
Abstract
In this study, we characterized 110 strains of Salmonella enterica serovar Bovismorbificans contaminating environment, animals, food of animal origin, and human, to assess their significance along the food chain in Hungary. Additionally, five strains from Germany were tested for comparative purposes. Characterization involved antibiotic susceptibility testing, class 1 integron detection by polymerase chain reaction, plasmid profiling, virulotyping (using virulence gene-specific polymerase chain reactions), and pulsed-field gel electrophoresis. Pathogenic potential of selected strains was tested in orally infected 1-day-old specific pathogen-free chicks. Eighty-two percent of the strains were susceptible to the 16 antibiotics tested, and none of them had class 1 integron. A multidrug-resistant human isolate harbored a bla SHV5-type extended-spectrum β-lactamase gene, first reported in this serotype. All the strains possessed avrA, ssaQ, mgtC, spi4, and sopB genes indicating the presence of Salmonella pathogenicity islands 1–5, respectively, missed the phage-related genes sopE and gipA, but retained the phage-related gene sodC1. An ∼90 kb large plasmid was characteristic to 80% of the strains, all of which carried the spvC gene. In vivo colonization testing of four selected strains in 1-day-old chicks resulted in significantly reduced liver and spleen colonization ability as compared with the Salmonella Enteritidis control strain, whereas their caecal colonization ability differed less from that of Salmonella Enteritidis. Pulsed-field gel electrophoresis data revealed the dominance of two pulsotypes (C2 and C5) without any specific temporal, geographical, and/or source-related linkages. The results show that Salmonella Bovismorbificans studied here are less invasive than Salmonella Enteritidis, but they may colonize and persist in several animal species and successfully contaminate meat products of different animal origin in Hungary.
Introduction
S
Materials and Methods
Bacterial strains and samples
There were 110 Hungarian strains of Salmonella Bovismorbificans included in this study. For comparative purposes, additional five strains of German Turkey origin (kindly donated by Dr. Burkhard Malorny, BfR, Berlin) were included. The animal, food, feed, and environmental isolates were referred to the National Food Investigation Institute (National Salmonella Reference Laboratory), Budapest, from county veterinary and food safety laboratories at different parts of the country, based on different sampling activities, using harmonized methods during 1999–2006. The animal and food isolates represented fecal samples, as well as raw and processed meat and meat products of poultry (n = 35) (one duck, one goose, and the rest of chicken origin), swine (n = 50), bovine (n = 8), and game (n = 1) origin. One strain was isolated from pasta. The environmental strains were from hatchery drug swabs (n = 4), and the feed isolates were of fish powder (n = 1) and meat powder (n = 2) origin. The human isolates (n = 8) represented stool samples of sporadic cases received during 2005–2006 by the Phage Typing and Molecular Epidemiology Department of the National Center for Epidemiology from several public health laboratories of the country. All strains were serotyped according to the Kauffmann–White scheme.
Antibiotic susceptibility testing and detection of extended-spectrum β-lactamase genes
Antibiotic susceptibility testing of all the isolates was done by disc-diffusion method on Mueller–Hinton agar using antibiotic discs (Oxoid Ltd., Basingstoke, UK) as follows: ampicillin (10 μg), amoxicillin (10 μg), cefotaxime (30 μg), chloramphenicol (30 μg), ciprofloxacin (5 μg), enrofloxacin (5 μg), gentamycin (10 μg), kanamycin (30 μg), nalidixic acid (30 μg), neomycin (30 μg), spectinomycin (100 μg), streptomycin (10 μg), sulphonamide (300 μg), trimethoprim (5 μg), trimethoprim–sulfamethoxazole (1.5/23.75 μg), and tetracycline (30 μg). The zones of growth inhibition and the breakpoints were evaluated according to the Clinical and Laboratory Standards Institute recommendations (CLSI, 2005). The Escherichia coli ATCC 25922 was used as a reference strain. The phenotypic confirmation of the extended-spectrum β-lactamase (ESBL) production of the cefotaxime resistant isolate was performed by double-disc approximation test and Etests (Ab Biodisk, Solna, Sweden). The ESBL-producing isolate was tested for the presence of the bla TEM, bla CTX-M, and bla SHV genes by polymerase chain reaction (PCR) (Damjanova et al., 2008). The obtained bla SHV amplicon was purified and sequenced. The sequence obtained was Basic Local Alignment Search Tool (BLAST) analyzed and compared with those registered in the GenBank. All the strains were tested for the presence of class 1 integrons by PCR according to Lévesque et al. (1995).
