
Abstract
Escherichia coli is the most common cause of urinary tract infection (UTI). Phylogroup B2 and D isolates are associated with UTI. It has been proposed that E. coli causing UTI could have an animal origin. The objective of this study was to investigate the phylogroups and antimicrobial resistance, and their possible associations in E. coli isolates from patients with UTI, community-dwelling humans, broiler chicken meat, broiler chickens, pork, and pigs in Denmark. A total of 964 geographically and temporally matched E. coli isolates from UTI patients (n = 102), community-dwelling humans (n = 109), Danish (n = 197) and imported broiler chicken meat (n = 86), Danish broiler chickens (n = 138), Danish (n = 177) and imported pork (n = 10), and Danish pigs (n = 145) were tested for phylogroups (A, B1, B2, D, and nontypeable [NT] isolates) and antimicrobial susceptibility. Phylogroup A, B1, B2, D, and NT isolates were detected among all groups of isolates except for imported pork isolates. Antimicrobial resistance to three (for B2 isolates) or five antimicrobial agents (for A, B1, D, and NT isolates) was shared among isolates regardless of origin. Using cluster analysis to investigate antimicrobial resistance data, we found that UTI isolates always grouped with isolates from meat and/or animals. We detected B2 and D isolates, that are associated to UTI, among isolates from broiler chicken meat, broiler chickens, pork, and pigs. Although B2 isolates were found in low prevalences in animals and meat, these sources could still pose a risk for acquiring uropathogenic E. coli. Further, E. coli from animals and meat were very similar to UTI isolates with respect to their antimicrobial resistance phenotype. Thus, our study provides support for the hypothesis that a food animal and meat reservoir might exist for UTI-causing E. coli.
Introduction
U
UTI is most often caused by Escherichia coli belonging to phylogroup B2, and to a lesser extent phylogroup D (Picard et al., 1999; Escobar-Paramo et al., 2004a; Johnson et al., 2005d; Moreno et al., 2008). UTI is generally believed to be caused by bacteria—often the dominant isolate—from the host's own fecal flora (Gruneberg, 1969; Yamamoto et al., 1997; Moreno et al., 2006; Moreno et al., 2008). The origin of these fecal isolates with uropathogenic potential in the human intestine is so far unknown. One hypothesis is that the uropathogenic E. coli may originate from contaminated food sources (Ramchandani et al., 2005; Johnson et al., 2005a). Because of the use of antimicrobials in food animals, meat is frequently contaminated with antimicrobial-resistant E. coli. In a recent American study, the investigators demonstrated a close resemblance between foodborne E. coli and E. coli causing UTI and urosepsis (Johnson et al., 2005b). This suggests a possible transmission of E. coli via food products. If contaminated meat transmits E. coli that can colonize and infect humans, this may have substantial public-health and medical significance. Yet, only a few studies have been performed to evaluate this hypothesis. To our knowledge, most studies have investigated retail meat isolates only and/or human isolates (Ramchandani et al., 2005; Johnson et al., 2005a, 2005b, 2007), or had small sample sizes and little or no epidemiologic data were available (Johnson et al., 2003, 2004, 2006a). Further, geographic and climate factors seem to influence the population structure of E. coli, and to our knowledge studies of large strain collections from animals, meat, and humans in Northern Europe are missing (Duriez et al., 2001; Escobar-Paramo et al., 2004b; Escobar-Paramo et al., 2006).
Strains of E. coli from animals, meat, community-dwelling humans, and UTI patients were collected in Denmark. The number of isolates, the sampling scheme, and the epidemiological information make this strain collection unique. Using this material it was possible to test the hypothesis of an association between E. coli from animals and UTIs in humans. In 2004 up to 80,389 tons of broiler chicken meat and 148,649 tons of pork was consumed in Denmark (population: 5.5 million) (DANMAP, 2004; DANMAP, 2006). The chicken broiler meat and pork constituted the majority of the meat consumption, making this the focus of our study.
In this study, our objective was to investigate the phylogenetic background and antimicrobial resistance profile, and their possible associations in E. coli isolates from patients with UTI, community-dwelling humans, broiler chicken meat, broiler chickens, pork, and pigs in Denmark.
Materials and Methods
Source of strains and sampling schemes
The 102 E. coli urine isolates were collected from November 2005 to April 2006 in a general practitioner clinic (not associated to any hospital) from patients with community-acquired uncomplicated or complicated UTI. The clinic, which was based south of Copenhagen, consisted of 10 general practitioners serving approximately 10,800 inhabitants. Patients with UTI were included in the study only if they showed typical symptoms of UTI and delivered a midstream urinary sample taken after washing the external urethral meatus with sterile saline, and the urine sample was positive for leucocytes by the dipstick test and revealed at least ≥103 CFU/mL of a typical urinary pathogen (Sobel and Kaye, 2005).
