
Abstract
Cholera, travelers' diarrhea, or colibacillosis in pigs can possibly be prevented or attenuated by dietary provision of competitive inhibitors that react with the GM1-binding sites of the enterotoxins cholera toxin (CT), human Escherichia coli heat-labile enterotoxin of serogroup I (LTh-I), and porcine LT-I (LTp-I). The interfering efficiency of natural substances with binding of the toxins to the gangliosid receptor GM1 was tested using a specially adapted GM1-coated-microtiter-well enzyme-linked immunosorbent assay. The substances tested for their GM1 displacing capacity were galactose-containing or -related saccharides from bovine milk, skim milk powder, galactan from gum arabic, food stabilizers as well as ground fenugreek seed and soy bean constituents that contain galactomannans, the galactopolysaccharides agar and agarose, and larch wood and other plant materials that contain arabinogalactans. Skim milk powder, compared with the pure milk saccharides tested, interfered to a higher extent with LTh-I (65–66% inhibition at 5 mg test substance/mL) and CT binding (63–67% inhibition at 5 mg test substance/mL) when supplied before or simultaneously with the toxins in the GM1-enzyme-linked immunosorbent assay. Ground fenugreek seed counteracted GM1 binding of 5 ng LTh-I/mL as well as 5 ng and 1 μg LTp-I/mL (43–65% inhibition at 5 mg test substance/mL), and 4 ng CT/mL (61–92% inhibition at 5 mg test substance/mL) very efficiently when supplied before the toxin–GM1 complex had formed. With 50 mg/mL fenugreek seed, inhibition percentages of even 92–99% were reached for LTh-I and CT binding. Efforts to resolve already bound toxin from GM1 with the test substances were less effective than preincubations and concurrent incubations.
Introduction
E. coli LT-I is very similar to cholera toxin (CT) in amino acid sequence, in mode of action, and in biological properties (Sixma et al., 1993; Hol et al., 1995; Nataro and Kaper, 1998). Both toxins have an AB5 composition, in which the B pentamer has a membrane-binding function and the A subunit exerts enzymatic activity after cleavage (Hol et al., 1995). The five B subunits of LT-I are arranged in a plane donut shape, mediating strong host cell attachment via ganglioside GM1 and, much weaker, via GD1b and some intestinal glycoproteins (Teneberg et al., 1994).
Group I heat-labile enterotoxins of E. coli comprise two closely related antigenic variants, called LTp-I and LTh-I, after their initial discovery in strains isolated from pigs and humans, respectively (Tsuji et al., 1982). Group II contains type II heat-labile enterotoxins, which have been associated with diarrhea, but have not been positively identified as disease-causing agents in animals or humans so far (Van den Akker et al., 1996; Nardi et al., 2005).
The genes for the diarrhea-inducing LT-I are located on plasmids in E. coli (Gyles et al., 1974; Smith, 1977) and are hence transmissible. Therefore, it is not surprising that LT-I production has been associated with enterotoxigenic E. coli (ETEC) carrying different adhesion factors or belonging to different serogroups (Willshaw et al., 1980; Nagy et al., 1999; Do et al., 2006; Khac et al., 2006). As a consequence, counteracting specific E. coli might not be the straightest way toward an inhibition of LT activity in the intestine. Hence, this article focuses on the inhibition of LT-I binding to GM1 to prevent the toxin from causing cell damage. Svennerholm and Holmgren (1978) were the first who described a GM1-enzyme-linked immunosorbent assay (ELISA) procedure for the identification of E. coli LT-I. More recently, Dawson (2005) gave an overview of different adaptations of this ELISA technique in his paper about a GM1-ELISA assay for CT. Based on Dawson's revised concept, we designed our GM1-ELISA assay for LT-I.
On a molecular level, Merritt et al. (1994a) compared the binding mode of GM1 for LT-I and CT with a two-fingered grip. GM1 puts forth a galactose residue as tip of a disaccharide forefinger, and a sialic acid residue as thumb to grasp the toxins (Merritt et al., 1994b; Minke et al., 1999). Of the two terminal sugar residues, the galactose gets more deeply buried into the toxin and is crucial for the toxin–GM1 binding. Hence, the galactose-binding site of LT-I was supposed to offer great promise as a target for blocking receptor binding (Merritt et al., 1994b, 1997; Hol et al., 1995). In view of this potential Achilles' heel, the aim of this study was to determine the interfering efficiency of galactose-rich substances with LT-I and CT binding to GM1. We chose to test natural substances, for it is especially the low-priced inhibitors that might have prospects to reach those who need them most in the developing world or help protect pigs against diarrhea.
Materials and Methods
Toxins and primary antibodies
Purified type I human E. coli heat-labile enterotoxin (LTh-I) was derived from the strain HC22/TP235km (Dertzbaugh and Cox, 1998) and was purchased from Sigma-Aldrich Chemie B.V. (E-8015; Zwijndrecht, The Netherlands). Purified porcine E. coli heat-labile enterotoxin (LTp-I) was derived from strain C600/P307 (Neill et al., 1983) and was acquired from Lactive B.V. (Lelystad, The Netherlands). CT was obtained from Sigma-Aldrich Chemie B.V. (C8052).
