14.8% each), gentamicin and kanamycin (7.4% each), and chloramphenicol (3.7%). Resistance to nalidixic acid was not detected, in contrast to other serovars found in poultry, such as S. enterica Enteritidis and S. enterica Hadar (Threlfall, 2002). With respect to the resistance genotypes, (i) in all cases, resistance to ampicillin, chloramphenicol, kanamycin, sulphonamides, and trimethoprim were encoded by bla TEM-1, catA1, aphA1, sul1, and dfrA1, respectively; (ii) three and one gentamicin-resistant isolates were positive for accC2 and accC4, respectively; (iii) streptomycin-resistant isolates collected before 1996 carried strA-strB, whereas those recovered after this year were positive for aadA1; (iv) similarly, tetracycline-resistant isolates contained tet(A) or tet(B), depending on whether they were collected before or after 1996; and (v) the remaining resistance genes checked for were not detected.
Integrons and transposons in multidrug-resistant isolates of S. enterica Ohio
Integrons of classes 2 and 3 were not found in S. enterica Ohio. However, six multidrug-resistant isolates (with profiles R8 to R10) proved to be positive for class 1 integrons with the dfrA1-aadA1 gene cassette configuration in the variable region of about 1600 bp (Table 1). When the detailed structure of these integrons was investigated (Fig. 1A, B), all proved to be positive for intI1, qacEΔ1, sul1, and orf5. However, they were negative for IS1326 and IS1353, the two insertion sequences characteristic of In2, the integron of Tn21, and for the tniA and tniBΔ1 genes, which encode a defective transposition module, widely represented in class 1 integrons (Fig. 1C).
With respect to the multidrug-resistant profiles associated with class 1 integrons (Table 1), the following results are of note: (i) in two chloramphenicol-resistant isolates (R10 profile), the integron was part of a Tn21-like transposon that carried apparently intact tnp genes (tnpA, tnpR, and tnpM), as well as the mer operon, and was in turn inserted within Tn9 that supplied the catA1 gene (Fig. 1A); (ii) in four chloramphenicol-susceptible strains with R8 or R9 profiles, the integron was inserted within a defective Tn21-like transposon that had a deletion affecting the tnpA gene but maintained tnpR, tnpM, and the mer operon (Fig. 1B); and (iii) the tet(B)-positive isolates of profiles R8, R9, and R10 contained the transposase gene of IS10 (the insertion sequence flanking Tn10), and in all of them the linkages tet(B) and tetR, tetR and ybdA, as well as ybfA and tnpL (the latter encoding the transposase of the left copy of IS10) were demonstrated (Fig. 1A, B). However, the amplicons joining tetC with tetD and tet(B) with tnpR could not be obtained. All tet(B)-positive isolates carried the 1600 bp/dfrA1-aadA1 integron inserted within a Tn21-like element, which was associated or not with Tn9 (see earlier). Based on results previously obtained for S. enterica Brandenburg strains with similar characteristics and from the same Spanish region (Martínez et al., 2007), we amplified a fragment of about 2800 bp joining the tet(B) gene of Tn10 with orf5 of the integron in the six tet(B)-positive isolates. Sequencing of the fragment from one of the isolates (LSP191/98) revealed a copy of IS1 placed between orf5 of the class 1 integron (that maintained one of the inverted repeats of IS1326) and a Tn10-related element, which lacked tetD and part of tetC (Fig. 1A, B). This organization corresponds with that previously reported for S. enterica Brandenburg.
In relation to the multidrug-resistant profiles negative for class 1 integrons (R5, R6, and R7; Table 1), (i) all isolates were ampicillin resistant and contained a Tn3-like transposon that carried bla TEM-1, and this element was not found in ampicillin-resistant isolates of the remaining profiles; and (ii) all were positive for the tet(A) gene characteristic of Tn1721. The presence of the tetR, tet(A), pecM, and ΔtnpA (which corresponds to a partial duplication of tnpA) genes of this transposon (Fig. 2B) was confirmed by PCR amplification of the individual genes (Fig. 2A). However, mcp, tnpR, and an entire tnpA gene could not be detected. These results, together with overlapping PCR amplifications, indicated the presence of a defective Tn1721 in the isolates tested. The detailed structure of the element was then established for LSP334/94 by cloning and sequencing experiments. These revealed the presence of IS26 adjacent to a truncated tnpA gene, which has suffered a 1450 bp deletion at the 5'-end, while confirming the absence of mcp and tnpR. Moreover, a second deletion removed 430 bp internal to tnpA, which have been substituted by the Tn3-bla TEM-1 element.
