
Abstract
The goal of this study was to characterize the starvation survival response (SSR) of a wild-type Listeria monocytogenes 10403S and an isogenic ΔsigB mutant strain during multiple-nutrient starvation conditions over 28 days. This study examined the effects of inhibitors of protein synthesis, the proton motive force, substrate level phosphorylation, and oxidative phosphorylation on the SSR of L. monocytogenes 10403S and a ΔsigB mutant during multiple-nutrient starvation. The effects of starvation buffer changes on viability were also examined. During multiple-nutrient starvation, both strains expressed a strong SSR, suggesting that L. monocytogenes possesses SigB-independent mechanism(s) for survival during multiple-nutrient starvation. Neither strain was able to express an SSR following starvation buffer changes, indicating that the nutrients/factors present in the starvation buffer could be a source of energy for cell maintenance and survival. Neither the wild-type nor the ΔsigB mutant strain was able to elicit an SSR when exposed to the protein synthesis inhibitor chloramphenicol within the first 4 h of starvation. However, both strains expressed an SSR when exposed to chloramphenicol after 6 h or more of starvation, suggesting that the majority of proteins required to elicit an effective SSR in L. monocytogenes are likely produced somewhere between 4 and 6 h of starvation. The varying SSRs of both strains to the different metabolic inhibitors under aerobic or anaerobic conditions suggested that (1) energy derived from the proton motive force is important for an effective SSR, (2) L. monocytogenes utilizes an anaerobic electron transport during multiple-nutrient starvation conditions, and (3) the glycolytic pathway is an important energy source during multiple-nutrient starvation when oxygen is available, and less important under anaerobic conditions. Collectively, the data suggest that the combination of energy-dependent internal adaptation mechanisms of cells and external nutrients/factors enables L. monocytogenes to express a strong SSR.
Introduction
I
Energy is the driving force of all ecosystems; therefore, the amount, quality, bioavailability, turnover, and replenishment of energy-rich molecules during starvation are important factors to be considered. Cells normally harness energy trapped within molecules from the environment to produce the ATP needed for cellular processes. In most cells, the energy required by cells is generated by either substrate level phosophorylation or oxidative phosphorylation (Moat et al., 2002). Lactic acid bacteria lack a cytochrome system and have been shown to produce energy via another method involving the proton motive force (PMF) (Thompson and Thomas, 1977; Poolman et al., 1991; Salema et al., 1996).
During starvation, with energy being limited, the SSR provides a mechanism by which certain bacterial species remain viable in the environment until the proper environmental conditions become available for growth. Therefore, the metabolic state of organisms during starvation is important in determining the mechanisms by which organisms survive.
An effective response to external stress such as starvation usually requires synthesis of proteins needed for cell maintenance (Helloin et al., 2003). The process of protein synthesis is energy dependent. Therefore, we hypothesized that to express an effective SSR, L. monocytogenes remains metabolically active and is thus able to produce the energy required for its protein synthesis and survival during starvation. Some light has been shed on the mechanisms and pathways used for survival during starvation by certain Gram-positive bacteria such as the lactics (Poolman et al., 1987; Redon et al., 2005; Ganesan et al., 2007). However, the mechanisms and pathways used by L. monocytogenes during growth and more importantly during starvation are yet to be fully elucidated.
Therefore, the objective of this study was to characterize the SSR of L. monocytogenes 10403S and a ΔsigB mutant strain during multiple-nutrient starvation by (1) examining the effects of starvation buffer changes on the viability of each strain, (2) investigating the effect of a known protein synthesis inhibitor (chloramphenicol), (3) determining the effects of inhibitors of substrate level phosphorylation (iodoacetate [IA], sodium fluoride [SF], and sodium arsenite), uncouplers of oxidative phosphorylation (sodium arsenate and potassium cyanide), or a PMF dissipater (2,4-dinitrophenol [DNP]) on the SSR of L. monocytogenes 10403S and a ΔsigB mutant, and (4) investigating the role of SigB in the SSR of L. monocytogenes. We also considered the effects of anaerobiosis versus aerobiosis on the activity of the metabolic inhibitors during starvation survival in L. monocytogenes 10403S and an isogenic ΔsigB mutant strain.
Materials and Methods
Preparation of bacterial strains
L. monocytogenes 10403S (wild type) and an isogenic ΔsigB mutant strain FSLA1-254 (a gift from Martin Wiedmann, Cornell University, Ithaca, NY) were used for this study. Stock cultures of both strains were kept frozen at −80°C in brain heart infusion (BHI) broth (TEKnova, Hollister, CA) with 20% glycerol until use. Bacterial strains from frozen stocks were each grown independently in BHI broth for 18 h and then subcultured in fresh BHI broth for 18 h at 37°C with shaking (200 rpm) before each experiment.