Polyvinylidene difluoride/LT-ELISA
Disks with a diameter of 5.5 mm were punched out of a polyvinylidene difluoride (PVDF) membrane (Boehringer, Mannheim, Germany). The PVDF disks were immersed in methanol (106009; Merck Chemicals, Darmstadt, Germany) for a few seconds until the membranes became translucent, and then equilibrated in tris-buffered saline (TBS: 0.05 M Tris-base and 0.15 M NaCl, pH 7.5) until the disks had sunken into the buffer, employing Petri dishes at room temperature. Subsequently, the PVDF disks were submerged in 100 μL of 2 μg LTp-I/mL TBS or phosphate-buffered saline (PBS: 0.01 M phosphate buffer and 0.15 M NaCl, pH 7.2) or TBS/PBS containing 0.05% Tween 20 (7374; Mallinckrodt Baker B.V., Deventer, The Netherlands), respectively, at room temperature for 1 h. This LT-binding step and all further steps were performed in the wells of a microplate (762070; Greiner Bio-One B.V., Alphen a/d Rijn, The Netherlands) that had been precoated with 1% bovine serum albumin (BSA; A7906; Sigma-Aldrich Chemie B.V.) in TBS. Then, the disks were washed three times with 200 μL TBS/PBS or TBS/PBS with Tween 20 for 5 min each. Afterward, the PVDF membranes were blocked with 1% BSA in TBS/PBS or TBS/PBS with Tween 20. Next, the PVDF disks were incubated with primary antibodies (Table 1; 1:200–1000 diluted in TBS/PBS or TBS/PBS with Tween 20 with 1% BSA), or solely in TBS/PBS or TBS/PBS with Tween 20 with 1% BSA for the negative control, at RT for 1 h, followed by washing as described above. For recognition of unlabeled antibodies, secondary antibodies were added as totals of 100 μL of horseradish peroxidase–conjugated bovine anti-goat IgG (1:2000 diluted in TBS/PBS or TBS/PBS with 1% BSA and 0.05% Tween 20; 805-035-180; Jackson ImmunoResearch Europe Ltd., Newmarket, UK) or peroxidase-conjugated goat anti-mouse IgG (1:1000 diluted in TBS with 1% BSA; 115-035-062; Jackson ImmunoResearch Europe Ltd.), followed by incubation at RT for 1 h. After washing as described above, the disks were incubated with 100 μL diaminobenzidine solution (10 mg DAB per 20 mL TBS; D8001; Sigma-Aldrich Chemie B.V.) and 4 μL 30% H2O2 (108597; Merck Chemicals) until the brown color reaction was fully developed. The color reaction was stopped by washing with TBS. The end-point absorbance of the brown-colored PVDF membranes was measured at a wavelength of 460 nm in a spectrophotometric plate reader in comparison with an untreated control membrane (SpectraMax 340; Molecular Devices Ltd., Wokingham, UK).
LT, Escherichia coli heat-labile enterotoxin.
Polystyrene/LT-ELISA
The wells of a high-binding polystyrene (PS) microplate (762071; Greiner Bio-One B.V.) were coated with 100 μL of 2 μg LTp-I/mL TBS or PBS or TBS/PBS, respectively, at 4°C overnight. To remove nonbinding LTp-I, plates were washed three times with 200 μL PBS containing 0.05% Tween 20. Uncovered nonspecific binding sites were blocked by the addition of 200 μL PBS, containing 1% BSA and 0.05% Tween 20, during incubation at RT for 1 h. Subsequently, plates were emptied and washed again as described above. Then, 100 μL of the antibodies each (Table 1) in PBS containing 1% BSA and 0.05% Tween 20 was added to the wells. Plates were incubated at RT for 1 h, followed by washing as described above. For recognition of unlabeled antibodies, secondary antibodies were added and incubated as described above. After washing as described above, 100 μL of ready-to-use TMB solution (Ceditest® 3,3′,5,5′-Tetramethyl benzidine; Prionics Lelystad B.V., Lelystad, The Netherlands) was added to all wells, followed by incubation at RT for 10 min. The reaction was stopped by addition of 50 μL 1.8 N H2SO4 per well. The end-point absorbance of the solutions was measured at a wavelength of 450 nm in the SpectraMax 340 plate reader.
GM1/LT-ELISA
Flat-bottom wells of medium- and high-binding PS strip plates mounted in frame (Microlon® 200 [762070] and Microlon 600 [762071]; Greiner Bio-One B.V.) were coated with 100 μL of GM1 monosialoganglioside (G7641; Sigma-Aldrich Chemie B.V.) in PBS at 37°C overnight. To remove nonbinding GM1, plates were washed three times with 200 μL PBS containing 0.05% Tween 20. Uncovered nonspecific binding sites were blocked by the addition of 200 μL ELISA buffer (0.01 M phosphate buffer, 0.5 M NaCl, and 0.003 M KCl, pH 7.0), containing 1% BSA and 0.05% Tween 20, during incubation at 37°C for 1 h.
