
Abstract
During 2004 and 2006, multidrug-resistant Salmonella enterica subspecies enterica serovar Infantis (Salmonella Infantis) isolates (n = 120) were recovered from broiler cecal samples collected from a meat-processing plant, and the isolates were examined. The study was conducted to detect and characterize extended-spectrum β-lactamase (ESBL)-producing Salmonella Infantis isolates recovered from broiler chickens and determine the mechanisms of transfer of the resistance traits. Extended-spectrum cephalosporins-resistant Salmonella Infantis isolates producing ESBL TEM-52 were detected. The mutant bla TEM-52 gene and the wild-type bla TEM-1 gene that mediated resistance to ampicillin (an extended-spectrum penicillin) and cephalothin (a narrow-spectrum cephalosporin) were located on ∼50-kb conjugative plasmids among β-lactam-resistant (n = 29) isolates. The bla TEM genes did not cotransfer with aadA1, sul1 (both associated with class 1 integrons), tetA, and dfrA5, signifying a chromosomal location of these non-β-lactam resistance-encoding genes. This is the first report describing TEM-52-producing S. enterica from food-producing animals in Japan. An emergence of TEM-type ESBL is an important concern to public health because this readily transferable resistance mechanism threatens the value of the third-generation cephalosporins and may reduce the clinical utility of this class of antibiotics against pathogenic Gram-negative bacteria.
Introduction
S
The principal mechanism of resistance to the extended-spectrum β-lactam antibiotics involves the production of ESBLs and AmpC β-lactamases in Enterobacteriaceae. Generally, ESBLs hydrolyze oxyimino-cephalosporins and monobactams, but not cephamycins, and are inhibited by clavulanic acid (CVA). Conversely, AmpC β-lactamases hydrolyze cephamycins and cephalosporins, and CVA does not inhibit these β-lactamases (David et al., 2006). In Japan, one of the most frequent types of ESBLs in human clinical isolates of enterobacteria was the cefotaxime (CTX)-M family of enzymes (Yagi et al., 2000). Recently, CTX-M-type ESBL-producing Escherichia coli isolates were reported from food-producing animals, including broilers (Kojima et al., 2005). Moreover, Weill et al. (2004b) revealed a horizontal transfer of plasmid-mediated bla CTX-M-9 from poultry and human Salmonella Virchow to the E. coli isolates. Thus, because strains of E. coli and Salmonella Infantis frequently occur together as gut flora in broilers, there exists a possibility of bla CTX-M gene transfer from E. coli to the Salmonella Infantis isolates.
In the late 1990s, Salmonella Infantis became a predominant serovar of Salmonella both in broiler flocks and in retail chicken meat in Japan (Kusunoki et al., 2000; Doi et al., 2003; Shahada et al., 2008). Previously, Salmonella Infantis isolates were reported to exhibit resistance against tetracycline and streptomycin (STR) with or without trimethoprim (TRI) (Asai et al., 2007). During 2003, serotype Infantis from broiler chickens exhibiting bla TEM-mediated ampicillin (AMP) resistance was detected for the first time in the southern part of Japan (Shahada et al., 2006). Since 2005, Salmonella Infantis isolates from broilers exhibiting resistance to the extended-spectrum cephalosporins, such as CTX and ceftazidime (CAZ), were reported from the same area (Shahada et al., 2010). Occurrence of extended-spectrum cephalosporin resistance mediated by bla TEM-52 among Salmonella Infantis isolates from poultry and humans has been reported in France and Belgium (Cloeckaert et al., 2007). Therefore, the present study was conducted to detect and characterize ESBL-producing Salmonella Infantis isolates recovered from broiler chickens and determine the mechanisms of transfer of the resistance traits.