Survival of nutrient-deprived L. monocytogenes following buffer changes
Two-hundred milliliters of cultures of stationary-phase L. monocytogenes 10403S and the ΔsigB mutant, each containing an initial inoculum of about 9 log CFU/mL, were harvested by centrifugation (8000 rpm, 10 min), washed twice with equal volumes of phosphate-buffered saline (PBS; pH 7, 20 mM), and resuspended in equal volumes of PBS. PBS was used as the starvation buffer throughout this study. To change the starvation buffer, cells were harvested by centrifugation (8000 rpm, 10 min) and then resuspended in equal volumes of sterile PBS. Two sets of cells for each strain were prepared. One set of cells remained in their original starvation buffer for the duration of the study, and the other set of cells had their starvation buffer changed every 4 days for 28 days. Sampling tubes were incubated under static conditions at room temperature (23°C) for the duration of the study.
Survival of nutrient-deprived L. monocytogenes in the presence of the protein synthesis inhibitor (chloramphenicol)
Two hundred milliliters of cultures of stationary-phase cells of both strains containing about 9 log CFU/mL were prepared as described earlier with a few modifications. Cell pellets for each strain were washed and resuspended in 200 mL PBS with or without chloramphenicol (100 μg/mL), a known protein synthesis inhibitor. The minimum inhibitory concentration of chloramphenicol was 8 μg/mL for both strains. Chloramphenicol was added to L. monocytogenes cells immediately (0 h) or after starvation for 1, 2, 4, 6, 8, 24, and 48 h and incubated under static conditions at room temperature (23°C). Sampling was conducted on days 0, 2, 4, 7, 14, 21, and 28.
Preparation of aerobic and anaerobic media with or without inhibitors
Eighty milliliters of solution of aerobic PBS was prepared and autoclaved prior to use. Anaerobic conditions were created by the addition of 0.05% cysteine-HCl (Fisher Scientific, Pittsburgh, PA) with an indicator (0.001% resazurin; Difco, Detroit, MI), followed by boiling simultaneously with an argon gas flush. Eighty milliliters of aliquots of each solution were dispensed into anaerobic bottles (Bellco Glass, Vineland, NJ) that were sealed with butyl rubber septa and screw caps before autoclaving. Metabolic inhibitors were filter sterilized and added to sterilized aerobic or anaerobic PBS to obtain final concentrations of 0.1 M sodium arsenate, 0.01 M potassium cyanide, 0.01 M DNP, 0.001 M sodium IA, 0.1 M SF, or 0.01 M sodium arsenite. For the anaerobic study, metabolic inhibitors were dissolved in sterile water containing 0.05% cysteine-HCl and 0.001% resazurin and left in an anaerobic Coy Hood for at least 24 h. Inhibitors were aseptically added at appropriate concentrations to sterile anaerobic PBS in an anaerobic Coy Hood.
Preparation of aerobic nutrient-deprived cells
Eighty milliliters of overnight cultures of both strains containing about 9 log CFU/mL were harvested by centrifugation (8000 rpm, 10 min) and washed twice with PBS. Each strain was resuspended in 80 mL PBS with or without inhibitors and sampled on days 0, 4, 7, 14, 21, and 28. Sampling tubes were incubated under static conditions at room temperature (approximately 23°C) for the duration of the study.
Preparation of nutrient-deprived cells for anaerobic exposure
Eighty milliliters of overnight cultures of both strains containing about 9 log CFU/mL were harvested by centrifugation (8000 rpm, 10 min) and washed twice with PBS. The final PBS wash was conducted in anaerobic PBS containing 0.05% cysteine-HCl. The cell pellets were resuspended in the remaining liquid following supernatant removal (approximately 100–200 μL) and then transferred into anaerobic bottles containing anaerobic PBS with or without inhibitors using a sterile 23-G1 needle and syringe under an anaerobic Coy Hood to avoid reintroduction of oxygen into the medium. Sampling bottles were incubated under static conditions at room temperature for the duration of the study and sampling was conducted on days 0, 4, 7, 14, 21, and 28.