Macrorestriction analysis by PFGE
PFGE was carried out according to the standardized Salmonella protocol of the CDC PulseNet (Ribot et al., 2006). The Salmonella Braenderup H9812 strain was used as molecular standard. PFGE-generated DNA profiles were entered into the Fingerprinting II Software (Bio-Rad Laboratories, Ventura, CA) for analysis. Cluster analysis was performed by the unweighted pair-group method with arithmetic averages; DNA relatedness was calculated on the basis of the Dice coefficient. A 1.0% position tolerance and 1.0% optimization setting were applied. Being uncertain to detect, bands that were smaller than 33.3 kb were not included in the analysis.
Plasmid analysis
Plasmid preparation was conducted by the alkaline lysis method of Kado and Liu (1981). Agarose gel electrophoresis was performed using a 0.75% agarose in a vertical system. The approximate sizes of plasmids were estimated by comparing them with the reference plasmids of E. coli V517 (2.0–53.7 kb) and E. coli R27 (168 kb) using the Quantity One software (Bio-Rad Laboratories).
Virulence gene PCRs
The presence of the Salmonella plasmid virulence gene spvC was detected by the PCR method described by Haneda et al. (2001). To localize the spvC gene on some selected plasmids by Southern hybridization, the digoxigenin-high prime DNA labeling and detection starter kit (Roche Diagnostics GmbH, Mannheim, Germany) were used. Digoxigenin-labeled spvC PCR product obtained from a randomly selected Salmonella Bovismorbificans strain was used as a probe. The probe target hybrids were observed by chromogenic assay. Throughout the whole process, instructions of the manufacturer were followed. The presence of the chromosomal virulence genes avrA, gipA, mgtC, sodC1, sopB, sopE, spi4, and ssaQ was detected according to the methods of La Ragione et al. (2007) and Malorny et al. (2007).
In vivo colonization and organ invasion
As the result of PCR screening for the selected nine virulence genes revealed that the strains varied only in the presence or absence of the spvC gene, the question was raised whether it may affect the in vivo virulence of the strains. Therefore, two strains with spvC gene (443 and 482) and two others without spvC (715 and 3733) were selected to test their in vivo caecal and organ colonization in a 1-day-old chicken model. The strains 443 and 482 were isolated as resistant to nalidixic acid, whereas the strains 715 and the German strain 3733 were marked by nalidixic acid resistance by passing isolated colonies on Mueller–Hinton agar plates containing nalidixic acid in increasing concentration. Specific pathogen-free 1-day-old Ross chicks, supplied from the Bábolna hatchery, Hungary, were housed in plastic boxes in groups of six. In the four experimental groups, the birds were inoculated orally on the day of arrival with 0.5 mL of the stationary-phase nutrient bouillon culture of Salmonella Bovismorbificans 443 NalR, 482 NalR, 715 NalR, and 3733 NalR. Inocula contained ∼2–5 × 107 colony-forming units/mL bacteria. One group of chicks was inoculated with the highly invasive Salmonella Enteritidis 147 NalR strain (Methner et al., 2004) and was used as a control group for invasiveness. Another group of chicks remained uninoculated (intact control group). Five days after the inoculation, chicks from each group were killed humanely, and ∼0.2 g of the caecal contents as well as ∼0.2 g of spleen and liver of each bird were removed and homogenized (Stomacher®, Seward, UK) in Rappaport–Vassiliadis (RV) broth in a ratio of 1:10 for caecal content and 1:50 for liver and spleen. Thereafter, decimal dilutions