Materials and Methods
Bacterial isolates and antimicrobial susceptibility testing
One hundred and twenty Salmonella Infantis isolates recovered from broiler cecal samples collected from a poultry-processing plant in southern Japan between January 2004 and December 2006 were analyzed in this study. The isolation, identification, and serotyping of the bacteria were performed as previously described (Shahada et al., 2008). Antimicrobial susceptibility was assayed by the standard plate dilution method and the disk diffusion test using Mueller–Hinton agar (Oxoid, Basingstoke, United Kingdom), and isolates were tested for sensitivities to AMP, cephalothin (CET), cefoxitin (FOX), CAZ, CTX, and imipenem (10 μg disks). Non-β-lactam STR, sulfamethoxazole (SUL), oxytetracycline (OTC), and TRI were also tested. The minimum inhibitory concentration (MICs) ranged from 0.125 to 512 μg/mL for all antimicrobials tested. Resistance MIC breakpoints for AMP, CET, FOX, CAZ, SUL, and OTC were interpreted according to the Clinical and Laboratory Standards Institute (CLSI, formerly NCCLS) guidelines (NCCLS, 2002). Disk inhibition zones for IMP were interpreted according to the manufacturer's guidelines (Nissui Pharmaceutical, Tokyo, Japan). The breakpoint proposed by Jones et al. (2005) was used for CTX, and it was set as the midpoint between the peaks of the bimodal MIC distributions (Esaki et al., 2004). Resistance MIC breakpoint for STR was adopted as previously recommended (Shahada et al., 2006), and the breakpoint for TRI was chosen according to the European Committee on Antimicrobial Susceptibility Testing guidelines (EUCAST, 2009). Quality control for susceptibility testing was performed using E. coli ATCC 25922 and Staphylococcus aureus ATCC 29213.
Double-disk synergy assay
The double-disk test was performed as a standard Kirby–Bauer disk diffusion assay onto Mueller–Hinton agar according to confirmatory criteria of CLSI (2007) in three representative isolates exhibiting resistance to extended-spectrum cephalosporins. Kirby–Bauer disks (Nissui Pharmaceutical) containing 30 μg of CAZ and CTX were placed 30 mm apart (center to center) around a disk containing amoxicillin (AMPC, 20 μg) plus CVA (10 μg). Enhancement of the inhibition zone, indicating synergy between CVA and test antibiotics, was regarded as presumptive ESBL production. AMP and CET disks were used in the assay as negative controls.
Polymerase chain reaction detection of resistance determinants
Total DNA was extracted using the InstaGene Matrix Kit (Bio-Rad Laboratories, Hercules, CA) after an overnight culture of bacteria onto the brain–heart infusion broth (Eiken Chemicals, Tokyo, Japan). The β-lactamase-encoding genes and other non-β-lactam resistance-mediating genes were examined by polymerase chain reaction (PCR) using primers and conditions depicted in Table 1. The isolates were examined for the presence of the following genes: bla TEM, bla SHV, bla OXA, bla PSE, Toho-1, bla CTX-M825, and bla CTX-M914 encoding β-lactamases; bla FOX, bla MOX, bla CIT, bla DHA, bla ACC, and bla EBC encoding AmpC cephamycinases; aadA1 that mediates STR resistance; tetA that encodes tetracycline efflux proteins; dfrA5 that confers resistance to TRI; sul1 that encodes resistance to SUL; and intI1 that encode class 1 integrons integrase. Amplification of the variable region of class 1 integrons was performed using the 5′-conserved segment (CS) and 3′-CS primers.
T a, annealing temperature; F, forward primer; R, reverse primer; CS, conserved segment.
Sequencing of TEM and class 1 integrons amplicons
PCR cycle sequencing employed the same primers used for the conventional PCR described above. The purified amplified fragments of the two β-lactam-resistance phenotypes representatives (AMPRCETR, n = 3; AMPRCETRCAZRCTXR, n = 3) were sequenced on both strands by the dideoxynucleotide chain termination method with the BigDye Terminator v3.1 Ready Reaction Sequencing Kit and an ABI 3100 automated DNA sequencer (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions. The resulting DNA sequence data were compared with the published DNA sequences using the BLASTN program (
Plasmid extraction and resistance transfer determination
Plasmids carried by all β-lactamase producers were extracted by the alkaline lysis method of Kado and Liu (1981), and purified using a GFX Micro Plasmid Prep Kit (Amersham Biosciences, Buckinghamshire, United Kingdom). The molecular size of plasmids was determined by the standard Salmonella Typhimurium DT104 strain 300-98 (90 kb) and Salmonella Choleraesuis ATCC 7001 (50 kb). To test the plasmid transferability, conjugal mating was performed onto the brain–heart infusion broth with Salmonella Infantis donor isolates (AMPRCETR, n = 3; AMPRCETRCAZRCTXR, n = 3) and the plasmid-free rifampicin-resistant E. coli JM 109 recipient derivatives. Transconjugants were selected onto deoxycholate hydrogen sulfide lactose (DHL) (Eiken Chemicals) agar containing 128 μg/mL rifampicin and 128 μg/mL AMP, followed by antimicrobial susceptibility testing and the detection of transferred resistance determinants as described above.
