
Abstract
Application of high hydrostatic pressure has been proposed as an alternative quarantine process for the Mexican fruit fly, Anastrepha ludens Loew (Diptera: Tephritidae), one of the most important insects infesting mangoes, citrus fruits, and other fruits in Mexico and other Latin American countries. The present study was performed to determine the effect of high pressure treatments at 0°C on the survivorship of eggs and larvae of this pest. The effect of time and pressure level at near-freezing temperatures on the egg hatch and survival of larvae is discussed. Eggs and larvae were pressurized at 25, 50, 75, 100, or 150 MPa for 0, 5, 10, or 20 min at 0°C. Hatch was recorded for 1-, 2-, 3-, and 4-day-old pressurized eggs. For pressurized larvae (first, second, and third instars), percentage of survival was registered. Further, third instars were studied for their ability to pupate and develop to adulthood. The results showed that eggs had higher resistance than larvae to pressure. Larvae were not able to resist pressure treatments at 75 MPa for 20 min at 0°C. Treatments at 150 MPa for 20 min at 0°C were needed to destroy all eggs and larvae of A. ludens, indicating that this process might be useful as a quarantine method for infested fruits. However, more studies involving combination of high hydrostatic pressure at low temperature with precooling treatments are needed in order to decrease the pressure level to avoid fruit damage.
Introduction
Fruit fly metabolism is affected by cold temperatures, and several countries consider cold storage as an important disinfestation treatment. In Israel, citrus fruits are disinfested from the Mediterranean fruit fly, Ceratitis capitata (Wiedemann), by storing them for up to 16 days at 1.5°C (Gazit et al., 2004). USDA-APHIS has approved holding fruit at 1.1°C or less for 12 days as a cold treatment to be used against this species for 17 commodities from 49 countries (Phillips et al., 1997).
Cold storage has been used as a quarantine method since the beginning of the 20th century (Mangan and Hallman, 1998). Mediterranean fruit fly eggs and larvae do not survive when stored at 0–1.1°C or less for at least 12 days. However, a protective effect of fruits might influence the resistance of the first instars of Medfly larvae to cold storage at 1.1°C, requiring at least 16 days to reach the lethality required (the probit 9 standard for quarantine treatment efficacy is 99.9968% mortality tested in more than 100,000 insects) (Phillips et al., 1997). On the other hand, the most tolerant Medfly stage (third instar) did not survive after a 14-day cold treatment at 1.5°C (Lanza et al., 2005).
The Mexican fruit fly, Anastrepha ludens, is perhaps the most important insect infesting mangoes, citrus fruits, and other fruits in Mexico and other countries of Latin America. Cold treatment reduces the value of many tropical fruits due to chilling injury (Phillips et al., 1997; Gazit et al., 2004; Lanza et al., 2005), which imposes a practical limitation on selecting cold treatment as a quarantine method over other physical or chemical alternatives (Mangan and Hallman, 1998). Mangoes and other tropical fruits stored at temperatures near to 0°C suffer changes in their physiology, which alter their external and internal appearance as well as sensorial acceptation. These changes are highly dependent on the time of storage (Lederman et al., 1997).
High hydrostatic pressure processing (HPP) (300–600 MPa) is an alternative method to preserve the quality and stability of processed fruits (Otero et al., 2000; Boynton et al., 2002). HPP treatments have been reported as an alternative quarantine process for Mediterranean fruit fly, Mexican fruit fly, western cherry fruit fly, and codling moth (Butz and Tauscher, 1995; Neven et al., 2007; Candelario-Rodríguez et al., 2009). However, high pressure conditions reported as effective in destroying eggs and larvae of these pests caused unacceptable damage to the fruit. The use of combined treatments to meet quarantine requirements has been reviewed, and high or low temperature plus a chemical treatment (such as fumigation) or a physical treatment (such as radiation) have been proposed and tested (Mangan and Sharp, 1994). Reduced levels of pressure to destroy eggs and larvae are required, and the combination of up to 200 MPa pressure with cold temperature could be an alternative. Earlier research (Candelario-Rodríguez et al., 2009) has showed that eggs are the most resistant to treatments at ambient temperatures.