In 2004, about 109 fecal E. coli isolates were collected from humans in the community (community-dwelling humans) selected through the Danish Civil Registration system (a continuously updated register of all residents in Denmark) (scientific ethics committee approval (KF) 01-006/02) (DANMAP, 2004). Participants invited for the study were chosen by a selection algorithm so that they represented the age and sex distribution of the total Danish population taking the differential participation rates of various demographic groups into account. A total of 988 individuals were invited for the study and 125 confirmed their participation by returning the signed consent form. A total of 109 fecal E. coli isolates and questionnaire information on any antibiotics taken were obtained from 109 community-dwelling humans. The questionnaire information was used for interpreting the resistance data on community-dwelling human isolates. All E. coli isolates from community-dwelling humans, meat, and animals were collected in 2004 as part of the Danish Integrated Antimicrobial Resistance Monitoring and Research Program (DANMAP, 2004). Meat samples were collected throughout the year at randomly chosen wholesale and retail outlets in all regions of Denmark by the Regional Veterinary and Food Control Authorities, and represented the meat products sold in Denmark in 2004. One isolate was taken from each meat sample. In total, 197 E. coli isolates from Danish broiler chicken meat, 86 isolates from imported broiler chicken meat, 177 isolates from Danish pork, and 10 isolates from imported pork were included in the study. Fecal samples from Danish broiler chickens and Danish pigs were collected at slaughter in 2004 according to a stratified random sampling scheme by company staff or the Danish meat inspection staff. The number of samples taken at each slaughter house was proportional to the number of animals slaughtered each year at that slaughter house. The slaughter houses included processed 95% of the broiler chickens and 95% of the pigs slaughtered in Denmark. Samples from broiler chickens were collected weekly and samples from pigs were collected monthly. Only one animal per herd or flock was sampled. One isolate was taken from each sample. A total of 138 and 145 E. coli fecal isolates were obtained from broiler chickens and pigs, respectively. The meat from the slaughter houses was distributed nationwide.
Only one isolate per patient, community-dwelling human, meat sample, or food animal was included in this study. The community-dwelling human isolates were identified as E. coli using API20E (Biomérieux, France) and the UTI, meat, and animal isolates by indole, citrate, methyl red, and Voges–Proskauer reaction (DANMAP, 2004). The UTI isolates were deliberately sampled later than the community-dwelling human, animal, and food isolates to allow time for consumption of the meat and possible colonization and invasiveness of the E. coli isolates.
Determination of phylogroups
The phylogenetic background (A, B1, B2, and D) of all isolates was determined by triplex polymerase chain reaction using three DNA markers (Clermont et al., 2000). Results obtained allowed classification of the isolates into the four major E. coli phylogenetic lineages or nontypeable (NT) isolates according to Gordon et al., (A: yjaA positive; B1: TSPE4.C2 positive; B2: chuA and yjaA positive or chuA, yjA, and TSPE4.C2 positive; D: chuA positive or chuA and TSPE4.C2 positive; NT: yjaA, chuA, and TSPE4.C2 negative) (Gordon et al., 2008). E. coli strains ECOR 20 (yjaA), ECOR 48 (chuA), ECOR 58 (tspe4.c2), ECOR 62 (chuA, yjaA, and tspe4.c2) were used as positive controls.
Antimicrobial susceptibility
Susceptibility to ampicillin (1–32 mg/L), ceftiofur (0.5–8 mg/L), chloramphenicol (2–64 mg/L), ciprofloxacin (0.03–4 mg/L), gentamicin (1–32 mg/L), streptomycin (4–64 mg/L), sulfamethoxazole (64–1024 mg/L), tetracycline (2–32 mg/L), and trimethoprim (4–32 mg/L) was determined for all isolates by a micro broth dilution method (Trek Diagnostics Systems, East Grinstead, United Kingdom). The antimicrobial agents tested were chosen because of their use in human antimicrobial therapy of E. coli infections. Tetracycline is used mostly in veterinary therapy and was therefore included as a potential marker of antimicrobial resistance from animals. Ceftiufur is used for animals only. However, it was included as a representative for third-generation cephalosporins and could be used to detect possible extended-spectrum beta-lactamase positive isolates. E. coli ATCC 25922 was used for quality control. Results were interpreted according to the Clinical Laboratory Standards Institute except for ciprofloxacin data, which were interpreted according to The Danish Reference Group for Antibiotic Susceptibility Testing (NCCLS, 2003; DSKM, 2004). Multiresistant isolates were defined as isolates resistant to three or more of the tested antimicrobial agents.
Statistical analysis
The individual isolate was the unit of statistical analysis. Possible associations between phylogroups and antimicrobial resistance, and the distribution of phylogroups between isolate origins were investigated by comparing proportions using Fisher's exact test (two-tailed) with a significance level of p ≤ 0.05 (GraphPad Prism 5; GraphPad Software, San Diego, CA).