Bacterial enumeration
Each strain was enumerated separately and the surviving bacteria from each aerobic sampling tube were decimally diluted with PBS. Dilutions from tubes were surface spread plated in duplicate onto Listeria-selective agar (LSA) plates (Oxford formulation; EM Science, Merck KgaA, Gibbstown, NJ). Tests to determine cell injury in starved cells were conducted by spread plating cells onto both LSA and tryptic soy agar with 0.6% yeast extract (Becton, Dickinson and Company, Sparks, MD). Surviving bacteria from anaerobic bottles were decimally diluted with anaerobic PBS under an anaerobic Coy Hood and surface spread plated in duplicate onto anaerobic LSA plates. The plates were incubated aerobically or anaerobically at 37°C for 48 h before colonies were counted.
Analysis and interpretation of experimental data
The mean numbers of surviving cells of L. monocytogenes were reported as log CFU/mL values from three separate experiments, each performed in duplicate. The minimum detection limit was 10 CFU/mL. Analysis of variance was conducted to determine if the treatment mean values in log CFU/mL of nutrient-deprived L. monocytogenes 10403S and the ΔsigB mutant were affected by strain, metabolic inhibitor, gas phase incubation conditions, replication, or time. The means for each treatment sample were analyzed using the Proc GLM function of the SAS software (SAS Institute, Cary, NC). The experimental design model used was the split plot, with time as the split plot factor. The treatment means were judged to be statistically significant for the F-test if p ≤ 0.05.
Results and Discussion
SSR of L. monocytogenes 10403S and a ΔsigB mutant strain following starvation buffer changes
When wild-type L. monocytogenes and the ΔsigB mutant strain were left in their original starvation buffer, both strains expressed an SSR, with no significant differences in their survival (p > 0.05). This suggested that L. monocytogenes likely utilizes SigB-independent mechanisms to survive during multiple-nutrient starvation. In contrast, other studies (Ferreira et al., 2001; Herbert and Foster, 2001; Moorhead and Dykes, 2003) reported that SigB contributes to the viability of L. monocytogenes cells during glucose-limiting conditions. Significant differences (p ≤ 0.0001) were found in the survival of cells kept in their original starvation buffer versus cells whose starvation buffer was changed every 4 days (Fig. 1). Cells with starvation buffer changes were unable to express any SSR, and cell counts were decreased to less than 3 log CFU/mL by day 28. No significant differences were observed when the starved cells were plated on selective versus nonselective medium (p > 0.05), suggesting that nutrient-deprived cells are not critically injured. Therefore, cells of L. monocytogenes that have been previously starved do not suffer ill effects from starvation and grow readily once ideal nutrient growth conditions are available.
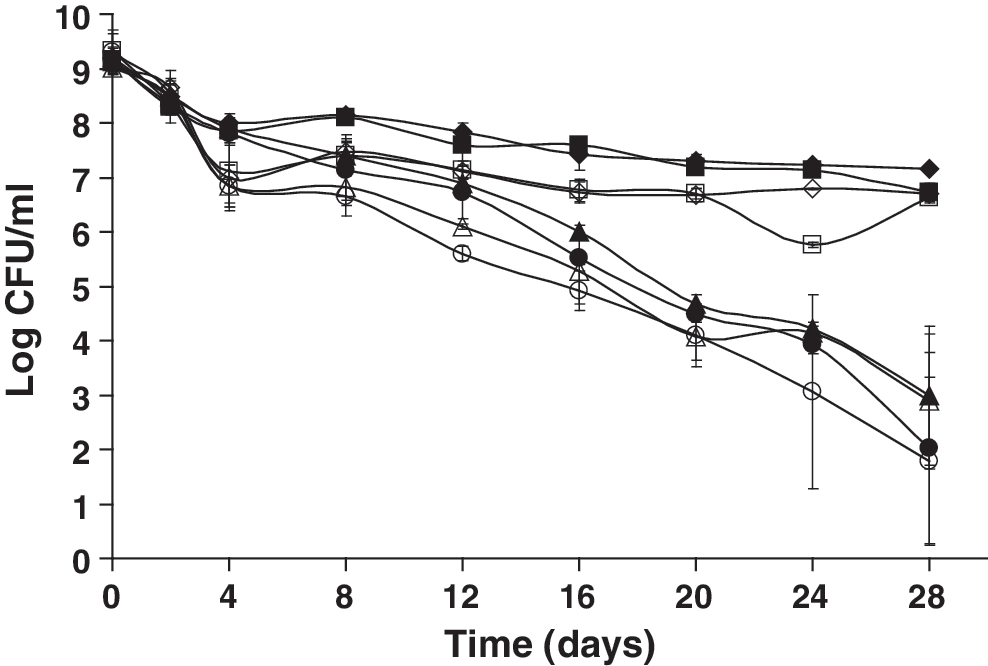
Survival of nutrient-deprived Listeria monocytogenes 10403S (closed symbols) and an isogenic ΔsigB mutant (open symbols) that were starved in the original starvation medium (phosphate-buffered saline) or had their starvation medium changed every 4 days, were held at 23°C, and then plated on tryptic soy agar with 0.6% yeast extract (TSAYE) or Listeria-selective agar (LSA) over 28 days. ♦, wt (NS) TSAYE; ▪, wt (NS) LSA; ▴, wt (CB) TSAYE; •, wt (CB) LSA; ◊, ΔsigB (NS) TSAYE; □, ΔsigB (NS) LSA; Δ, ΔsigB (CB) TSAYE; ○, ΔsigB (CB) LSA. NS, starvation buffer was not changed throughout the experiment; CB, starvation buffer was changed every 4 days; wt, wild type.