Subsequently, plates were emptied and washed again as described above. Then, 100 μL of LTp-I in ELISA buffer was added to the wells. ELISA buffer without toxin served as blanks. Plates were incubated on a platform shaker (Polymax; Heidolph Instruments, Schwabach, Germany) with 25 rpm at 37°C for 4 h, followed by washing as described above. Antibody detection of bound LTp-I was performed as described for the PS/LT-ELISA.
Interference of test substances with toxin binding to GM1
Flat-bottom wells of medium-binding PS strip plates mounted in frame (Microlon 200) were coated with 100 μL of 1 μg GM1/mL PBS at 37°C overnight. To remove nonbinding GM1, plates were washed three times with 200 μL PBS containing 0.05% Tween 20. Uncovered nonspecific binding sites were blocked as described for the PS-ELISA. Subsequently, plates were emptied and washed again as described above.
Three different protocols (A, B, and C) were followed. In protocol A, the toxin and the test substance were simultaneously incubated in a GM1-coated plate; in protocol B, the toxin was preincubated with the test substance before the transfer of the mixture into a GM1-coated plate; and in protocol C, the test substance was postincubated in a GM1 and toxin–treated plate to resolve bound toxin from GM1.
For protocol A, a total of 50 μL test substance solution (double concentrated in ELISA buffer containing 1% BSA and 0.05% Tween 20) and 50 μL toxin solution (double concentrated in the same buffer) were pipetted into the wells of the GM1-coated and blocked PS strip plate. Plates were incubated on a platform shaker (Polymax; Heidolph Instruments, Schwabach, Germany) with 25 rpm at 37°C for 4 h, followed by washing as described for the GM1/LT-ELISA. Antibody detection of bound LTp-I was performed with anti-LT Mab LT-4/PO (Table 1) as described for the PS/LT-ELISA, employing an incubation time with TMB solution of 40 min. Antibody detection of bound LTh-I was performed with Serum Polycl. Goat anti-LT (Table 1) as described for the PS/LT-ELISA. Antibody detection of bound CT was performed using polyclonal IgG Rabbit anti-CT/HRP (2060-0040; AbD Serotec, Oxford, UK) in PBS containing 1% BSA and 0.05% Tween 20. Plates were incubated and treated as described for the PS/LT-ELISA.
For protocol B, a total of 70 μL test substance solution was incubated with 70 μL toxin solution (see protocol A) for 4 h at 37°C in an inert PCR plate (AB-0600; Abgene, Epsom, UK). Then, 100 μL of the preincubated mixture was pipetted into the wells of the GM1-coated and blocked PS strip plate. The subsequent steps were performed as described in protocol A.
For protocol C, a total of 100 μL of 5 ng or 1 μg LTp-I, 5 ng LTh-I or 4 ng CT per mL ELISA buffer containing 1% BSA and 0.05% Tween 20 was pipetted into the wells of the GM1-coated and blocked PS strip plate. Plates were incubated on a platform shaker (Polymax; Heidolph Instruments) with 25 rpm at 37°C for 4 h, followed by washing as described for the GM1/LT-ELISA. Then, 50 μL ELISA buffer containing 1% BSA and 0.05% Tween 20 and 50 μL test substance solution (see protocol A) were added to the wells. Plates were again incubated on the platform shaker with 25 rpm at 37°C for 4 h, followed by washing as described for the GM1/LT-ELISA. The subsequent steps were performed as described in protocol A.
The predominantly galactose-rich substances tested for their interfering effect with LTp-I binding to GM1 are shown in Table 2. Those substances that most efficiently counteracted LTp-I binding were also tested with LTh-I and CT.
See Becker and Galletti (2008) for more details.
CF, crude fiber; DM, dry matter.
Data analysis
The raw data obtained with the SpectraMax 340 plate reader were converted into inhibition percentages of toxin-binding to GM1 by means of sigmoidal calibration curves for different concentrations of the toxins using the SoftMax Pro 5.2 software of the reader. Results were considered valid if they fell within the 95% confidence interval of the calibration curve, and the intra-assay variation coefficient was <10%. The inhibition percentages were subjected to a two-way analysis of variance using the GenStat 11 statistical software package (VSN International Ltd., Hemel Hempstead, UK).
Results
Antibody recognition of LTp-I
A dot blot PVDF/LT-ELISA was performed to check for recognition of LTp-I by various primary antibodies (Tables 1 and 3). The employment of Tween 20 in the LTp-I solution ensured an effectual reduction of the background activity for subsequent LT detection. TBS buffer with Tween 20 performed slightly better than PBS buffer with Tween 20 for LTp-I, except with the Mab Mouse BG12 antibody. All antibodies tested were shown to react with LTp-I in this assay (Table 3). In the PS/LT-ELISA, the employment of TBS buffer with Tween 20 gave the best results for LTp-I. In the GM1/LT-ELISA, PBS with Tween 20 as solvent for LTp-I delivered the best results.