Southern blot hybridization
Southern blot hybridization using plasmid DNA was conducted in 30 β-lactamase producers isolated from the study area (including one Salmonella Infantis isolate detected for the first time from the same locale in 2003) and selected E. coli transconjugants (n = 6), to confirm the location of the bla TEM gene. Plasmid DNA was subjected to 0.8% agarose gel electrophoresis using 1× Tris–Borate–ethylenediaminetetraacetic acid buffer. Hybridization with a digoxigenin-labeled PCR-generated bla TEM probe was carried out using nylon membranes (Hybond-N+) as recommended by the manufacturer (Amersham Biosciences).
Results
Antimicrobial susceptibility and double-disk synergy testing
Out of 120 Salmonella Infantis isolates, 29 (24%) showed resistance to AMP, 28 (23%) exhibited resistance to CET, and 11 (9%) were resistant against CAZ and CTX (Table 2). Resistance MICs for CAZ and CTX in these isolates ranged from 32 to 128 and 16 to 64 μg/mL, respectively. Besides, both CAZ and CTX presented characteristic bimodal MIC distributions with clearly distinguishable susceptible and resistance peaks. ESBLs were detected in those isolates screened by the double-disk synergy assay, whereby inhibition zone differences of >5 mm between CAZ/CVA and CAZ or CTX/CVA and CTX disks were observed. All isolates resistant to β-lactam antibiotics also exhibited STR/SUL/OTC/TRI resistance phenotype (Table 2). Majority of the β-lactam-resistant isolates were susceptible to FOX except two isolates, which showed reduced susceptibility (MIC = 32 μg/mL). Conversely, all isolates were susceptible to imipenem with the inhibition zone diameters >13–16 mm. The emergence of resistance to CAZ and CTX mediated by ESBL in Salmonella Infantis was observed for the first time in two isolates detected in 2005, and additional nine isolates were recovered in 2006.
These isolates (n = 2) exhibiting MIC = 32 μg/mL were presumed to show reduced susceptibility to FOX partly due to the absence of blaCMY AmpC β-lactamase-encoding genes and a unimodal (susceptible) MIC distribution.
The CTX resistance MIC breakpoint for Enterobacteriaceae was recommended by Jones et al. (2005).
MIC, minimum inhibitory concentration; AMP, ampicillin; CET, cephalothin; FOX, cefoxitin; CTX, cefotaxime; CAZ, ceftazidime; STR, streptomycin; SUL, sulfamethoxazole; OTC, oxytetracycline; TRI, trimethoprim.
Detection and sequence analysis of resistance determinants
The bla TEM-1 gene (GenBank accession no. FJ 527492) was detected in Salmonella Infantis isolates exhibiting resistance against AMP and CET, and the bla TEM-52 gene (GenBank accession no. AF027199) was found in ESBL producers. Sequencing analysis revealed that the 1080-bp bla TEM-52 amplicon differed from bla TEM-1 by three point mutations, Glu163 → Lys, Met241 → Thr, and Gly297 → Ser. Other families of β-lactamase-encoding genes bla SHV, bla OXA, bla PSE, bla CTX-M, and bla CMY tested negative by PCR in all β-lactamase producers. Non-β-lactamase-encoding genes aadA1, sul1, tetA, and dfrA5 were detected in all β-lactam-resistant isolates. PCR amplification of the class 1 integrons yielded a consistent fragment of 1000 bp in all isolates. The sequences of obtained amplicons in three representative isolates showed the presence of the aadA1 gene cassette (GenBank accession no. AB126599).