The protein contents in the cell pellets of both strains decreased over time for both strains irrespective of whether the cells remained in their original starvation buffer or not. However, there was an increase in supernatant nucleic acids and proteins which correlated with a sharp increase in dead cell debris within the first 8 days (data not shown). The initial increase in dead cells likely served as a source of nucleic acids and proteins. When the external buffer was changed, proteins and nucleic acids were washed off, and thus the remaining population was unable to establish itself or express an SSR. Previous studies have also suggested that lysis of dead cells during starvation may provide the remaining cell population with an available source of nutrients needed for energy production (Kolter et al., 1993; Watson et al., 1998).
Effect of chloramphenicol on the SSR of L. monocytogenes
The effects of chloramphenicol (100 (g/mL), a protein synthesis inhibitor, on the SSR of L. monocytogenes 10403S and the ΔsigB mutant strain were investigated (Fig. 2). When chloramphenicol was added within the first 4 h of starvation, neither strain could elicit an SSR. The data also showed that the ΔsigB mutant strain appeared to be more adversely affected than the wild type by the addition of chloramphenicol in the first 4 h of starvation (p ≤ 0.0022). Therefore, it is likely that in the absence of protein synthesis, the SigB-dependent proteins produced in the wild type prior to starvation allowed better survival compared with the ΔsigB mutant strain. In a study involving the Gram-positive bacterium Bacillus subtilis, Volker et al. (1999) observed that exhaustion of glucose or phosphate induced the SigB response but surprisingly SigB did not appear to be required for starvation survival in B. subtilis. Similarly, in L. monocytogenes it would seem that, as in the case with B. subtilis, the SigB-dependent response may be induced in the stationary phase prior to starvation but may not be required for the cell maintenance or cell viability during long-term nutrient starvation. In contrast, the wild-type or ΔsigB mutant strains were still able to express an SSR following the addition of chloramphenicol after 6 h of starvation. The addition of chloramphenicol after 6 h of starvation yielded colony counts that were significantly lower than the controls (p ≤ 0.0001), by about 1–2 log CFU/mL. It appears that cells starved for at least 6 h become stress hardened and have probably already synthesized the proteins necessary for an effective SSR. Collectively, these observations show the importance of protein synthesis during starvation and suggest that proteins necessary for an SSR are likely produced within the first 6 h of starvation. Other studies (Watson et al., 1998; Herbert and Foster, 2001) have also described the presence of starvation-associated protein synthesis in Gram-positive bacteria under carbon starvation conditions.

Survival of nutrient-deprived L. monocytogenes 10403S (closed symbols) and an isogenic ΔsigB mutant (open symbols) in phosphate-buffered saline following the addition of chloramphenicol at different times at 23°C over 28 days. ♦, wt control; ▪, wt 0 h; ▴, wt 2 h; •, wt 4 h; +, wt 6 h; ◊, ΔsigB control; □, ΔsigB 0 h; Δ, ΔsigB 2 h; ○, ΔsigB 4h; ×, ΔsigB 6 h.