We analyzed how the E. coli isolates clustered within their phylogenetic group (A, B1, B2, D, and NT) according to their antimicrobial resistance phenotype using PROC CLUSTER in SAS 9.2, (SAS Institute, Cary, NC). In the cluster analysis, we used Jaccard's coefficient matching resistance with resistance. Hence, fully susceptible isolates were noninformative and excluded from the analysis. Ward's minimum variance clustering method was used and the number of clusters was based on maximizing the number of resistances shared by all isolates in a cluster. The results were observed in a dendrogram (PROC TREE in SAS 9.2).
Results
Antimicrobial resistance
Ampicillin, chloramphenicol, ciprofloxacin, streptomycin, sulfamethoxazole, tetracycline, and trimethoprim resistance was observed in isolates from all origins except for the isolates from broiler chicken (chloramphenicol susceptible) and imported pork (chloramphenicol and ciprofloxacin susceptible) (Table 1). Gentamicin resistance was only observed in one UTI isolate. Ceftiofur resistance was not detected in any isolates from food animals, meat, community-dwelling humans, or UTI patients. The resistance frequencies toward the tested antimicrobials varied among the different origins (0–58%) (Table 1). Among isolates from patients, community-dwelling humans, Danish and imported broiler chicken meat, Danish and imported pork, and pigs, antimicrobial resistance frequencies were highest for ampicillin, streptomycin, sulfamethoxazole, and tetracycline (Table 1). Among isolates from broiler chickens, antimicrobial resistance frequencies were highest toward ampicillin, sulfamethoxazole, tetracycline, and ciprofloxacin (Table 1). Ciprofloxacin resistance was most frequently observed in isolates from Danish (7%, n = 13) and imported broiler chicken meat (22%, n = 19) and broiler chickens (12%, n = 17) (Table 1). Of note, antimicrobial resistance to all of the antimicrobials tested and multiresistance were detected more frequently in isolates from imported broiler chicken meat compared with Danish broiler chicken meat (p ≤ 0.0004) (Table 1 and Fig. 1A). Low to moderate frequencies of multiresistance were also observed in isolates from other origins (Fig. 1A). A total of 22% of the imported broiler chicken meat isolates were susceptible to all eight investigated antimicrobial agents. In contrast, 46–66% of the other isolates were fully susceptible (Fig. 1A). Of the 109 community-dwelling humans, two reported having taken antibiotics within 1 month before the sampling, two humans with multiresistant isolates had been in contact with broiler chickens and none with pigs, and most humans reported consuming broiler chicken meat and/or pork several times a week.

(
Gentamicin resistance was not included in the table as only one UTI patient isolate was resistant. All isolates were susceptible to ceftiofur. Some isolates were resistant to more than one antimicrobial.
Phylogenetic distribution
Isolates belonging to all four major phylogenetic lineages and NTs were found among isolates from all origins except for imported pork isolates, where D isolates were absent among the 10 isolates (Fig. 1B). B2 isolates were the predominant phylogroup among isolates from UTI patients and community-dwelling humans (Fig. 1B). Phylogroup A was the dominant phylogroup among isolates from Danish and imported broiler chicken meat, broiler chickens, Danish and imported pork, and pigs (Fig. 1B). Using Fisher's exact test to analyze proportions, we found that the phylogroups were distributed similarly among isolates from UTI patients and community-dwelling humans (p = 0.144). Isolates from Danish and imported broiler chicken meat, and broiler chickens (p = 0.053), and isolates from Danish and imported pork, and pigs (p = 0.055), respectively, were different in their phylogroup distributions.
Antimicrobial resistance and phylogroups
Disregarding the imported pork isolates due to the low number (n = 10), isolates representing phylogroups A, B1, D, and NT isolates from all origins were resistant to one or more of the same five antimicrobial agents (ampicillin, streptomycin, sulfamethoxazole, tetracycline, and trimethoprim) (Fig. 2A, B, D, E). Isolates representing phylogroup B2 were resistant toward one or more of ampicillin, streptomycin, and sulfamethoxazole regardless of origin (Fig. 2C). The antimicrobial resistance patterns observed for the different origins varied from phylogroup to phylogroup or NT group (Fig. 2A–E). Statistical associations between specific phylogroups and specific antimicrobial resistance were investigated using Fisher's exact test. We found significant associations between antimicrobial resistance and phylogroups among isolates from different origins (UTI patients, broiler chickens, Danish and imported pork, and pigs) (Table 2).