PVDF, polyvinylidene difluoride; LT, Escherichia coli heat-labile enterotoxin; ELISA, enzyme-linked immunosorbent assay; PS, polystyrene.
Compared to the PVDF/LT-ELISA, the PS/LT-ELISA and the GM1/LT-ELISA resulted in a lower detection incidence of LTp-I (Table 3).
GM1 concentration dependency of LTp-I binding
Figures 1 and 2 show the GM1 concentration dependency of LTp-I binding for different microplate materials (Microlon 200 vs. Microlon 600). With medium-binding microplates (Microlon 200), more LTp-I was bound at lower GM1 concentrations than with high-binding plates, indicating higher capacities for LTp-I recognition by GM1 in medium-binding plates.
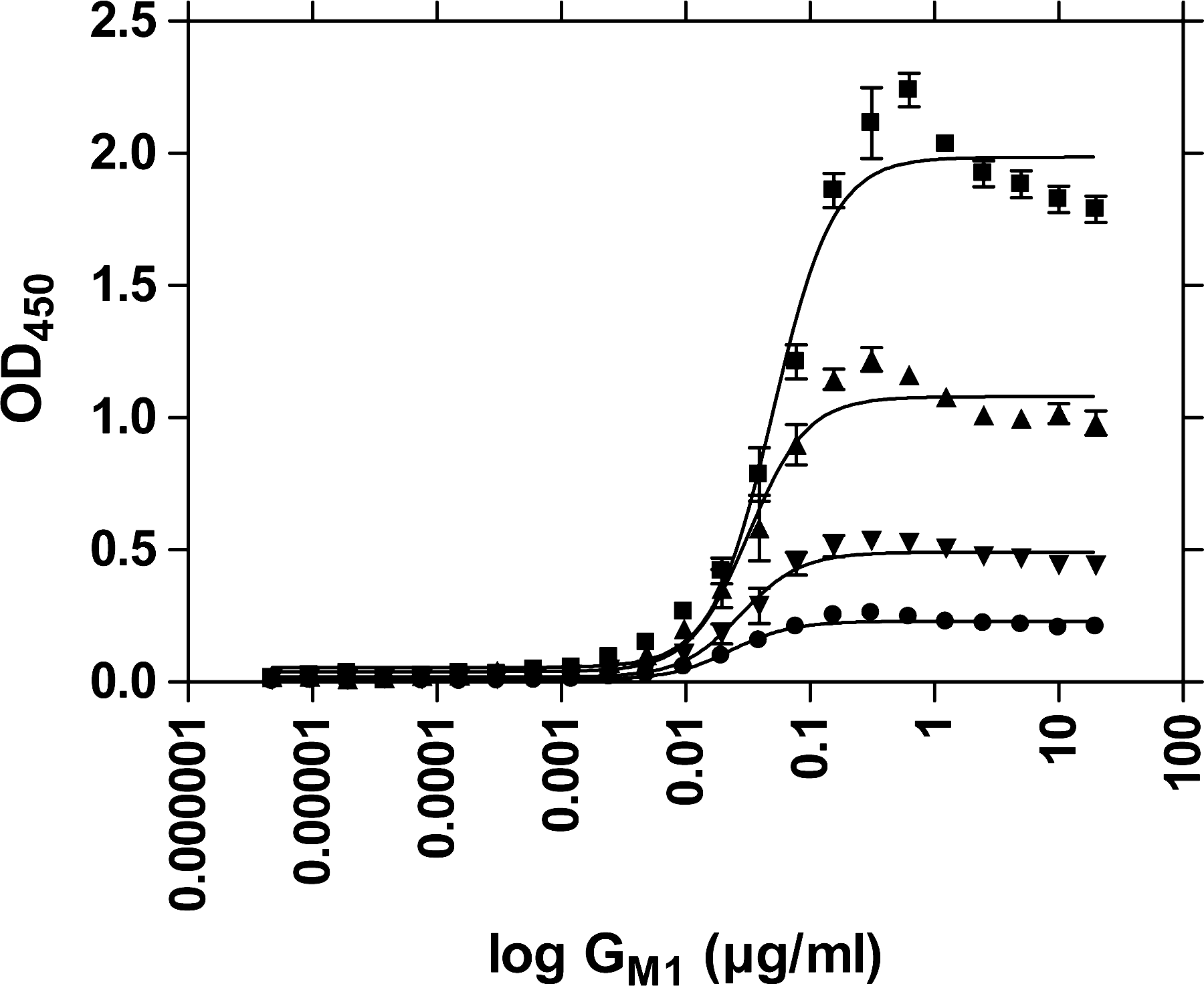
GM1 concentration dependency of LTp-I binding: ▪, 1 μg/mL LTp-I; ▴, 0.5 μg/mL LTp-I; ▾, 0.25 μg/mL LTp-I; ∙, 0.125 μg/mL LTp-I. Absorption results obtained after Mab LT-4/PO detection from six experiments using Microlon 200 microplates. Error bars indicate SEM.