Effect of a PMF dissipater on the SSR of L. monocytogenes
DNP uncouples energy-rich phosphate bonds and dissipates the H+ gradient and is thereby a known dissipater of the PMF (Loomis and Lipman, 1948; Shioi and Taylor, 1984). In the presence of 0.01 M DNP, neither the wild-type nor the ΔsigB mutant strain was able to express an SSR (Fig. 3). Further, the survival of both strains incubated anaerobically in the presence of DNP was reduced when compared with cells starved in an aerobic environment. The addition of DNP under aerobic or anaerobic conditions did not yield significant differences in the survival of the wild-type versus the ΔsigB mutant strain (p > 0.05), suggesting that the SigB-dependent response plays no essential role in the SSR of L. monocytogenes during PMF dissipation by DNP. Classically, during aerobic conditions, the membrane ATPase is in charge of the direction of proton influx and ATP synthesis, whereas during anaerobic conditions, it is responsible for the direction of proton expulsion via ATP hydrolysis (Becker et al., 2003). The faster death rates of the cells kept under anaerobic conditions may result from their defect of ATP when compared with those held aerobically. It appears that in L. monocytogenes, PMF-derived energy is likely necessary for effective cellular responses and cell maintenance during starvation under both aerobic and anaerobic conditions.

Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to the proton motive force dissipater 0.01 M 2,4-dinitrophenol (DNP) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt DNP + oxy; ▴, ΔsigB control +oxy; •, ΔsigB DNP + oxy; ◊, wt control − oxy; □, wt DNP −oxy; Δ, ΔsigB control − oxy; ○, ΔsigB DNP − oxy.
Effect of inhibitors of oxidative phosphorylation on the SSR of L. monocytogenes
In the presence of 0.1 M sodium arsenate (Fig. 4) under aerobic conditions, both the wild type and the mutant could express an SSR, with the mutant exhibiting much less survival. However, when treated with the inhibitor under anaerobic conditions, neither strain was able to express an SSR. Sodium arsenate, though not specific in its mode of action, has been shown to deplete intracellular ATP (Pillai, 1938). The depletion of intracellular ATP upon sodium arsenate addition coupled with less ATP production through anaerobic fermentation possibly explains why L. monocytogenes cells kept anaerobically appeared to be more adversely affected than those incubated aerobically. The more adverse effects of 0.1 M sodium arsenate on the ΔsigB mutant suggest that the SigB-dependent mechanisms activated during the stationary phase prior to starvation likely allowed the wild type to survive better than the mutant strain in an ATP-depleted environment.
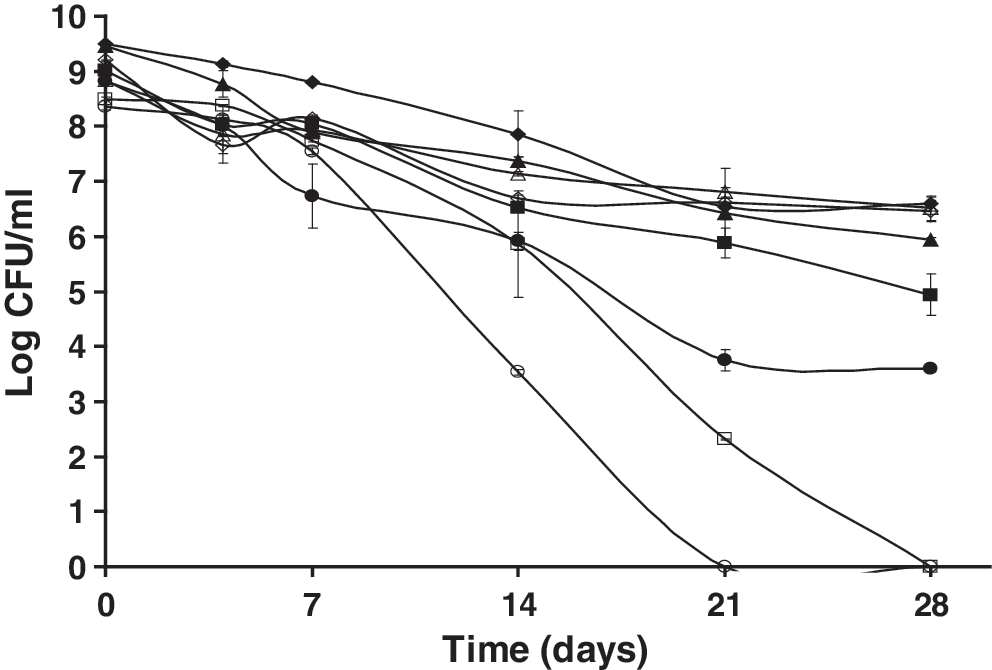
Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to 0.1 M sodium arsenate (ATP depletion) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt sodium arsenate + oxy; ▴, ΔsigB control + oxy; •, ΔsigB sodium arsenate + oxy; ◊, wt control − oxy; □, wt sodium arsenate − oxy; Δ, ΔsigB control − oxy; ○, ΔsigB sodium arsenate − oxy.