The objective of this work was to determine the combined effect of HPP applied at cold temperature (0°C) on the ability of the Mexican fruit fly eggs (the most resistant stage) to hatch and develop into larvae and pupae, with the expectation of reducing the stress on the fruit to levels below those causing damage.
Materials and Methods
The experimental work was conducted in the facility of Moscafrut at Metapa de Domínguez, Chiapas, México, with production of 220 million A. ludens weekly for sterile insect release programs. The eggs and larvae used in our study were supplied by this facility and reared as reported elsewhere (Candelario-Rodríguez et al., 2009).
High hydrostatic pressure system
Pressure treatments were carried out using a Cold Isostatic Press Model CIP42260 (Avure Autoclave Systems, Columbus, OH), with a high pressure chamber of 101.6 mm inner diameter and 584.2 mm length and an operating pressure capacity of 206 MPa. A mixture of crushed ice and anticorrosive lubricant in the ratio 5:1 (Hydrolubric 120-B; EF Houghton & Co., Valley Forge, PA) was used as the high pressure fluid (to minimize adiabatic heating), and the temperature of this mixture was adjusted at 0°C before high pressure treatment. The isostatic press equipment had no temperature control, and thus temperature varied with the time of pressurization, pressure level, and the nature of the samples introduced. Temperature of the high pressure fluid was measured after the treatment, and it was never higher than 5–6°C at the end of the treatments.
Eggs and larvae were pressurized at 25, 50, 75, 100, or 150 MPa for 0, 5, 10, or 20 min at 0°C. The time required to reach each final pressure was 8, 17, 44, 60, and 82 sec, respectively. The time period for release of pressure was always less than 30 sec.
High hydrostatic pressure treatment on eggs
Eggs 1, 2, 3, or 4 days old were processed independently. An aliquot of an egg/water solution containing 600 eggs/mL was used to completely fill Eppendorf® tubes, closing them immediately to avoid the presence of air bubbles. The tubes were held at 26 ± 1°C before the pressure treatment. The isostatic pressurization was in the range of 25–150 MPa for 0–20 min at 0°C. After treatments, a brush was used to separate the treated eggs from the Eppendorf® tubes. Treated eggs were separated into three rows of 100 eggs in Petri dishes using a stereoscope and incubated at 26 ± 1°C for 7 days to record the egg hatch.
High hydrostatic pressure treatment on larvae
The high pressure treatments on larvae used 25–150 MPa for 0–20 min at 0°C. The treatments were applied to first, second, and third instars, when they were 2, 5, and 8 days old, respectively. Approximately 1200 larvae from each instar plus 300 g of diet were introduced into plastic pouches of 200 mL per instar. The pouches were hermetically closed immediately before the high pressure treatment. After treatment, the pouches were opened and larvae in diets were returned to a 1 L plastic container, dispersed smoothly, and incubated for 9 days at 26 ± 1°C. The living larvae were recorded and transferred to Petri dishes. Groups of 100 larvae were deposited in each one of three Petri dishes containing vermiculite to induce pupation. They were held at 20°C, 80% relative humidity in darkness for 48 h. After the holding period, the number of pupae formed was recorded. The survivorship to emergence or reproductive capacity (the ability to have offspring) of the flies was not recorded.
Results
Effect of high pressure on eggs
Hatching rates for eggs of various ages at different pressurization levels and times of pressurization are given in Table 1. The eggs aged 1 to 4 days showed a high hatching rate (84.3–95.3%) when pressurized at 25 MPa for 0 min (pressurized and shifted immediately). The hatching rate was in the range 78.7–91.7% after 20 min of pressurization. For 1-, 2-, and 4-day-old eggs the hatching rate decreased with time. However, this decrease was not observed for 3-day-old eggs. Overall, eggs 4 days old were the most resistant to this level of pressure. The hatching rate of 1- to 4-day-old eggs pressurized at 50 MPa decreased from 81.0–92.7% when pressurized for 0 min to 22.0–82.7% after 20 min. Two-day-old eggs were the most resistant to this level of pressurization. At 75 MPa, the percentage of hatched eggs decreased from 54.7–86.7% at 0 min of processing to 1.3–45.7% after 20 min. Eggs 3 and 4 days old were the most resistant to this level of pressurization.