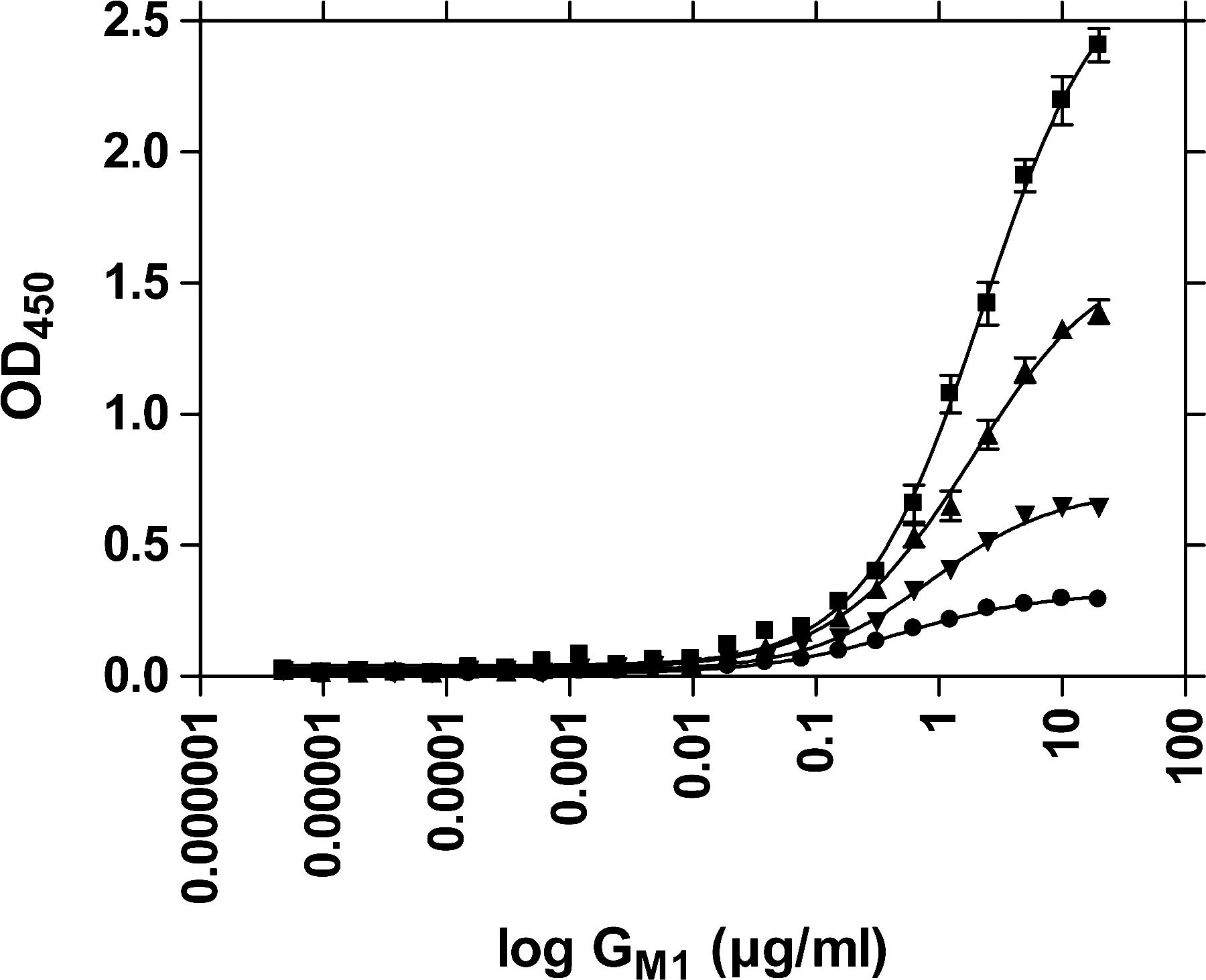
GM1 concentration dependency of LTp-I binding: ▪, 1 μg/mL LTp-I; ▴, 0.5 μg/mL LTp-I; ▾, 0.25 μg/mL LTp-I; ∙, 0.125 μg/mL LTp-I. Absorption results obtained after Mab LT-4/PO detection from six experiments using Microlon 600 microplates. Error bars indicate SEM.
Time dependency of LTp-I binding to GM1
Binding of LTp-I to GM1 was completed after about 4 h (Fig. 3).