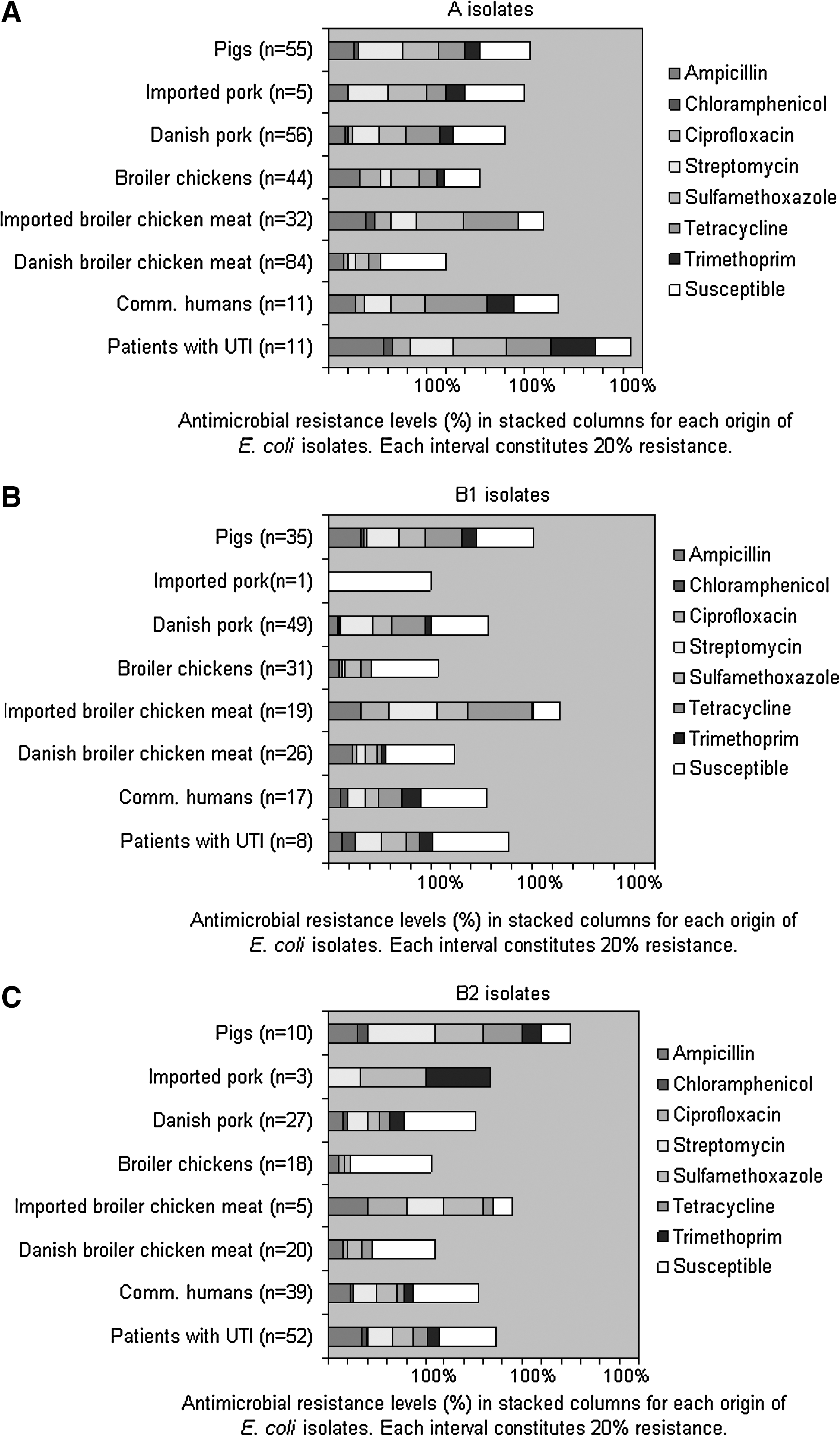
Antimicrobial resistance frequencies (%) in (
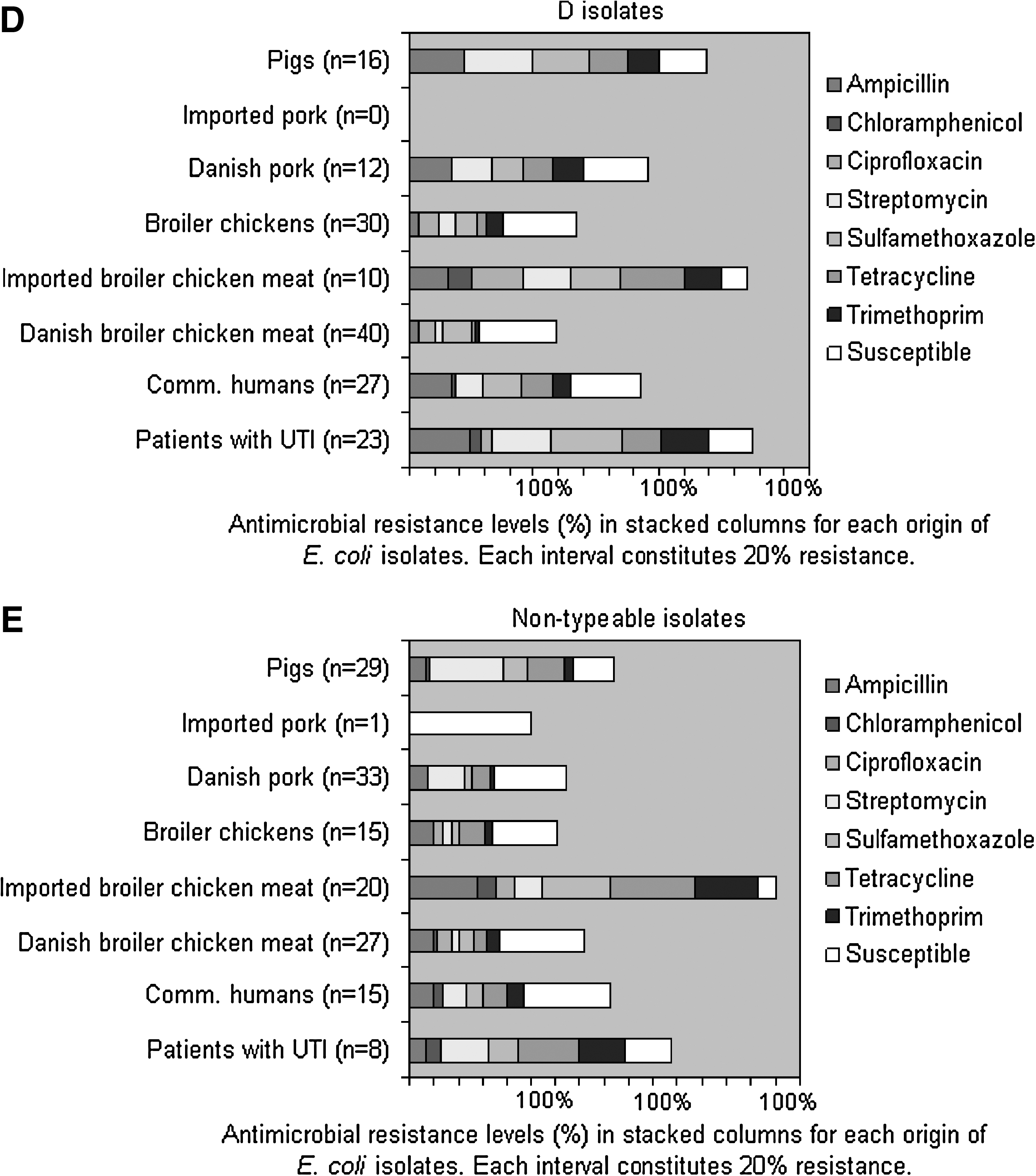
Only isolates of the same origin belonging to A, B1, B2, or D were compared. Fisher's exact test was used.
AMP, ampicillin; SUL, sulfamethoxazole; TET, tetracycline; TRI, trimethoprim; R, resistant; S, susceptible.
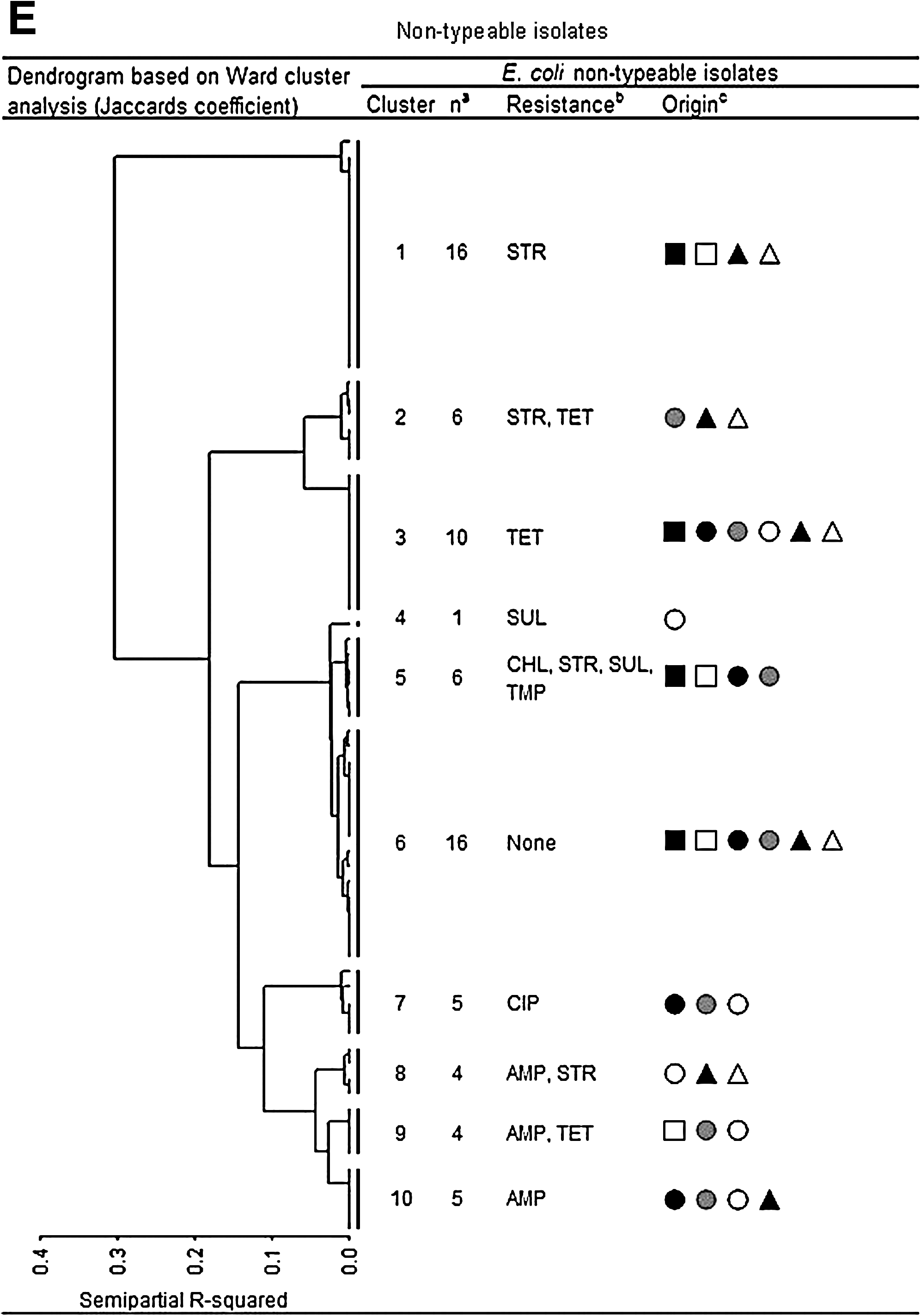
Clustering of isolates
When analyzing the antimicrobial resistance data using Ward's minimum-variance method, we found that the UTI isolates clustered together with isolates from meat and/or animals, indicating similar antimicrobial resistance phenotypes across the reservoirs (Fig. 3A–E). Often the UTI isolates clustered with both broiler chicken/broiler chicken meat as well as pork/pig isolates. UTI isolates also often grouped with isolates from community-dwelling humans.
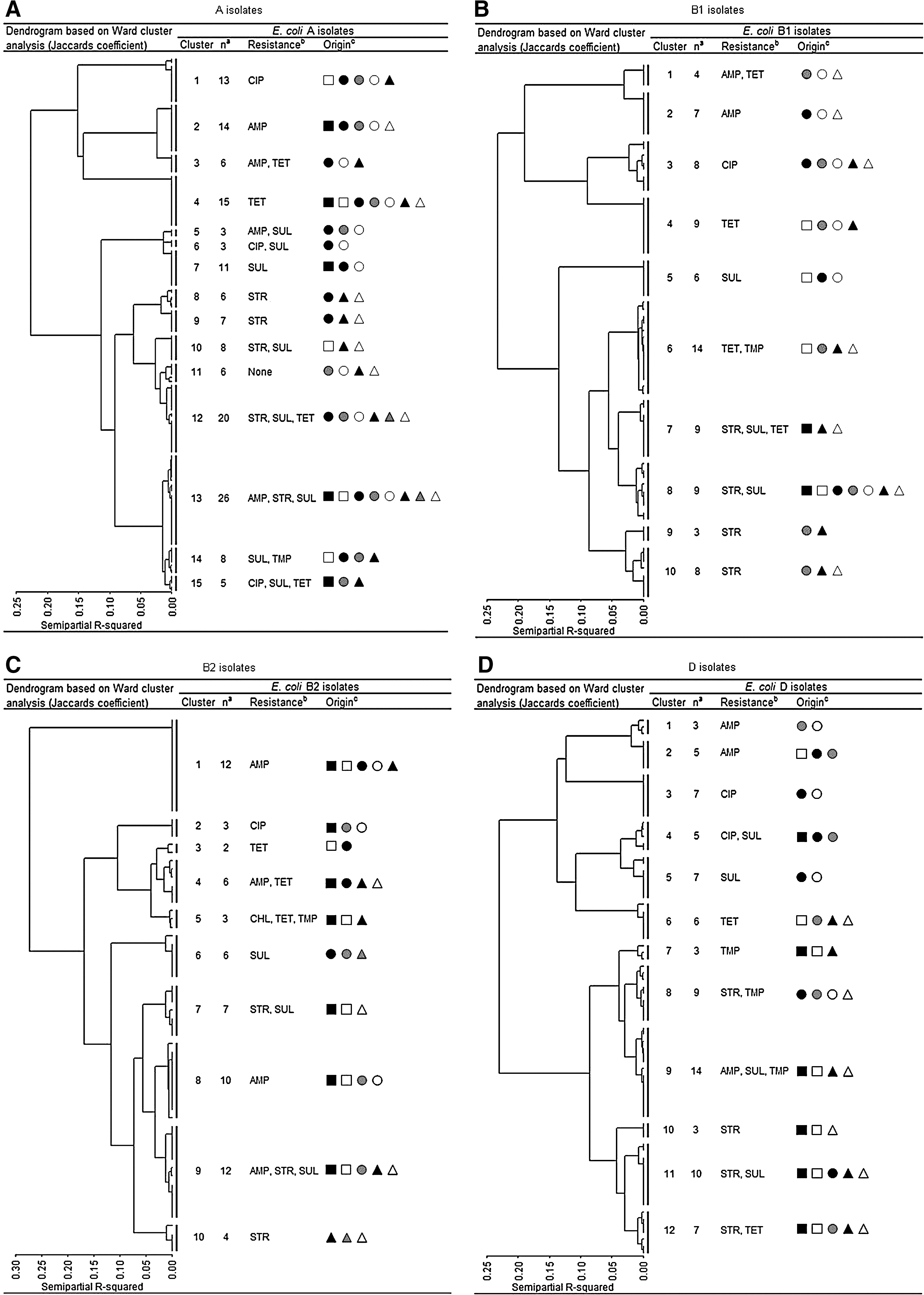