Potassium cyanide is a strong inhibitor of the electron transport in the aerobic cytochrome system (Cline, 1970). In the presence of molecular oxygen and 0.01 M potassium cyanide, both strains were unable to express an SSR (Fig. 5). However, the ΔsigB mutant strain appeared to be more adversely affected by the presence of potassium cyanide (p ≤ 0.0001) over 28 days. These data suggest a possible role for SigB in electron transport during multiple-nutrient starvation. Potassium cyanide appeared to have an adverse effect on the SSR of L. monocytogenes under anaerobic conditions as well, though less severe than under aerobic conditions. This observation provides evidence for anaerobic electron transport in L. monocytogenes.
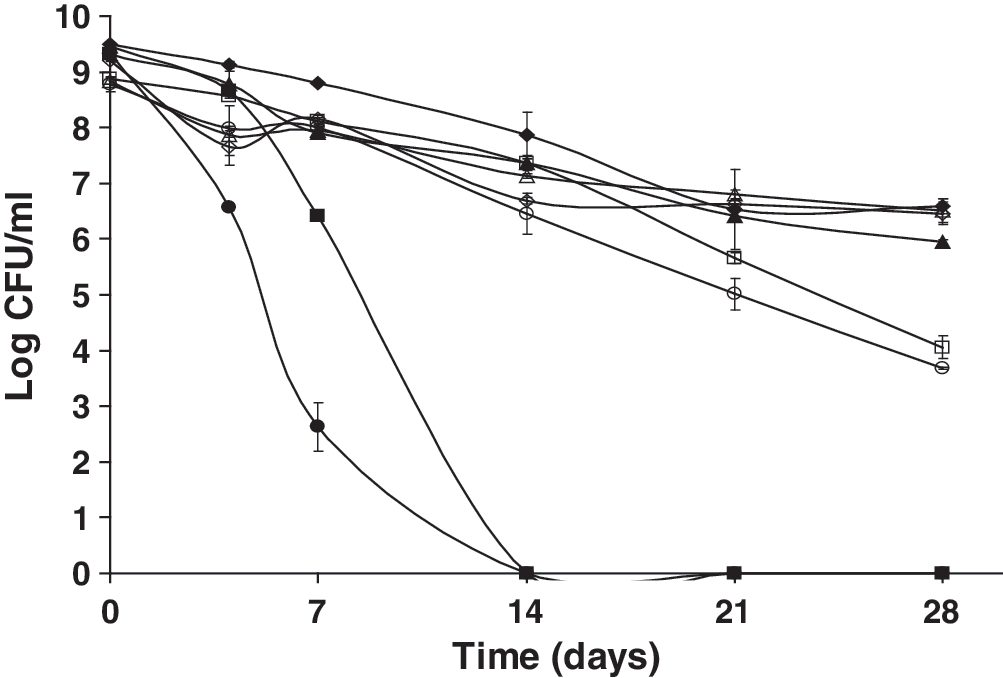
Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to 0.01 M potassium cyanide (inhibitor of cytochrome system) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt potassium cyanide + oxy; ▴, ΔsigB control +oxy; •, ΔsigB potassium cyanide + oxy; ◊, wt control − oxy; □, wt potassium cyanide − oxy; Δ, ΔsigB control − oxy; ○, ΔsigB potassium cyanide − oxy.
Effect of inhibitors of substrate level phosphorylation on the SSR of L. monocytogenes
The results (Fig. 6) showed that there was a significant effect following treatment of starved cells with IA only under aerobic conditions (p ≤ 0.0001) over 28 days. IA specifically inhibits the glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase (GPDH) (Even et al., 1999), and our data suggest that in the presence of molecular oxygen, the reaction catalyzed by GPDH is important for cell maintenance during starvation in L. monocytogenes. Similarly, Poolman et al. (1987) suggested that GPDH had almost full control (90%) over the glycolytic flux in nongrowing Lactococcus lactis cells. In contrast, under anaerobic conditions, cells were able to express a strong SSR in the presence of IA. This suggests that this pathogen likely utilizes a pathway that is not dependent on the glycolytic step catalyzed by GPDH to obtain energy for cell maintenance during starvation under anaerobic conditions. No significant differences were found in the SSR between the wild-type and the ΔsigB mutant strain (p > 0.05) in the presence of IA.

Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to 0.001 M sodium iodoacetate (IA) (inhibitor of the glycolytic enzyme glyceraldehyde-3-phosphate dehydrogenase) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt IA + oxy; ▴, ΔsigB control + oxy; •, ΔsigB IA + oxy; ◊, wt control − oxy; □, wt IA − oxy; Δ, ΔsigB control − oxy; ○, ΔsigB IA − oxy.