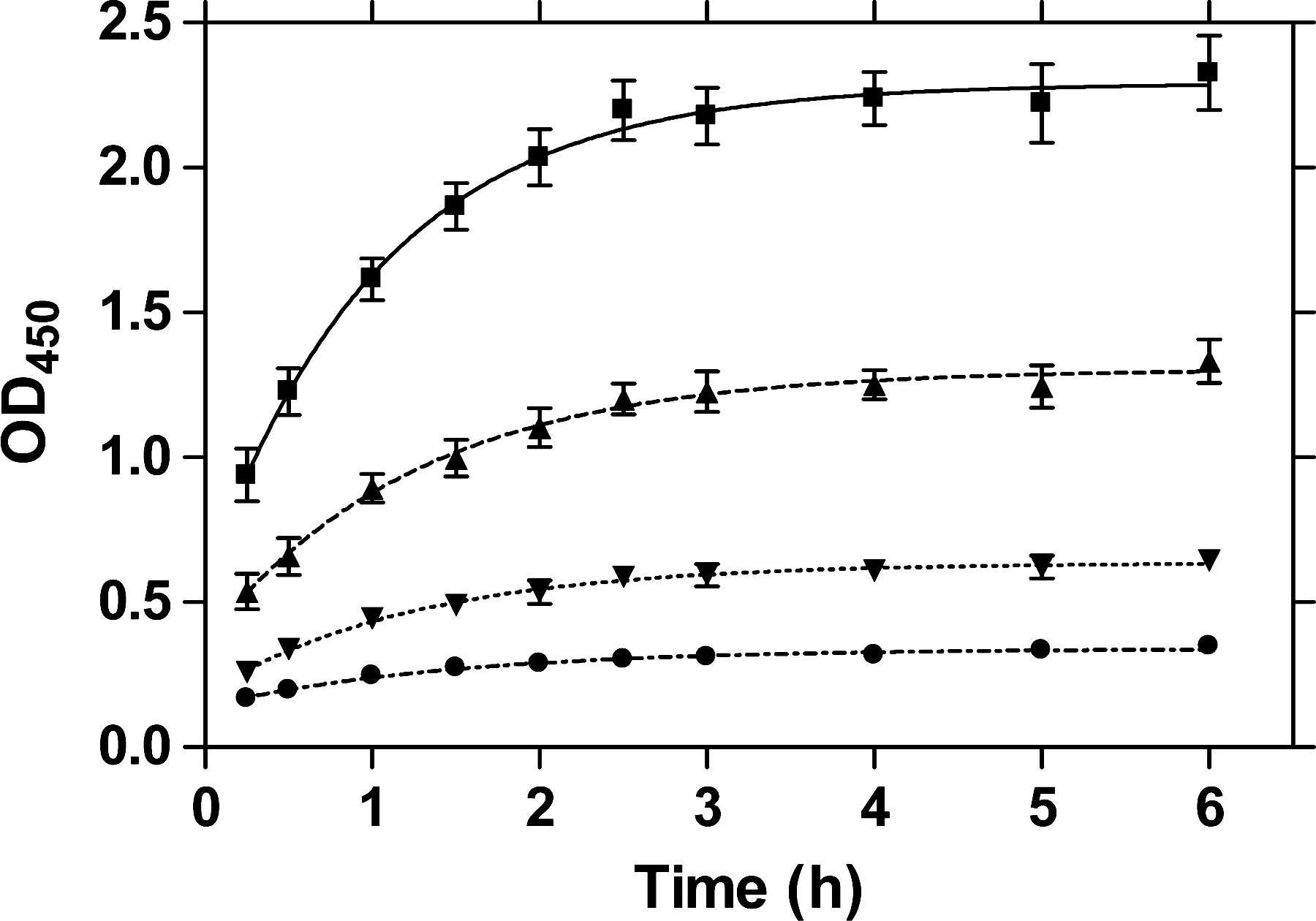
Time dependency of binding of LTp-I to 1 μg/mL GM1 coating: ▪, 1 μg/mL LTp-I; ▴, 0.5 μg/mL LTp-I; ▾, 0.25 μg/mL LTp-I; ∙, 0.125 μg/mL LTp-I. Absorption results obtained after Mab LT-4/PO detection from six experiments using Microlon 200 microplates. Error bars indicate SEM.
Interference of test substances with LT-I and CT binding to GM1
The interfering potential of different test substances with binding of LTp-I to GM1 was determined with the Mab LT-4/PO antibody (Table 4). At 1 μg/mL LTp-I, instantaneous mixing of toxin and substance sample (treatment A) resulted in comparable findings to those obtained by preincubation of the toxin with a substance sample (treatment B). The most efficient substance in terms of a reduction of toxin binding to GM1 was ground fenugreek seed. Skim milk powder, at a concentration of 5 mg/mL, also performed very well and did better than any of the individual milk saccharides lactose, lactulose, N-acetyl-D-galactosamine, 3′-sialyllactose, or 6′-sialyllactose. At 50 mg/mL, D-(+)-galactose and lactulose scored quite well. Further promising substances were tomato and willow wood. In contrast to the pretreatment and simultaneous treatment A and B, treatment C—after the toxin had already bound to GM1—was, in general, much less effective. At 50 mg/mL, in addition to fenugreek, larch and willow wood scored highest in treatment C. At 5 mg/mL, tara gum was the most potent one among all the substances tested in the after-treatment (Table 4).
LTp-I was determined with Mab LT-4/PO antibody.
Data are means of two independent experiments with duplicate measurements, except for data of substances marked by an asterisk (*), which are means of duplicate measurements without replication. Missing data in the table are due to costliness or solubility issues.