Dendrogram (Ward's Minimum-Variance method using Jaccards coefficient) of E. coli isolates belonging to the four phylogroups ((
Discussion
This is the first study to present data on antimicrobial resistance phenotypes and phylogroups of a large collection of animal, meat, community-dwelling human, and human urinary tract E. coli isolates obtained from the same geographical region and in the same time period using carefully designed sampling schemes to ensure epidemiological representativeness.
The presence of B2 and D isolates (the phylogroups that are related to UTI) of animal origin in our study with similar resistance patterns to UTI isolates suggests that food animals and meat might be a source of such isolates to humans. Consistent with previous findings, B2 isolates predominated among isolates from patients and community-dwelling humans (Zhang et al., 2002; Johnson et al., 2005c).
Although B2 isolates were present only in low frequencies in food animals and meat (6–15%), B2 isolates are much more successful in colonizing the human intestine than A and or B1 isolates probably due to the virulence genes present in B2 isolates (Picard et al., 1999). It is usually the host's dominant fecal isolate that causes UTI (Gruneberg, 1969; Yamamoto et al., 1997; Moreno et al., 2006; Moreno et al., 2008). Animals and meat may pose a risk for intestinally acquiring E. coli of uropathogenic potential. Thus, even low prevalence of B2 isolates (7–30%) among the E. coli from broiler chicken meat, broiler chickens, pork, and pigs may pose a risk for acquiring E. coli.
It has been suggested that B2 E. coli isolates are less resistant to antimicrobial agents than non-B2 isolates (Johnson et al., 2003). We could confirm this for our UTI isolates, Danish and imported pork, and broiler chicken isolates. We could not find any other significant associations for any of the other isolate origins, possibly due to low frequencies of antimicrobial resistance.
A minor proportion of the E. coli isolates from patients and community-dwelling humans belonged to group A (10–11%) and group B1 (8–16%). Spread of clinical E. coli clones belonging to phylogroup A (e.g., ST10 and ST23) and B1 (e.g., ST359 and ST155) have been reported (Oteo et al., 2009). The broiler chicken meat and pork were dominated by A and B1 isolates (56–60%). Consuming broiler chicken meat and pork may therefore more often expose the human intestine to A and B1 isolates than B2 and D isolates, and although A and B1 isolates may be less successful colonizers compared with B2 isolates, A and B1 isolates may predominate the fecal E. coli population in community-dwelling humans transiently from time to time (Picard et al., 1999). If we accept the theory of the dominant fecal isolate causing the UTI, then these A and B1 isolates from meat may cause UTI (Gruneberg, 1969; Yamamoto et al., 1997; Moreno et al., 2006, 2008).