were made in RV broth up to 10−6. Dilutions of caecal contents were immediately plated onto Bromothymol blue–Lactose (BTB) agar plates containing nalidixic acid (50 μg/mL−1) that allowed quantitative assessment of colony-forming units per gram of bacteria inoculated. Homogenates of organ samples were diluted in RV broth up to 10−6 as above, and all dilutions were incubated. After 2 days of incubation at 37°C, 10 μL volumes of each cultured dilution steps were plated onto BTB agar plates as above to determine the highest dilution from which Salmonella could be cultured. This way the Salmonella titre of organ samples could be determined. Chicks of the intact control group were processed on the fourth day same way as the experimental groups without nalidixic acid in BTB plates. License for these animal experiments was granted by the Veterinary and Food Control Station, Budapest (No. 273/003/2004). Each in vivo experiment was repeated twice, and the mean values calculated from both experiments are presented. The paired Student's t-test was used to assess whether the mean values obtained for the experimental groups were statistically different from that of the control group.
Results
Antibiotic resistance profiles of the 110 Salmonella Bovismorbificans strains are summarized in Table 1. Vast majority (82%, 90/110) of the Hungarian strains were susceptible to all the antimicrobials tested. A small group of strains (n = 10), mainly of swine origin (n = 8), was resistant to nalidixic acid. The remaining 10 strains of different origin harbored ampicillin (n = 4), amoxicillin (n = 4), kanamycin (n = 4), neomycin (n = 3), streptomycin (n = 4), spectinomycin (n = 2), sulphonamide (n = 3), and/or tetracycline (n = 4) resistances alone, or in combination. None of the strains had class 1 integron. Nine strains were resistant to more than one antimicrobial, and most importantly one of the human isolates proved to be multidrug resistant including cefotaxime resistance, encoded a bla SHV5-type ESBL gene. It is also interesting to note that only the turkey strains of German origin had trimethoprim (n = 3) and trimethoprim–sulphametoxazole (n = 3) resistances. Further, in contrast to the Hungarian strains, four out of the five German strains were resistant to at least two antimicrobials and two of them were multidrug resistant.
Antimicrobial resistance of Salmonella Bovismorbificans strains of German turkey origin isolated in 2006 and 2007 and tested for comparative purposes were as follows: susceptible (n = 1), AmpSu (n = 1), SxtTmpSu (n = 1), AmpAmxSuTmpSxtTet (n = 1), AmpAmxSuTmpSxtTet (n = 1).
Amp, ampicillin; Amx, amoxicillin; Ctx, cefotaxime; Kan, kanamycin; Nal, nalidixic acid; Neo, neomycin; Spe, spectinomycin; Str, streptomycin; Su, sulphonamide; Tmp, trimethoprim; Sxt, trimethoprim-sulfamethoxazole; Tet, tetracycline.
The strains showed a relative genetic homogeneity when investigated for the presence of the most important virulence genes. None of the strains had the gipA and sopE, both of which are located on prophages in the Salmonella genome out of the nine investigated virulence genes. In contrast, all of them harbored the avrA, ssaQ, mgtC, spi4, and sopB genes known to be located on the Salmonella pathogenicity islands 1–5 (SPI-1–5), respectively, and also the phage-related sodC1 gene. Occurrence of the spvC gene, the location of which on an ∼90 kb sized virulence plasmid was proved by hybridization on selected strains (data not shown), was also frequent; it was characteristic to 80% (88/110) of the Hungarian strains, but was not detected in any of the five German strains.