A, simultaneous incubation of LTp-I and test substance in GM1-coated plate; B, preincubation of toxin with test substance before transfer of mixture into GM1-coated plate; C, postincubation of test substance in GM1 and toxin–treated plate to resolve bound LTp-I from GM1.
When testing concentrations of 5 ng/mL LTh-I and 4 ng/mL CT, fenugreek I most efficiently blocked GM1-binding not only of LTh-I (Table 5) but also of CT (Table 6). Testing the effect of the substances lactose, lactulose, skim milk powder, tara gum, and fenugreek I on binding of 5 ng/mL LTp-I to GM1 with the Serum Polyclonal Goat anti-LT antibody resulted in comparable findings to the ones reported in Table 4 (data not shown). However, because these measurements were close to the detection limit of the assay, they were not as reliable as the ones shown in Tables 4 –6. With the ng-per-mL toxin concentrations (Tables 5 and 6), preincubation with fenugreek I resulted in slightly higher inhibition percentages than simultaneous incubation for LTh-I and CT.
LTh-I was determined with serum polyclonal goat anti-LT antibody.
Data are means of two independent experiments with duplicate measurements.
CT was determined with polyclonal IgG rabbit anti-CT/HRP.
Data are means of two independent experiments with duplicate measurements.
CT, cholera toxin; A, simultaneous incubation of CT and test substance in GMl-coated plate; B, preincubation of toxin with test substance before transfer of mixture into GM1-coated plate; C, postincubation of test substance in GM1 and toxin–treated plate to resolve bound CT from GM1.
Discussion
ELISA assays
While all primary antibodies were shown to recognize LTp-I when attached to the hydrophobic PDVF membrane material, significantly less did so when LTp-I was bound to the hydrophilic, high-binding PS material or to GM1 (Table 3). Probably, most antibodies with a specificity to the B-subunit of the toxin gave no results in the PS/LT- and the GM1/LT-ELISA, because LTp-I attached with the B-subunit to the PS plate material and to GM1, thus turning distinct B-sites inaccessible for antibody recognition.
When comparing high-binding with medium-binding microplate materials for coating with GM1, the medium-binding Microlon 200 proved superior (Figs. 1 and 2). To be able to expose its hydrophilic LT-I receptor, GM1 needs to attach to the carrier material with its hydrophobic ceramide moiety. Obviously, Microlon 200 offers more hydrophobic and thus more adequate binding conditions for that purpose than Microlon 600. Conversely, the higher hydrophilicity of Microlon 600 compared to Microlon 200 seems to cause a higher proportion of GM1 to bind upside down, resulting in the effect that on Microlon 600, bound GM1 is less reactive toward LT-I (Figs. 1 and 2).
The time dependency of LTp-I binding to GM1 (Fig. 3) was in agreement with the finding reported by Dawson (2005) for CT. Hence, the GM1-ELISA protocol adapted for LTp-I was also used for testing LTh-I and CT binding, with the exception that different antibodies were employed for toxin detection based on our own findings in terms of the antibodies' sensitivities or the supplier's recommendation in the case of CT.
Interference of test substances with LT-I and CT binding to GM1
Because the galactose binding site of LT-I was reported to be a promising target for blocking GM1 receptor binding (Merritt et al., 1994b), we tested the intervening efficiency with toxin adherence of several galactose-containing substances (Table 2).
Ground fenugreek seed performed best among all substances for all toxins in treatments A and B (Tables 4 –6). Fenugreek seeds contain about 20% fenugreek gum, consisting of galactomannans (Brummer et al., 2003), which might be the active principle of fenugreek. The spice fenugreek (seeds of Trigonella foenum-graecum L.; Gal/Man = 0.8–0.9), soybean hulls (Glycine max L. Merr.; Gal/Man = 0.7), and food stabilizers such as guar gum (E412; Gal/Man = 0.6–0.7), locust bean gum (E410; Gal/Man = 0.3), and tara gum (E417; Gal/Man = 0.3) all contain galactomannans, which in the food stabilizers are the predominant components (Daas et al., 2000; Srivastava and Kapoor, 2005). Guar gum is extracted from the seed of the leguminous shrub Cyamopsis tetragonoloba L., locust or carob bean gum is obtained from the seeds of the carob tree (Ceratonia siliqua L.), and tara gum is the endosperm of the seeds of the tara tree (Caesalpinia spinosa [Molina] Kuntze). Other galactomannan sources than fenugreek performed inferior in our tests, which might be due to the plant-specific characteristics of the respective galactomannans. Tara gum might offer some potential to resolve bound LTp-I from GM1, but not LTh-I or CT (Tables 4 –6). Guar and locust bean gum were less efficient in this respect with LTp-I, despite having higher or similar ratios of galactose/mannose in their mannans. Konjac gum contains glucomannan as its main component and is obtained from the corm of the konjac plant (Amorphophallus konjac K. Koch). Konjac gum was included in this study for the purpose of comparison. In line with its composition, the results obtained with konjac gum were much poorer than the ones obtained with the other gums.