Through cluster analysis of the antimicrobial resistance phenotypes of all isolates within the individual phylogroups and the NT isolates, we found that antimicrobial-resistant UTI isolates appeared very similar to resistant meat and animal isolates. This indicates that broiler chicken meat, broiler chickens, pork, or pigs contaminated with antimicrobial-resistant E. coli could be a source of resistant isolates in community-dwelling humans and UTI patients. Antimicrobial resistance in E. coli from community-dwelling humans was similar to resistance frequencies in Danish broiler chicken meat, broiler chickens, and Danish and imported pork isolates. The antimicrobial resistance in community-dwelling humans was most likely not antimicrobial consumption driven because—to our knowledge—only 2 of 109 community-dwelling humans had received antimicrobial treatment within 1 month before sampling. One of these persons yielded a fully susceptible isolate. A recent case–control study identified frequent chicken and pork consumption as a risk for community-acquired UTI caused by antimicrobial-resistant E. coli (Manges et al., 2007), and Corpet concluded that diet was very important for tetracycline-resistant E. coli in healthy humans (Corpet, 1988). Most of the community-dwelling humans in this study reported consuming broiler chicken meat and/or pork several times a week. No information on the dietary habits of the UTI patients was available. None of the community-dwelling humans with multiresistant E. coli isolates reported being in contact with pigs. Two out of 22 community-dwelling humans yielding multiresistant isolates had been in contact with broiler chickens. Therefore, we assume that if food and food animals were a source of resistant E. coli recovered from community-dwelling humans in this study, direct contact accounted for only a small fraction of the transmission. We do not know if the UTI patients had been in contact with animals. Horizontal transfer of antimicrobial resistance between E. coli of animal, meat, and human origin was not directly investigated in this study. However, ampicillin, streptomycin, sulfamethoxazole, tetracycline, tetracycline, chloramphenicol and the recently reported plasmid-mediated ciprofloxacin resistances are all shown to transfer horizontally (Johnson et al., 2006b; Kikuvi et al., 2007; Jakobsen et al., 2008; Hawkey and Jones, 2009). Therefore, this mode of transmission is possible.
Our study had some limitations. The UTI patient isolates were limited to one region in Denmark. Further, only one isolate obtained per community-dwelling human, meat sample, and animals was investigated (Lautenbach et al., 2008). The participant rate of community-dwelling humans was lower than expected. The UTI isolates were deliberately sampled later than animal, food, and community-dwelling human isolates. However, since there is limited knowledge on the period from ingestion to a possible episode of UTI, this may influence the relevance of any sampling period for UTI isolates. The cluster analysis was limited to antimicrobial resistance phenotypes only and thus excluded fully susceptible isolates. Further, isolates with similar phenotypic resistance profile may have different genotypic profiles. However, in lack of high trough-put affordable genotyping methods the phenotypes do still provide valuable information. The strengths of this study included the stratified random sampling scheme for food animal, meat, and community-dwelling human isolates, making the distribution of phylogroups and occurrence of antimicrobial resistance representative of the distribution and occurrence in the populations. The sampling of all isolates was carried out during the same time period, with a lag period for the UTI isolates, and across the same geographical area. This approach offers a representative sample of isolates for comparison. This study also included a large sample of isolates across a wider ranger of food animal, meat, and human sources in contrast to previous studies (Johnson et al., 2003, 2004, 2005a, 2005b, 2006a, 2007; Ramchandani et al., 2005).
Conclusion
The main finding of this study was detecting B2 and D isolates—the phylogroups that are related to UTI—among isolates from broiler chicken meat, broiler chickens, pork, and pigs that were similar in antimicrobial resistance phenotype to E. coli from UTI patients. The cluster analysis proved resemblance of animal and food isolates to UTI isolates. This provides support for the hypothesis that a food animal and meat reservoir might exist for UTI-causing E. coli (Ramchandani et al., 2005; Manges et al., 2007; Johnson et al., 2008, 2009a, 2009b; Hannah et al., 2009). Due to the good colonizing abilities of B2, even a low prevalence of such phylogroup isolates in meat and food animals could pose a risk for acquiring uropathogenic E. coli gastrointestinally. More studies are needed to confirm that uropathogenic E. coli originate from contaminated food and animals and possibly other sources as well (Skyberg et al., 2006; Johnson et al., 2007).
Footnotes
Acknowledgments
Frank Hansen and Karin Sixhøj Pedersen are thanked for excellent technical assistance. The Danish Research Council is thanked for financial support (Grant no. 2101-05-001). This work is part of the Danish Integrated Antimicrobial Resistance Monitoring and Research Program (DANMAP) and the Marie Curie program Training Risk-Assessment In Non-human Antibiotic Usage (TRAINAU).
Disclosure Statement
No competing financial interests exist.
Part of this work was presented at:
American Society of Microbiology 107th General Meeting, May 2007, Toronto, Canada, abstract no. Y-049;
Twenty-fourth Annual Meeting of the Scandinavian Society for Antimicrobial Chemotherapy, September 2007, Tampere, Finland;
Eighteenth European Congress of Clinical Microbiology and Infectious Diseases, April 2008, Barcelona, Spain, abstract no. P-1625;
DANMAP 2004. Use of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from food animals, foods and humans in Denmark. ISSN 160-2032;
ICAAC 2007 47th Interscience Conference on Antimicrobial Agents and Chemotherapy, September 2007, Chicago, IL, abstract no. D-1592.