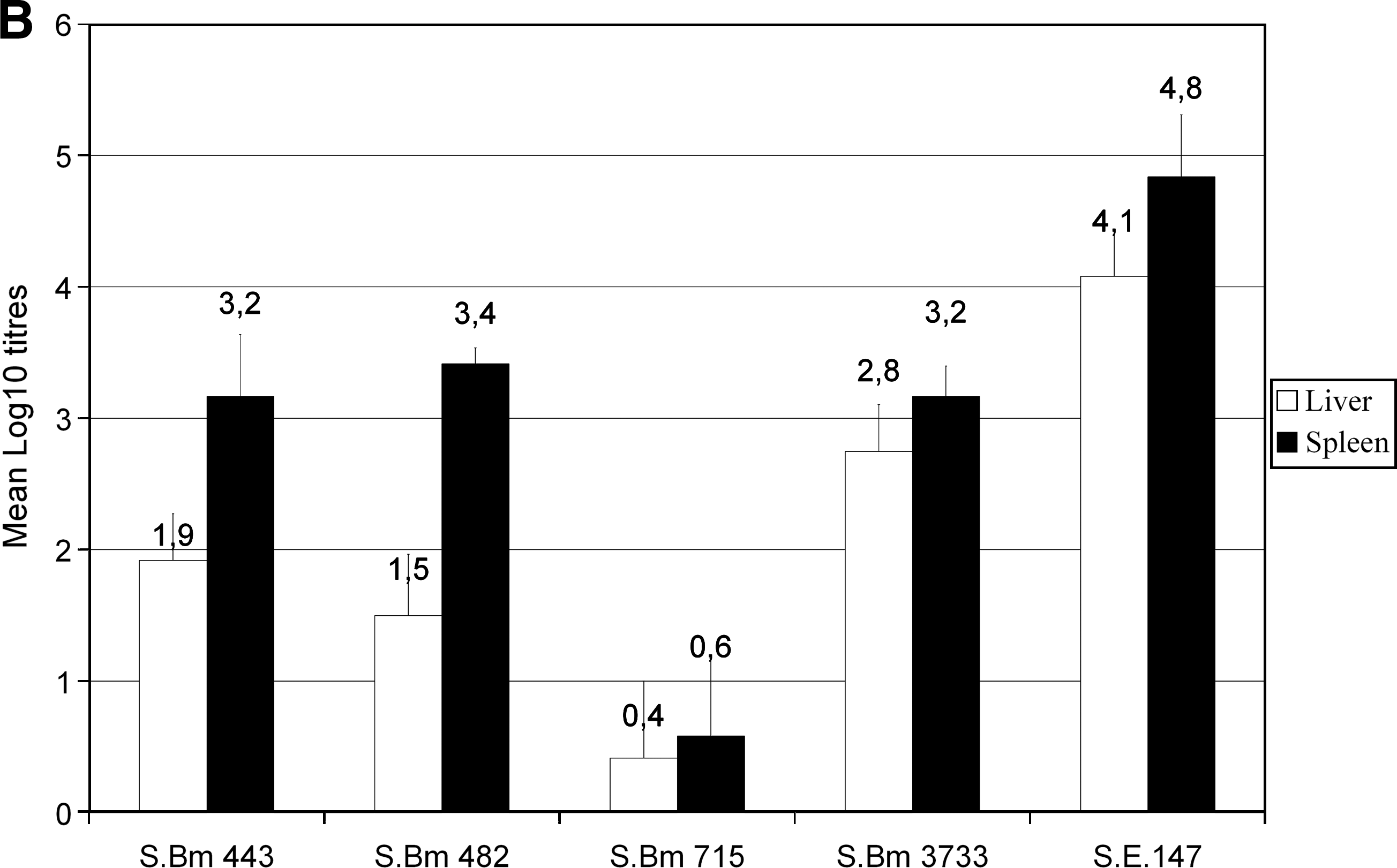
In vivo colonization experiments with some selected strains of Salmonella Bovismorbificans showed significantly reduced caecal colonization values in case of the strains 482 and 3733. Colonization in case of strain 443 was as efficient as that of the Salmonella Enteritidis 147 control, and even more efficient (p < 0.05) colonization was detected in case of strain 715 (Fig. 1A). In contrast, all Salmonella Bovismorbificans strains proved much more significantly (p < 0.05–p < 0.0005) reduced liver and spleen invasion as compared with the Salmonella Enteritidis 147 control strain (Fig. 1B). Strains 443 and 482 contain virulence plasmid and are spvC+ , while strains 715 and the German strain 3733 are both devoid of virulence plasmid and are spvC−. In spite of that common trait, the strain 715 showed significantly (p < 0.05) reduced organ invasion compared with the other virulence plasmid-less strain 3733, suggesting that the presence of the spvC carrier plasmid alone did not influence the colonization and organ invasion ability of the investigated strains.