Plasmids in multiresistant strains of S. enterica Ohio
Four types of resistance plasmids, termed pUO-SoR1 to pUO-SoR4, were detected by S1-PFGE in multidrug-resistant isolates of S. enterica Ohio (Tables 1 and 3; Fig. 3). pUO-SoR1 and pUO-SoR4 were both conjugative plasmids of about 190 kb. The former was present in isolates of the R5, R6, and R7 profiles but carried only two of the resistance genes present in the parental isolates, specifically bla TEM-1 and tet(A), which are contained in the Tn3-Tn1721 element (Fig. 2A). pUO-SoR4, present only in R10 isolates, transferred the same resistance pattern detected in its hosts, when conjugated into E. coli. This pattern was justified by the complex element resulting from the association of Tn21-like transposons carrying the 1600 bp/dfrA1-aadA1 integron, with Tn9- and Tn10-like transposons (Fig. 1A). Finally, pUO-SoR2 (ca. 145 kb) and pUO-SoR3 (ca. 160 kb) found in R8 and R9 isolates, respectively, could only be transferred into E. coli by triparental mating. As a result, all the transconjugants obtained contained either pUO-SoR2 or pUO-SoR3 together with pRK600, the chloramphenicol-resistant plasmid present in the helper strain. Such transconjugants showed the same resistance phenotype and genotype, class 1 integron, and transposons (Tn21- and Tn10-like) as the respective parental strains (Fig. 1B), in addition to catA1 carried by pRK600 (not shown).

Plasmid profiles of representative S. enterica serovar Ohio isolates and Escherichia coli transconjugants visualized by pulsed-field gel electrophoresis (PFGE) after S1 nuclease digestion. (
The helper plasmid pRK600, which carries the catA1 gene for chloramphenicol resistance, provided the functions for mobilization of pUO-SoR2 and pUOSoR3.
The percentage was calculated with respect to the total number of LSP isolates (54) included in our study.
R, resistance; C, conjugation; M, mobilization; N, number of isolates carrying the indicated plasmid; +, positive; −, negative; nt, not tested.
Genomic typing of S. enterica Ohio by macrorestriction PFGE
Genomic macrorestriction identified 10 XbaI-PFGE profiles (X1–X10; Fig. 4A) and six BlnI-PFGE patterns (B1–B6; Fig. 4B). Of these, X1 and B2 (59.3% each) were the most common. Accordingly, the prevalent combined profile was X1B2, shown by 27 isolates (50%) scattered along the 1992–2005 period. Of these, 24 were susceptible to all antimicrobials tested, and three were resistant to sulphonamides. The latter were involved in a family outbreak, while eight X1B2 susceptible isolates, recovered in January 2003 in adjoining health areas, could have been associated with a community outbreak, undetected at the time. The 10 multidrug-resistant isolates displayed one of four XbaI-BlnI profiles (X2B2, X3B4, X6B5, or X7B5). Those containing pUO-SoR1 had the same X3B4 profile, shared also by a single isolate carrying pUO-SoR2. Two additional pUO-SoR2 isolates were X7B5, a profile closely related to X6B5, shown by the only isolate positive for pUO-SoR3 and by one of the two isolates carrying pUO-SoR4. The second isolate with the latter plasmid was X2B2. It is finally worth noting that when a dendogram of similarity was constructed with the XbaI-BlnI combined profiles (Fig. 4c), all S. enterica Ohio isolates grouped in a single cluster at a cutoff point of S= 0.74, indicating a close relationship. Moreover, the X1B2 clone can be regarded as endemic, because it was detected from 1992 to 2005.

Analysis of S. enterica serovar Ohio isolates by PFGE. (
Discussion
The plasticity of the bacterial genome underlies the amazing adaptability of bacteria to hazardous environments, such as those imposed by the presence of antimicrobial agents. Resistance genes have an exceptional ability to become associated with each other, helped by different types of genetic elements that greatly contribute to their recruitment, expression, persistence, and dissemination. The resulting clusters, known as resistance islands, are often responsible for the simultaneous resistance to multiple drugs, which may greatly complicate the treatment of important pathogens, including S. enterica (Miriagou et al., 2006; Walsh, 2006).