Conjugation and Southern blot hybridization
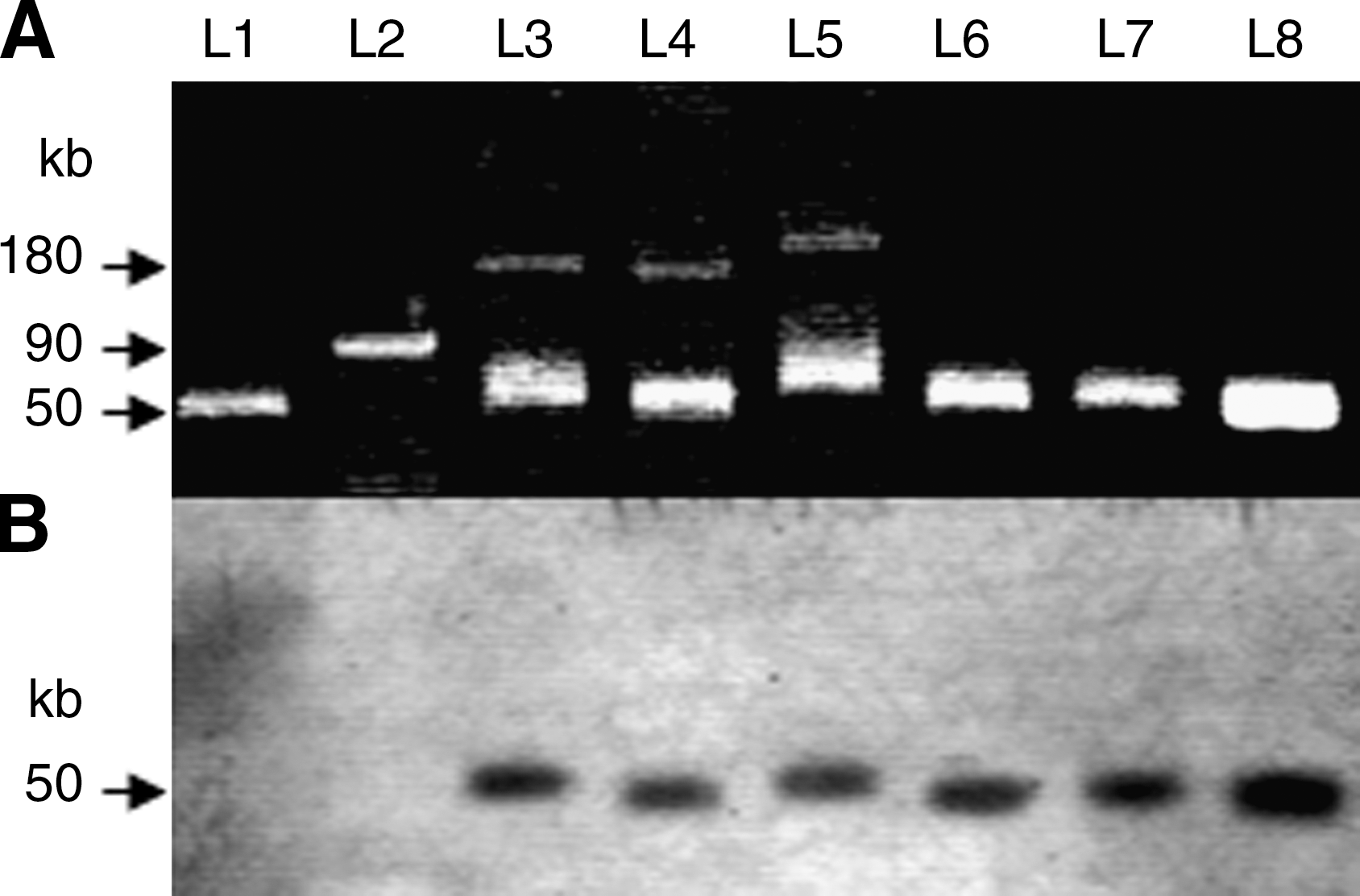
Two large plasmids of ∼50 and 180 kb were detected in the parent isolates; among them, only the plasmids of ∼50 kb were readily transferred by conjugation to the E. coli recipients (Fig. 1A) that involved the transmission of β-lactam resistance phenotypes (Table 3). The 50-kb transferable plasmids produced strong signals after hybridization with the TEM probe both in Salmonella Infantis donor isolates and E. coli transconjugants (Fig. 1B). Sequence analysis of the generated PCR amplicons confirmed the conjugal transfer of both bla TEM-1 and bla TEM-52 genes (Table 3). Conversely, all transconjugants were susceptible to FOX, STR, SUL, OTC, and TRI, and they tested negative when screened for the class 1 integrons intI1, aadA1, sul1, tetA, and dfrA5 genes (Table 3).
(
Reduced susceptibility against FOX.