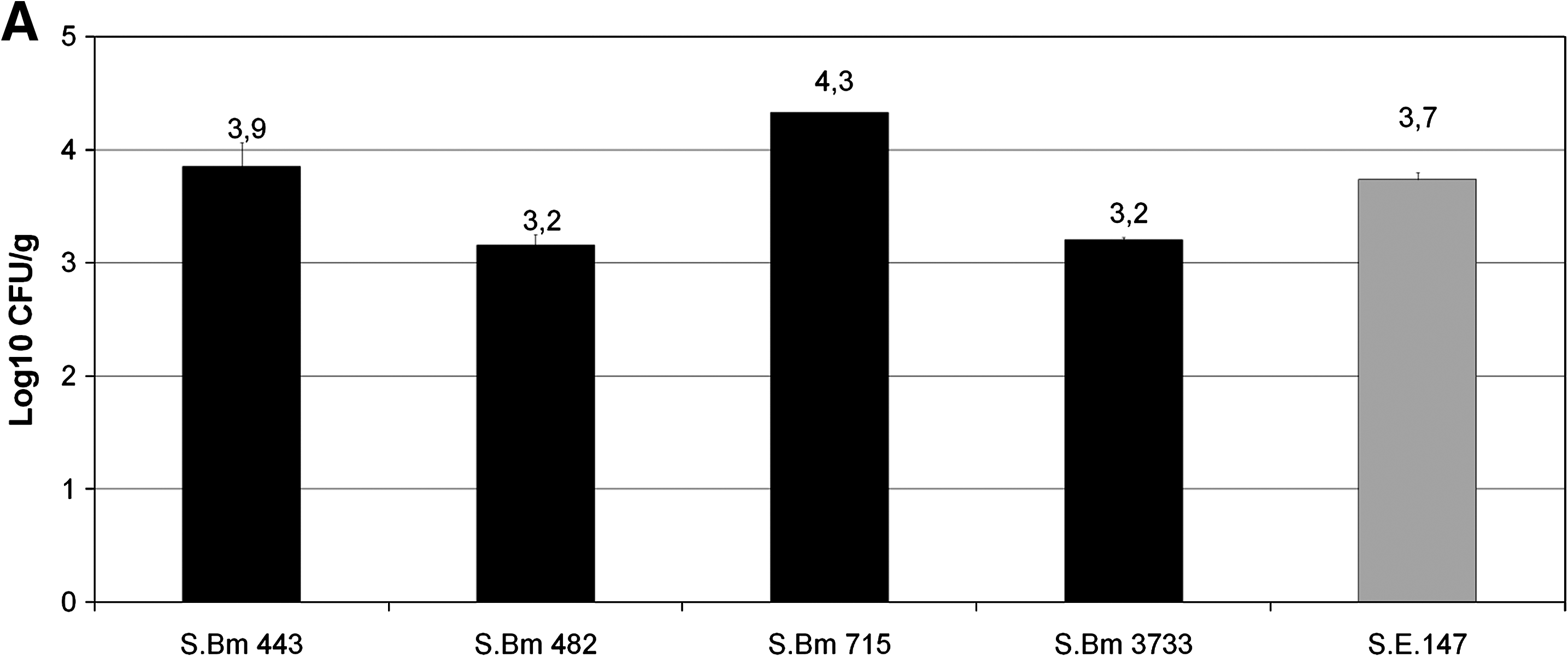
Caecal (
Although the 110 Hungarian strains represented 24 different plasmid profiles, the most frequently detected plasmid was an ∼90 kb serovar-specific plasmid that was present in all spvC harboring strains (Fig. 2). More than half (59%, 65/110) of the strains had this plasmid alone, but lot of them (37%) had other, usually smaller plasmids as well. Small plasmids were characteristic mainly of strains of swine origin. A group of nine swine and the only duck strain from 2004 that also had the same PFGE pattern had a small plasmid approximately the same size. Another group of seven swine and one strain of fish powder origin isolated from 1999–2001 also had a common small plasmid. Only four strains had no plasmid at all. None of the German strains contained the spvC carrier large plasmid, and one was plasmid less. Four of them represented four different plasmid profiles.