Percentage of surviving eggs denote mean values and standard deviation of three replicates.
The percentage of hatched eggs decreased at pressure levels of 100 and 150 MPa. At 100 MPa the percentage of hatching decreased from 1.3–61.3% at 0 min to 0.0–11.0% after 20 min. Eggs 1 and 3 days old were the most resistant to this level of pressurization. High pressure of 150 MPa reduced by almost 20% the ability of eggs to hatch just by reaching the pressure level (0 min). After 20 min at 150 MPa, all eggs were totally damaged, losing their ability to hatch.
Effect of high pressure on larvae
Larval survival at various developmental stages, pressure levels, and treatment times are given in Table 2. The percentage of larvae surviving the high pressure treatment of 25 MPa ranged from 0.2–12.2% at 0 min to 0.2–2.5% after 20 min. Third instars were the most resistant to this level of pressurization. At 50 MPa, the rate of survival decreased from 0.3–4.2% at 0 min to 0–4.0% at 20 min. Third instars were again the most resistant. At pressure levels of 75 MPa and above, survival was almost completely inhibited at all treatment times. For 0 min, the rate of survival varied from 0% to 1.7%, but none of the larvae survived when pressurized for 20 min. At 100 MPa for 0 min, the rate of survival ranged from 0.3% to 1.0%, and no larvae survived after 20 min.
The percentage of surviving larvae show the result of just one analysis.
The lowest levels of larval survival were obtained at 150 MPa. Larvae pressurized for 0 min showed 0–0.4% survival, and none survived after 5 or 20 min at this pressure level. First and third instars did not survive after 10 min at this level of high pressure, but second instar showed 0.1% survival.
Effect on pupation of surviving larvae
High pressure at 25 MPa for 0 to 20 min had less effect on the pupation of first instar than of third instar. The pupating capacity of second instar was totally inhibited even at 0 min (Table 3).
The values for percentage of pupating show the result of just one analysis.
Pupating ability of larvae pressurized at 50 or 75 MPa showed a similar behavior as at 25 MPa. However, third instars lost their ability to pupate after being pressurized at 75 MPa for any length of time. At 100 MPa for 0 min, a few first instars preserved their ability to pupate. Surviving second and third instars for all treatment times lost their ability to pupate. At 150 MPa, only the second instars that survived the high pressure treatment for 10 min preserved their ability to pupate.
Discussion
The inhibitory effects of high pressure treatments at 0°C on the hatching ability of the Mexican fruit fly eggs and survivorship of larvae are discussed as compared to another study that used high pressure treatment at 25°C on the same insect (Candelario-Rodríguez et al., 2009).
Effect on eggs hatching
HPP treatments at 0°C more efficiently reduced the hatching of A. ludens eggs compared to a similar study by Candelario-Rodríguez et al. (2009). In that study, the egg hatching rate at 75 MPa ranged from 87.3% to 100% for the 4-day-old eggs of A. ludens pressurized at 25°C. At 150 MPa with the same conditions of time and temperature, hatching rate ranged from 86% to 2.3%. These results agree with those reported by Butz and Tauscher (1995), who found that eggs of Mediterranean fruit fly, a very similar pest, were less resistant to HPP treatments when pressurized at 0°C than when pressurized at 12.5°C to 40°C.
Eggs of different ages showed different resistance to pressure treatment as a function of the time and pressure used. Most of the eggs were irreversibly damaged at pressures of 100 and 150 MPa applied for 10 or 20 min. However, some were able to resist such treatments and preserve their ability to hatch.
All eggs received the same amount of pressure for the period of time that each pressure was held because HPP treatment is instantaneous and uniform throughout the closed system inside the working chamber (Téllez-Luis et al., 2001). Thus, the response obtained might have been influenced by a different composition and physiology in accordance with the developmental stage of the eggs. Resistance of some specimens of the same species to extreme cold or heat has also been reported (Denlinger and Lee, 1998; Delinger and Yocum, 1998). No conclusions relating to age of eggs and pressure resistance could be drawn from this study.