Agar (E406), derived from red seaweeds, consists of agarose ([4)-3,6-An-α-L-Galp-(1 → 3)-β-D-Galp-(1 → ] n ) and agaropectin. In agaropectin, some sugar units are replaced by 4,6-O-(1-carboxyethylidene)-D-galactopyranose or by methylated or sulfated units (Labropoulos et al., 2002). Agar inhibited LTp-I binding to a higher extent than agarose (Table 4). However, compared to some non-agar products, the agar-related substances performed mediocre at best.
Arabinogalactans are composed of β-D-(1 → 3)-galactan main chains, with side chains made up of D-galactose and L-arabinose residues. Not only larch wood contains arabinogalactan (Gal/Ara = 6), but also carrots, tomatoes, and herbs (D'Adamo, 1990). Willow wood as a reference substance, despite containing only minor amounts of galactose (Skrigan et al., 1967), performed similar to larch wood in the LTp-I inhibition test A (Table 4).
Among mammalian substances, especially human milk is rich in the disaccharide lactose (55–70 mg/mL; β-D-Gal-(1 → 4)-D-Glc) and galactose-containing oligosaccharides (Kunz et al., 2000; Boehm and Stahl, 2007). Bovine milk contains about 40–50 mg lactose/mL (Kunz et al., 2000). During thermal processing of milk, lactose isomerizes to lactulose (β-D-Gal-(1 → 4)-D-Fru). Crystal structures have been described for LT-galactose, LT-lactose, and LT-lactulose complexes (Sixma et al., 1992; Hol et al., 1995; Merritt et al., 1997; Minke et al., 1999). Hol et al. (1995) had already pointed out that the galactose moiety of the sugars seemed more important for saccharide binding by LT than the nature of a second β1-linked hexose. However, knowledge about the displacing capacity of natural galactose-rich or -related compounds for the GM1 pentasaccharide remained sparse. Griffiths and Critchley (1991) reported that binding of LT-I to brush borders was effectively inhibited by galactose and fucose (6-deoxy-D-galactose). In our GM1/LTp-I inhibition test, however, galactose performed much better than fucose (Table 4). Minke et al. (1999) reported inhibition percentages for LT-binding to GD1b of 22% for galactose (20 mM; 4 mg/mL) and lactose (20 mM; 7 mg/mL) and 54% for lactulose (20 mM; 7 mg/mL) in a coated plate test similar to the one employed by us. GD1b has the same galactose–forefinger structure as GM1, but an extra N-acetylneuraminic acid (NANA) linked to the glycerol moiety of the NANA thumb of the GM1 pentasaccharide. Our findings agree with the ones of Minke et al. (1999) in terms of the superior performance of lactulose in the GM1/LTp-I ELISA. Apart from that, galactose did slightly better than lactose (Table 4). In contrast to LT binding, CT binding is quite specific for GM1 (Hol et al., 1995). Dawson (2005) observed practically no inhibition of CT binding to GM1 when trying to displace bound CT or preincubate CT with galactose. Likewise, we found somewhat lower inhibition potentials of lactose and lactulose for 4 ng/mL CT binding than for 5 ng/mL LTh-I binding (Tables 5 and 6). Free saccharides other than lactose occur only in trace amounts in bovine milk (Kunz et al., 2000). Low-level saccharides are, for example, modified lactoses, such as 3′-sialyllactose ( = 3′-N-acetylneuraminyl-D-lactose) and 6′-sialyllactose ( = 6′-N-acetylneuraminyl-D-lactose) (Urashima et al., 2001). These sialyllactoses amount to about 0.03–0.06 mg/mL milk (Kunz et al., 2000) and were also employed as test compounds in this study. Due to their costliness, the latter saccharides were only tested in low concentrations, which did not result in a noticeable inhibition (Table 4). Probably the linkage of NANA to the galactose moiety in these saccharides obstructed toxin binding. Comparably, the N-acetyl-group in N-acetyl-D-galactosamine most likely impeded the reaction with LTp-I. Unexpectedly, skim milk powder at a concentration of 5 mg/mL performed better in simultaneous (A) and in preincubation (B) treatments with 5 ng and 1 μg/mL LTp-I and 5 ng/mL LTh-I than all the single milk compounds tested, plus the galactose polymer galactan from gum arabic (Table 4). With 4 ng/mL CT, skim milk powder also did very well, especially when preincubated (treatment B) (Table 6). In skim milk, apart from saccharides, glycopeptides and glycolipids could be responsible for the observed effect (Otnæss et al., 1983; Shida et al., 1994).
Footnotes
Acknowledgments
This study was funded by the Knowledge Basis Research program of the Dutch Ministry of Agriculture, Nature, and Food Quality and by the project SAFEWASTES (no. 513949) of the European Commission FP6 Framework Programme. The authors are grateful to Dr. F.G. van Zijderveld (CVI, Lelystad, The Netherlands) for his kind provision of the B-specific antibody Mu-MoAb-α-LT4-HRPO, and to Monique Verveer for excellent technical assistance.
Disclosure Statement
No competing financial interests exist.