Pulsed-field gel electrophoresis (PFGE) patterns, plasmid profiles, and some characteristics of 115 Salmonella enterica serovar Bovismorbificans strains of different origin. *: + indicates the presence, −indicates the lack of the spvC gene product. **Abbreviations used for the antimicrobials are the same as indicated in the footnote of the Table 1. aStrains marked by framing were used in the in vivo experiments.
PFGE analysis carried out by XbaI digestion revealed the existence of 26 different patterns (pulsotypes) (Fig. 2). When one representative strain from each pattern was selected for further similarity analysis, the dendrogram obtained showed that they could be divided into three clusters and six individual types by applying 85% as a similarity cut-off value (Fig. 3). The dominance of one large cluster designated as C, representing 92% (101/115) of the strains, could be observed. Within this cluster, most of the strains (76%, 77/101) belonged to only two patterns: pulsotype C2 (n = 34) and C5 (n = 43, 39 Hungarian plus four German strains).
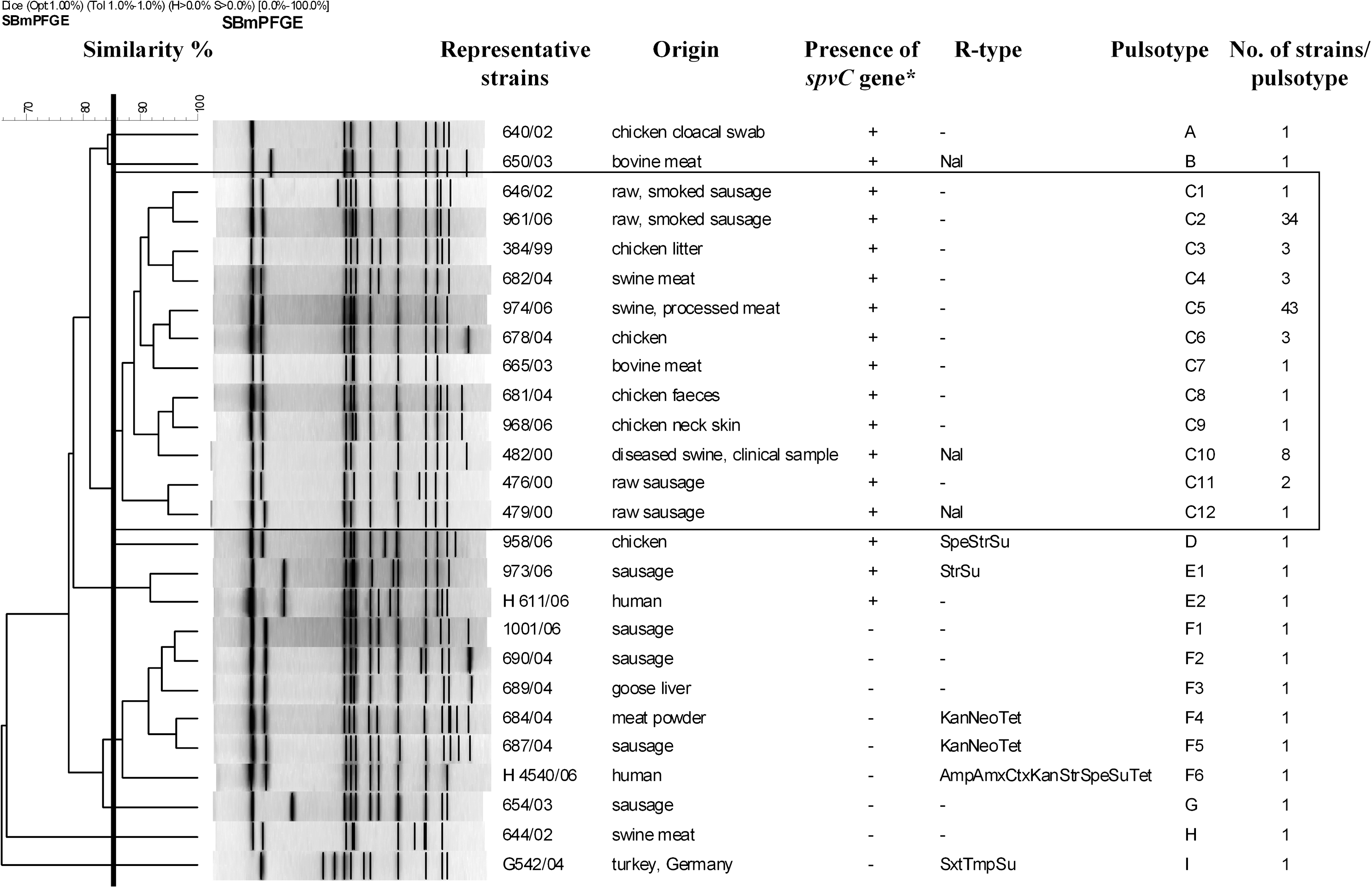
PFGE patterns of representative Salmonella enterica serotype Bovismorbificans strains. Twenty-six strains representing different patterns were selected from the collection of 115 isolates from human, animal, food, feed, and environmental samples to represent 26 pulsotypes. The clusters and individual patterns that were observed using 85% similarity cut-off are indicated by capital letters (A–I). Subtypes are indicated by digits. *: + indicates the presence, − indicates the lack of the spvC gene product. **Abbreviations used for the antimicrobials are the same as indicated in the footnote of the Table 1. Vertical bar is drawn at the 85% similarity value. The most frequent pulsotype (C) is highlighted by framing.
Discussion
Antimicrobial resistance of Salmonella strains is a growing concern as it can limit therapeutic options and may undermine efficacy of empiric treatment (Mølbak, 2005). Moreover, evidence is accumulating to suggest that antimicrobial-resistant strains cause more severe human illnesses than susceptible strains (Varna et al., 2005). There are also examples for the emergence of multidrug-resistant Salmonella strains in animal populations before they emerge in the human population (Angulo et al., 2000). Very few data exist regarding the antimicrobial resistance of Salmonella Bovismorbificans of either human or food and animal origin. In concordance with the findings of Liesegang et al. (2002) who characterized a large collection of Salmonella Bovismorbificans strains of differing sources from several countries, we also found that most of our strains were susceptible to the antimicrobials tested and only few strains exhibited resistance or multidrug resistance. Most of the resistant strains were of swine origin and mostly were resistant to nalidixic acid, which may reflect the use of this antibiotic in the therapy. Indeed, two of these strains originated from sick animals.