The effect of 0.1 M SF (Fig. 7), a known inhibitor of the glycolytic enzyme enolase (Kanapka and Hamilton, 1971; Wang and Himoe, 1974), was also assessed. Neither strain was able to express an SSR in the presence of SF under aerobic incubation. However, when incubated anaerobically, only the wild type was able to elicit a strong SSR. These findings indicate that starved L. monocytogenes likely utilizes the reaction involving enolase during aerobic but not anaerobic metabolism. However, the inability of the ΔsigB mutant strain to elicit an SSR under anaerobic conditions does suggest a possible role for SigB when this pathogen encounters energy stress provided by inhibiting enolase in the absence of oxygen.
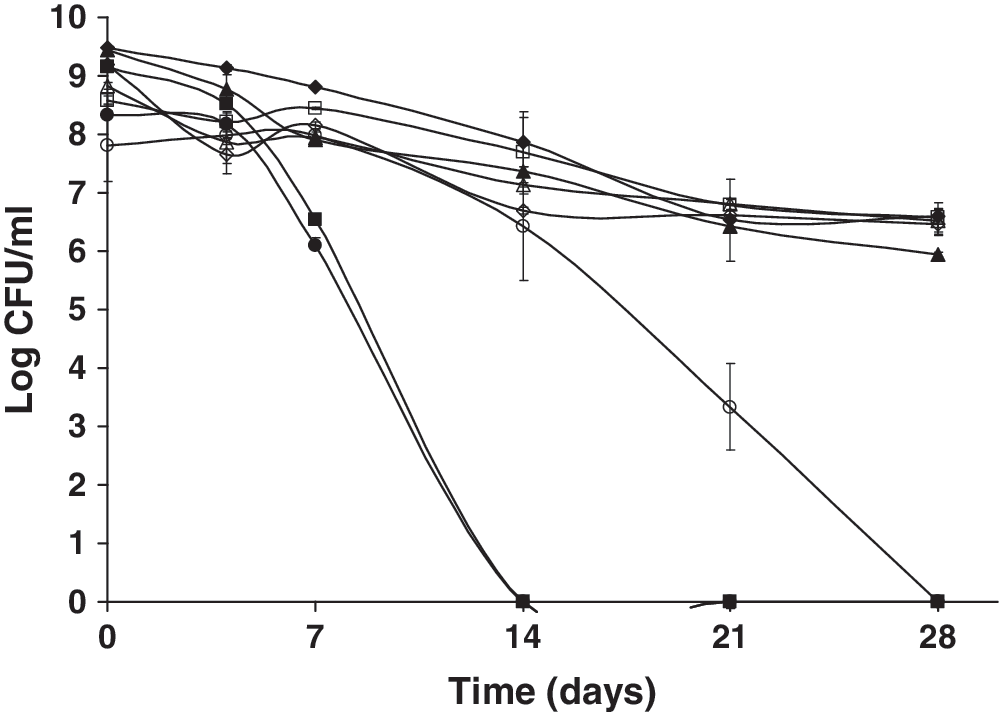
Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to 0.01 M sodium fluoride (SF) (inhibitor of the glycolytic enzyme enolase) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt SF + oxy; ▴, ΔsigB control + oxy; •, ΔsigB SF + oxy; ◊, wt control − oxy; □, wt SF − oxy; Δ, ΔsigB control − oxy; ○, ΔsigB SF − oxy.
The effect of sodium arsenite (Fig. 8), a known inhibitor of pyruvate dehydrogenase (Peters et al., 1946), was also assessed. Both the mutant and wild-type strains were unable to elicit an SSR under aerobic incubation. Under anaerobic conditions, the wild-type, unlike the mutant strain, was able to elicit a strong SSR in the presence of sodium arsenite. Johnson and Collins (1973) observed that under anaerobic conditions, Streptococcus faecalis appeared to use the pyruvate dehydrogenase enzyme complex in glucose metabolism. Conversely, the results of our study show that starved L. monocytogenes likely utilizes the pyruvate dehydrogenase enzyme complex during aerobic but not anaerobic metabolism.