In S. enterica Ohio, a class 1 integron and four transposons (Tn21-, Tn9-, Tn10-, and Tn1721-related), with their associated resistance genes, have coincided with a number of other genes to confer multidrug resistance. All these elements and most of the resistance genes were carried by large conjugative or mobilizable plasmids. The integron of S. enterica Ohio was identified by the 1600 bp/dfrA1-aadA1 variable region, a gene cassette configuration that is widely distributed in S. enterica (Daly et al., 2004; Antunes et al., 2006; Miriagou et al., 2006; van Essen-Zandbergen et al., 2007). In Spain, it has been previously found in the following Salmonella enterica serotypes: Brandenburg, Enteritidis, Panama, Typhimurium, Wien, and Virchow, apart from Ohio (Guerra et al., 2000; Soto et al., 2000, 2003; Martínez et al., 2007; Rodríguez et al., 2008). However, only the integrons from a number of S. enterica Brandenburg isolates recorded in Asturias between 1995 and 2002 have been characterized in detail (Martínez et al., 2007). They shared the structure of those reported for S. enterica Ohio in our work, which consisted of a “minimal” integron platform, constructed by the variable region, flanked by the 5'CS with the integrase gene, and the 3'CS with qacEΔ1, sul1, and orf5. Interestingly, all S. enterica Ohio and Brandenburg isolates with this “minimal” integron were resistant to tetracycline because of the presence of tet(B), and a defective Tn10 carrying this gene was found in the proximity of the integron, separated from orf5 by a copy of IS1. The resulting structure was inserted within Tn21- ± Tn9-like transposons in the two serovars, located on large conjugative or mobilizable plasmids, termed pUO-SoR2 to pUO-SoR4 in the case of S. enterica Ohio, and pUO-SbR4 and pUO-SbR5 in S. enterica Brandenburg (Martínez et al., 2007). The possible relationship existing between these plasmids will be the subject of further investigation. Those of S. enterica Ohio were found to be established in four genetic backgrounds (X2B2, X3B4, X6B5, and X7B5, detected between 1996 and 2001) and evolved either by acquisition of additional resistance genes (bla TEM-1 and/or aphA1) or by lost of gene(s) from a larger structure.
Class 1 integrons were not detected in tetracycline-resistant isolates of S. enterica Ohio which carried tet(A). They were all resistant to ampicillin and contained Tn3 associated with a defective Tn1721, which lacked mcp and tnpR and has two deletions affecting tnpA. Different versions of Tn1721 have been previously identified in other serovars, mainly in S. enterica Typhimurium from various sources and countries and also in S. enterica Brandenburg collected in Asturias from clinical samples (Frech and Schwarz, 1999; Pezzella et al., 2004; Pasquali et al., 2005; Martínez et al., 2007; Doublet et al., 2008; Hradecka et al., 2008; Doublet et al., 2009). Like S. enterica Ohio, some S. enterica Typhimurium isolates carried a defective Tn1721 associated with Tn3-bla TEM. In one of them, recovered from a rabbit at a slaughterhouse in Italy, the conventional left arm of Tn1721 has been replaced by Tn3, whereas in two isolates from the Czech Republic the latter transposon was inserted within the pecM gene (also known as orf294) of Tn1721 (Pasquali et al., 2005; Hradecka et al., 2008). The three different structures described so far reveal a nonspecific affinity of Tn3 for Tn1721. The Tn1721-Tn3 element of S. enterica Ohio was carried by pUO-SoR1, a conjugative plasmid only recorded in 1994, associated with the X3B4 PFGE profile. In this case, intraclone evolution has taken place through loss and/or acquisition of resistance genes other than those provided by the plasmid [bla TEM-1 and tet(A)]. The additional genes [aacC2, dfrA1, strA-strB, and sul1] are probably located on the bacterial chromosome.
Regarding the trends of multidrug resistance, susceptible isolates of S. enterica Ohio were distributed throughout the period of study (1991–2005), whereas multidrug-resistant isolates were only found between 1994 and 2001. The absence of multidrug resistance in later years is an encouraging finding, which can be connected with the European Union strategies for controlling nontyphoid S. enterica in food-producing animals and for combating antimicrobial resistance (CEC, 2001; European Union, 2003; Smulders et al., 2008). In fact, a decrease in the total number of salmonellae that reach humans through the food chain will result in a concomitant decrease in the number of resistant isolates, particularly if the selective pressure posed by the inappropriate use of antimicrobials both in humans and animals is diminished.
Footnotes
Acknowledgments
This work has been supported by project FIS PI080656 of the Ministerio de Ciencia e Innovación, Spain. I.R. is the recipient of a grant from the Fundación para el Fomento en Asturias de la Investigación Científica Aplicada y la Tecnología (FICYT-BP04-086). M.R.R. and M.C.M. are members of a research unit associated with the Consejo Superior de Investigaciones Científicas, Spain. The authors are grateful to M.A. González-Hevia and M. Bances (Laboratorio de Salud Pública, Oviedo, Spain) for the isolates used in this work and to them and A. Echeita (Laboratorio Nacional de Referencia de Salmonella y Shigella, Instituto de Salud Carlos III, Madrid, Spain) for providing information on the trends of Salmonella enterica in Asturias and Spain.
Disclosure Statement
No competing financial interests exist.