It should also be noted that one of the human isolates was multidrug resistant and produced a bla SHV5-type ESBL, first reported from this serotype. Sporadic occurrence of ESBL producer strains among human isolates does not necessarily suggest medical intervention. However, as most of the examined strains of food and animal origin were susceptible and none of them was found to produce ESBL, these facts led us to assume that the observed multidrug resistance and ESBL production of this particular human isolate might rather be related to the human medical treatment than to food consumption, conveying an ESBL-producing strain of Salmonella Bovismorbificans.
Literature data about the different pathogenic determinants of Salmonellae encoded by either SPIs (Marcus et al., 2000) or virulence plasmids (Rotger and Casadesús, 1999) mostly deal with the characteristics of the best studied serovars Salmonella Typhimurium and Salmonella Enteritidis. The information available on the similar features of the other serovars like Salmonella Bovismorbificans is limited. To get some information on some virulence characteristics of Salmonella Bovismorbificans, we conducted both genetic (PCR) and in vivo (1-day-old chicken model) studies. Virulotyping conducted by PCR screening for the presence of virulence genes representing nine important virulence loci revealed high homogeneity of the strains. They all possessed avrA, ssaQ, mgtC, spi4, and sopB genes, indicating the presence of SPI-1–5, respectively, and missed the phage-related genes sopE and gipA, but retained the phage-related gene sodC1. These results are similar to those obtained by Huehn et al. (2009) who found that the representative strains of the serovars tested for the presence of different virulence genes in DNA microarray were highly homogenous in respect that almost all had the genes located on the chromosomally located pathogenicity islands. Variation within and between the serovars was found only in the occurrence of the phage- and the plasmid-encoded genes that are located on transmissible elements, and therefore can spread horizontally.
As our strains varied only with respect to the presence of the spvC gene-containing large plasmid, the question was raised about its in vivo significance. For that purpose, two spvC+ and two spvC − strains were randomly selected to compare their colonization and invasion ability to each other and to that of a highly invasive Salmonella Enteritidis strain in 1-day-old chicks. The significantly decreased organ invasion obtained for all Salmonella Bovismorbificans strains and the better or similar colonization ability of two out of them compared with that of the Salmonella Enteritidis control strain may contribute to the persistence of this serovar, at least in chicken. The differences obtained in the colonization and invasive ability among the four Salmonella Bovismorbificans strains, especially the very low organ invasiveness of the strain 715, may be attributable to the lack of other, not examined (or not yet known) virulence factors. Alternatively, this low level of organ invasiveness might have been due to a stronger stimulation of heterophylic granulocytes leading to an increased protection against internal organ invasion as suggested by Van Immerseel et al. (2005).
PFGE is the gold standard method that we complemented with plasmid profiling to analyze genetic relationship among the strains. PFGE data revealed the existence of 26 different patterns with the dominance of two pulsotypes (C2 and C5). With the exception of two small groups of strains mainly of swine origin representing the same PFGE pattern, plasmid profile and antibiotic susceptibility, no further clustering was found among the other strains tested. These results are in concordance with those that were obtained by Liesegang et al. (2002), who have found 28 different PFGE patterns with the dominance of some clonal types among the investigated 162 Salmonella Bovismorbificans strains collected from different sources from European and overseas countries.
This study indicates that this serovar is armored with several virulence genes necessary to colonize and persist at least in poultry, but most likely also in different mammalian species. It seems to continuously persist, with a relatively low frequency in different animal reservoirs in Hungary and therefore represent potential threat for food safety and public health.
Footnotes
Acknowledgments
Excellent technical assistance of Mrs. Margit Király is acknowledged. We thank Dr. Burkhard Malorny (BfR, Berlin, Germany) for the Salmonella Bovismorbificans strains of German origin; Ama Szmolka, Judit Szántó, Erika Sajtós, and Márta Puruczki for their help in animal experiments; and Endre Zukál for statistical analysis. We appreciate the financial support for this work provided by the MedVetNet, EU FP6 Network of Excellence.
Disclosure Statement
No competing financial interests exist.