Discussion
In the present study, we detected for the first time Salmonella isolates from broilers exhibiting resistance to the extended-spectrum cephalosporins mediated by TEM-52 ESBL. The enzyme TEM-52 was first reported in a Klebsiella pneumoniae strain isolated from hospitalized children in 1996 in France (Poyart et al., 1998). The first TEM-52-producing Salmonella was isolated in 1998 from hospitalized Yugoslavian infant (Vahaboglu et al., 2001). Salmonella serotype Saintpaul-, Stanley-, Agona-, and Enteritidis-producing TEM-52 were reported from hospitalized patients in Korea in 2003 (Lee et al., 2003). The occurrence of TEM-52 ESBL in Salmonella isolates of animal origin was reported in France and Belgium between 2001 and 2005 (Weill et al., 2004a; Cloeckaert et al., 2007). The emergence of ESBL-producing Salmonella isolates in Japan is a new phenomenon raising public health concerns, as resistant isolates may be transmitted to humans through the food chain.
It is noteworthy that one of the TEM-1 β-lactamase-producers characterized in the present study was isolated for the first time from the same study area in 2003 (Shahada et al., 2006). This isolate, together with other TEM-1 producers, presented resistance to AMP and CET, but they were susceptible to the extended-spectrum cephalosporins. TEM-1, a narrow-spectrum β-lactamase conferring resistance to the penicillins and the first-generation cephalosporins, differs from TEM-52 by three point mutations, Glu → Lys, Met → Thr, and Gly → Ser (Weill et al., 2004a). We also observed similar three point mutations when the nucleotide sequences of the 1080-bp bla TEM-52 amplification products were compared with the nucleotide sequence of bla TEM-1 gene.
On the other hand, nucleotide sequencing of class 1 integrons revealed the presence of a single aadA1 gene cassette located in the variable region and the sul1 gene located at the 3′-CS. Similar findings were previously reported on Salmonella Infantis isolated from Japan (GenBank accession no. AB126599).
Conjugation experiments revealed a readily transfer of the conjugative plasmids to the E. coli recipients. The location of the bla TEM-52 gene was confirmed by blot hybridization to be ∼50-kb transferable plasmids. Similar size plasmids were carried by other isolates exhibiting resistance to the narrow- and extended-spectrum cephalosporins. Of major concern was the observation that the genetic determinants responsible for the β-lactam resistance were harbored by this type of readily transferable plasmids. Indeed, there exists a possibility of horizontal dissemination of these transferable plasmids to other intestinal flora of the animal hosts that may lead to a further spread of the resistance traits. Cloeckaert et al. (2007) reported the dissemination of the bla TEM-52 gene among various Salmonella serotypes and E. coli strains, suggesting a horizontal interspecies transmission of this resistance gene. Moreover, genes encoding CTX-M- and TEM-type ESBLs have been found coexisting in similar transferable plasmids observed in multidrug-resistant Salmonella serotypes (Weill et al., 2004b; Chouchani et al., 2006).
Given a continued emergence of cephem-resistant Enterobacteriaceae, including ESBL-producing E. coli and Salmonella serotypes, the appropriateness of current CLSI susceptibility breakpoints has been the focus of recent attention. Besides, reports describing preclinical, pharmacokinetic–pharmacodynamic target attainment, and clinical data have emerged supporting a reduction in the CTX MIC breakpoint (Jones et al., 2005; Bhavnani and Tillotson, 2008); in Europe, cephalosporin breakpoints have already been reduced 2- to 16-fold (EUCAST). By comparing shapes of the CTX bimodal MIC distributions in this study, a trend may be developing that support the proposed reduction of CTX MIC breakpoint. However, more data are needed to accumulate evidence and justify the proposed MIC breakpoint reduction.
In conclusion, this is the first report describing the emergence of TEM-52 ESBL-producing Salmonella from food-producing animals in Japan. Although there are no reports describing the occurrence of TEM-52 ESBL-producing Salmonella in humans, this observation may raise public health concerns because resistance can disseminate to humans via the consumption of reported contaminated domestic raw chicken. The findings warrant a systematic, detailed study to detect ESBL-producing Gram-negative bacteria in farms and along the food chain to determine the actual incidence and, hence, establish the magnitude of the problem.
Footnotes
Disclosure Statement
No competing financial interests exist.