The resistance of eggs to high pressure treatments also varies among insect species. The ability of Mediterranean fruit fly eggs to hatch was inhibited by pressure treatment for 5 to 20 min at 50 to 100 MPa at 25°C or at 125 MPa for 5 min (Butz and Tauscher, 1995). These authors reported a decrease in hatching ability on increasing the temperature during pressurization to 40°C or on using high pressure at 0°C. Eggs of western fruit fly (Rhagoletis indifferens) did not hatch after being pressurized at 172.4 MPa for 1 min at 16–18°C, and codling moth eggs lost their ability to hatch with high pressure treatments above 200 MPa (Neven et al., 2007).
Effect on surviving larvae
Larvae were less resistant to HPP treatments than eggs, and were also dependent on time and level of pressure treatment. After being treated for 20 min at 25 MPa, the rate of larval survival varied from 0.2% to 2.5% and at 50 MPa from 0% to 4.0%, but none of the larvae were able to resist pressure treatment at 75, 100, or 150 MPa for 20 min.
The mortality of A. ludens larvae was higher when HPP treatments were conducted at 0°C compared with similar treatments conducted at 25°C (Candelario-Rodríguez et al., 2009). At least 0.2–0.5% of A. ludens larvae were able to survive and pupate after receiving HPP treatments of 150 MPa for 0–20 min at 25°C. In the present study, differences in resistance to HPP treatments were influenced by the larval growth stage, and resistance decreased in the following order: third, first, and second instars (Table 2). A similar resistance to HPP has been reported for western cherry fruit fly larvae (Neven et al., 2007). The resistance of larvae to HPP varies also among insect taxa; Western cherry fruit fly larvae died after high pressure treatment at 172.4 MPa for 1 min, but codling moth larvae required a pressure of 208.9 MPa for 1 min to reach total mortality (Neven et al., 2007).
As discussed for egg hatching, the higher resistance of some specimens to HPP treatments might be associated with their physiological properties and not due to their failure to reach the same pressure in some parts of the system, since HPP is instantaneous and uniform in the closed system (Téllez-Luis et al., 2001; Torres and Velazquez, 2005; Velazquez et al., 2005a, 2005b).
Effect of cold temperature and HPP on organisms
Low temperature affects the development of insects in different ways, including division of eggs into multiple embryos or altering the postembryonic development, which may produce individuals with a mixture of larval and adult features. Tolerance to cold temperatures varies, depending on the geographic region of habitat. Tropical flies show low resistance to low temperature. The tolerance to cold shock injury and tolerance to long-term chilling injury are two forms of cold tolerance that rely on distinct mechanisms and that seem to have high heritability (Denlinger and Lee, 1998).
Organisms vary in their response to HPP. These differences are also present among strains of the same species (Galazka et al., 1995). For example, the same protein (myosin) from beef and four fish species treated at 200 MPa for 20 min at 5°C showed different levels of denaturation (Ashie et al., 1999).
HPP affects organisms in different ways, including changes in cell morphology, changes in the permeability of the membrane cell associated with separation of the cell membrane from the cell wall, and contraction of the wall cell with formation of pores. Figure 1 shows the larval damage in our study. Dark regions, air bubbles, liquid accumulation, and separation of the body from the external membrane are clearly identified.