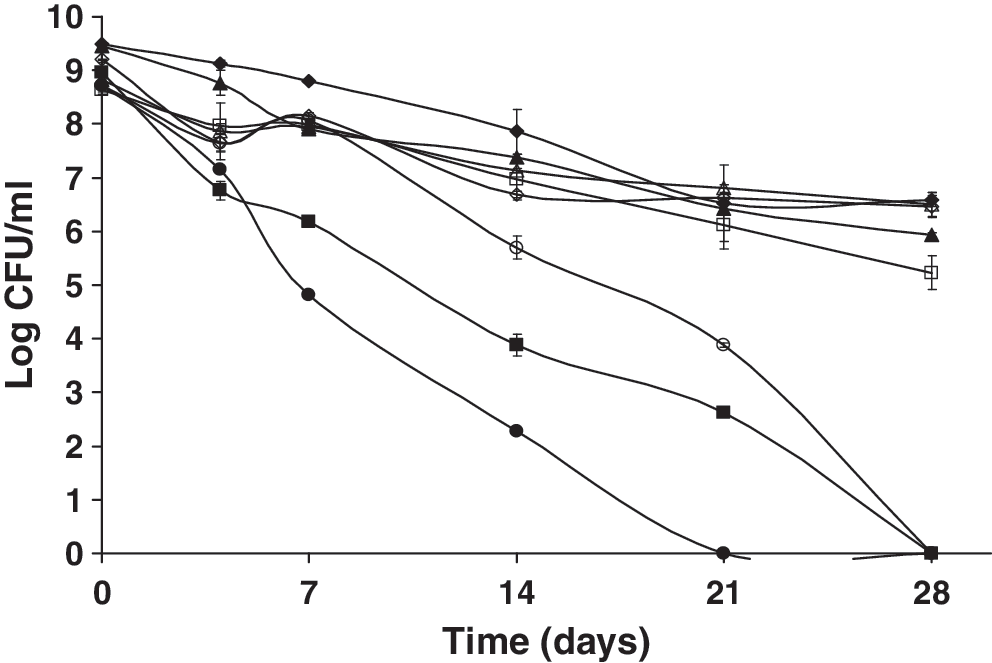
Survival of nutrient-deprived cells of L. monocytogenes 10403S and an isogenic ΔsigB mutant kept at 23°C following exposure to 0.01 M sodium arsenite (inhibitor of the glycolytic enzyme pyruvate dehydrogenase) in the presence (closed symbols) or absence of oxygen (open symbols) over 28 days. ♦, wt control + oxy; ▪, wt sodium arsenite + oxy; ▴, ΔsigB control + oxy; •, ΔsigB sodium arsenite + oxy; ◊, wt control − oxy; □, wt sodium arsenite −oxy; Δ, ΔsigB control − oxy; ○, ΔsigB sodium arsenite − oxy.
Conclusions
The goal of this study was to characterize the SSR of L. monocytogenes, an important foodborne pathogen. It is essential that we understand the survival mechanisms of this pathogen to devise effective intervention strategies. To express an efficient SSR, L. monocytogenes appears to require protein synthesis, metabolic activity, and the presence of nutrients or factors from dead cell debris that are present in the external starvation medium. It is probable that endogenous and exogenous energy sources from stored substrates and dead cell material, respectively, are converted into usable energy to drive the SSR during starvation. This study showed that energy metabolism and specifically glycolysis is taking place during multiple-nutrient starvation of L. monocytogenes under aerobic conditions but not necessarily required under anaerobic conditions. Further, the study suggests that energy derived from the PMF is important during starvation under both aerobic and anaerobic conditions in L. monocytogenes. The adverse effect of potassium cyanide under anaerobic conditions is significant, as this provides evidence for the presence of anaerobic electron transport during multiple-nutrient starvation. Other findings in this study indicate that SigB-dependent mechanisms activated during the stationary phase prior to starvation may enable L. monocytogenes to survive better when faced with additional stresses due to energy depletion. Our findings so far are representative of a single temperature, single inoculum, and a single strain with its isogenic mutant. This study characterized the response of L. monocytogenes under temperatures representative of food processing conditions. However, as L. monocytogenes is ubiquitous in the environment, more studies are needed to investigate the response of this pathogen at different temperatures and different inoculum levels. In addition, future studies to further elucidate the SSR of this pathogen would require testing a collection of different L. monocytogenes strains comprising of different serotypes from different origins. Overall, a combination of both external nutrients/factors from lysed dead cells and internal adaptation mechanisms of cells upon metabolic activity and protein synthesis enable L. monocytogenes to express a strong SSR.
Footnotes
Acknowledgments
This study was supported in part by a grant from the U.S. Department of Agriculture-Food Safety Consortium. The authors thank Dr. Martin Wiedmann (Cornell University, Ithaca, NY) for providing the L. monocytogenes strains used in this study.
Disclosure Statement
No competing financial interests exist.