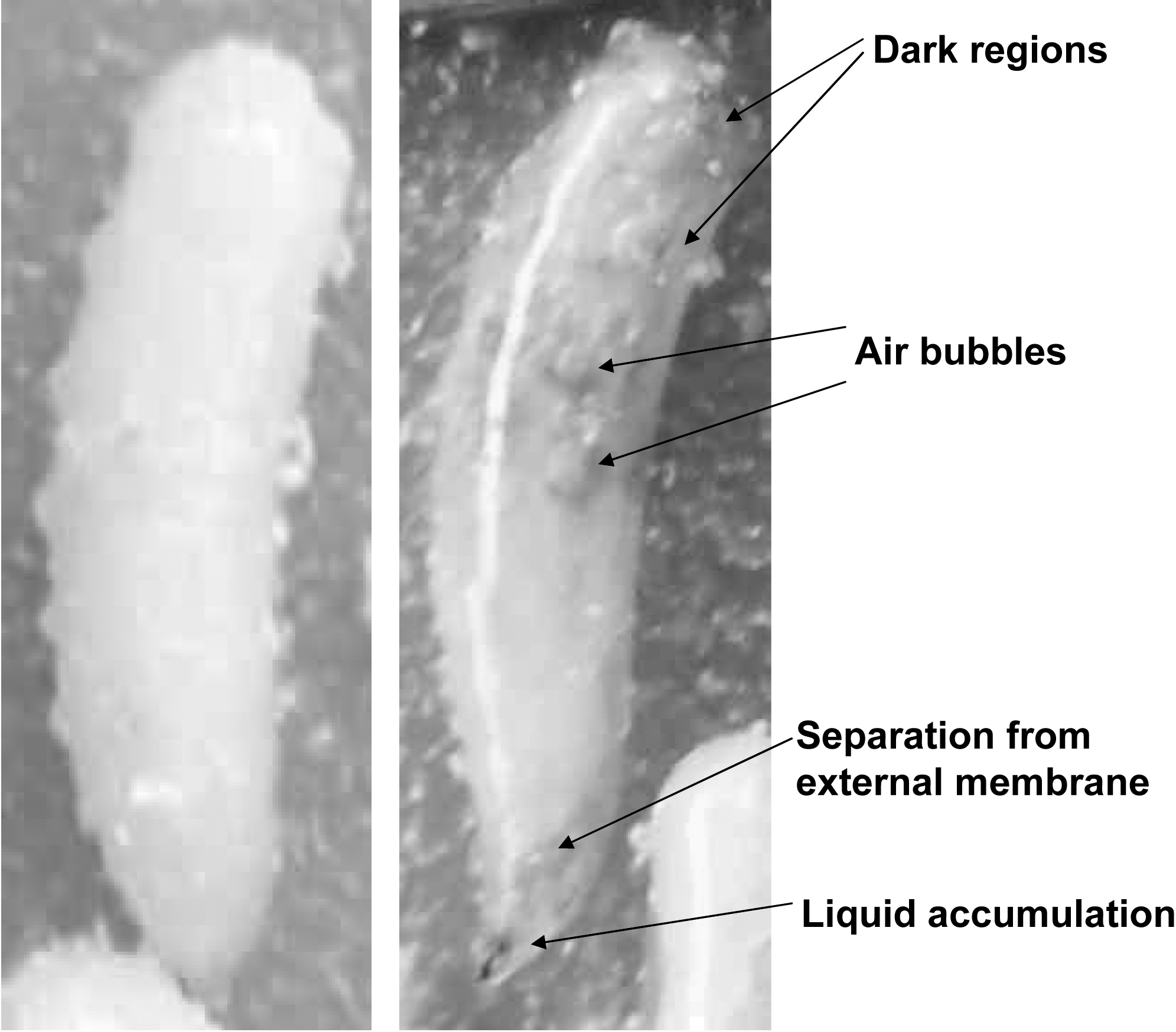
Anastrepha ludens third instar: (left) untreated control; (right) after pressurization at 150 MPa for 20 min at 0°C.
Denaturation of proteins is implied in the inhibition of the energy-producing reactions as well as in the modification of the essential enzymes involved in the growth and development of the cell, coagulation of cytoplasmic protein, and biochemical and genetic changes by inhibiting the enzymes involved in DNA replication and transcription (Farr, 1990; Pothakamury et al., 1995; Téllez-Luis et al., 2001). The adverse effect of HPP on egg hatching might be associated with a negative effect on the structure and permeability of external and internal membranes, a negative effect on proteins (denaturating/aggregation), or a combination of both. As larvae are multicellular organisms the adverse effect of HPP on hatching might result from damages to the cellular membranes as well as denaturation/aggregation of proteins, which affect their physiological functions. The physical damage to larvae by HPP treatments has been described recently (Candelario-Rodríguez et al., 2009) and completed in this study (Fig. 1).
Cold temperatures near to 0°C increased the lethality of HPP treatments for both eggs and larvae of A. ludens compared to similar treatments performed at 25°C (Candelario-Rodríguez et al., 2009). This increased lethality seems to be associated with cold shock effects from the low temperature near to the freezing point.
Protein denaturation/aggregation induced by high pressure depends on pressure level, pressure holding time, and temperature (Borderías et al., 1997; Fernández-Martín et al., 1998; Uresti et al., 2004). At lower temperatures (above freezing point) less damage of foods and lower protein denaturation/aggregation are expected (Dumay et al., 2006; Knorr et al., 2006; Neven et al., 2007). However, there is some evidence indicating that pressure and low/subzero temperatures have synergistic effects inactivating some microorganisms (Dumay et al., 2006).
The resistance of some specimens to HPP treatments at low temperatures might be explained as similar to the response of some specimens having tolerance to cold or heat shock injury and tolerance to long-term heating and chilling injury (Denlinger and Lee, 1998; Denlinger and Yocum, 1998).
Another factor that might be considered while comparing resistance of different organisms to high pressure treatments is the presence of solutes surrounding the proteins. Sugars and polyols at 8–12% have a stabilizing effect on proteins against high pressure (Ashie et al., 1999; Uresti et al., 2005).
Application on fruits
Fruits seem to have low tolerance to HPP treatments. Golden delicious apple was considerably affected by high pressure treatment at 96.6 MPa for 2 min. Sweet cherry was damaged by high pressure at 69 MPa (Neven et al., 2007). Ripe mangoes were instantaneously damaged by high pressure at 50 MPa for 0 min at 25°C during their storage at 20°C. But green mangoes showed little external changes when pressurized at 100 MPa for 20 min at 25°C, and no evident changes were observed under treatments lower than 75 MPa (Candelario-Rodríguez et al., 2009).
In our study, eggs and larvae of A. ludens were killed by pressure treatments of 150 MPa for 20 min at 0°C. On the other hand, green mangoes seem to resist high pressure treatments of 100 MPa for 20 min at 25°C, suggesting that HPP at cold temperatures can be considered as an alternative quarantine treatment. However, studies on the damage and ripening physiology of green mangoes pressurized in the range of 100–150 MPa at isothermal conditions (0°C) are needed to confirm this. Also, it should be determined if a precooling treatment for several hours or days at temperatures near to 0°C can increase the mortality rate of eggs and larvae by HPP without causing chilling injury to ripening of mangoes. The effects of combining high temperature with HPP to induce mortality of larvae and inhibit hatching of fruit fly eggs also require further study.
Conclusions
The efficacy of HPP treatments increased with treatment times over the range of pressure levels at 0°C. Eggs showed higher resistance than larvae to pressure. However, some eggs and larvae were able to resist even 150 MPa for 0–10 min, preserving the ability to hatch. Several larvae were able to resist and pupate even after being pressurized at 150 MPa, and a few larvae were able to pupate and develop adults after being pressurized. HPP treatments at 150 MPa for 20 min were able to inhibit the hatching of eggs and to kill the larvae of Mexican fruit fly. However, raw mangoes and other fruits are not able to support such treatment level. Therefore, HPP is a potential alternative quarantine treatment for the Mexican fruit fly, but more studies are needed.
Footnotes
Acknowledgments
The authors are grateful to Fondo Mixto CONACYT (The National Council on Science and Technology) and Tamaulipas state government for financially supporting this work (Project TAMPS-2003-C02-15) and to CONACYT for the scholarship granted to author Hugo Ernesto Candelario. We also thank José Manuel Gutiérrez Ruelas, M.Sc., Director of Fruit Flies, General Directorate of Plant Protection, Senasica, Sagarpa, and Dr. Pablo J. Montoya Gerardo, Assistant Director of Development Methods, Moscafrut Operational Program, for all the technical and scientific support.
Disclosure Statement
No competing financial interests exist.